2種野海棠屬植物的光合特性
2011-02-01 01:21:28錢仁卷劉洪見許利國張旭樂
浙江農業科學 2011年3期
錢仁卷,劉洪見,黃 建,許利國,張旭樂,林 霞
(浙江省亞熱帶作物研究所,浙江 溫州 325005)
2種野海棠屬植物的光合特性
錢仁卷,劉洪見,黃 建,許利國,張旭樂,林 霞
(浙江省亞熱帶作物研究所,浙江 溫州 325005)
采用LI-6400便攜式光合儀分析浙江2種野海棠屬植物的光合作用特性。結果表明,中華野海棠的光飽和點和補償點分別約為 769 μmol·m-2·s-1和 11.5 μmol·m-2·s-1,最大凈光合速率為 4.88 μmol·m-2·s-1; 秀麗野海棠的光飽和點和補償點分別約為2 270 μmol·m-2·s-1和 2.04 μmol·m-2·s-1,最大凈光合速率為4.18 μmol·m-2·s-1,晴天的凈光合速率日變化為雙峰曲線,次峰明顯低于首峰;中華野海棠凈光合速率與氣孔導度、光合有效輻射呈顯著正相關,與蒸騰速率、溫度呈極顯著正相關,與胞間 CO2濃度、相對濕度呈極顯著負相關 ;秀麗野海棠凈光合速率與氣孔導度呈極顯著正相關,與胞間 CO2濃度呈極顯著負相關,但與蒸騰速率、光合有效輻射、溫度、相對濕度的相關性不明顯。
中華野海棠;秀麗野海棠;光合速率;飽和點;補償點;相關性
野海棠屬是野牡丹科 (Melastomataceae)中的一個屬。該屬植物葉形美觀,亮麗,花色鮮艷奪目,具有較高的觀賞價值。該屬在浙江分布的有3種,秀麗野海棠 (Bredia amoenaDiels)、方枝野海棠 (Bredia quadrangularisCogn)、中華野海棠(Bredia sinensis),其中在溫州地區分布的主要為中華野海棠和秀麗野海棠,都為灌木,植株高度50~80 cm,在瘠薄向陽的山頂和潮濕陰暗的林下、水溝邊都有分布,既耐瘠薄又耐陰耐濕,適應性很強。非常適合用于園林應用,但目前尚未開發,也未發現有相關研究報道。本研究主要測定其光合作用及相關生理生態因子,旨在了解它們的光合生理生態特性,為野海棠屬植物的開發應用提供理論依據。
1 材料與方法
供試材料為野生條件下生長健壯的中華野海棠和秀麗野海棠成年植株,平均株高分別為75 cm和55 cm,生長于文成縣百丈祭海拔約700 m的林下。
用LI-6400便攜式光合儀系統測定植株功能葉片 (主枝上部4~6片葉,每種5株,每株1葉)的光合速率,同時記錄氣孔導度、胞間CO2濃度、蒸騰速率、葉溫、氣溫、光合有效輻射等生理生態因子。于2010年5月上旬晴天測定光合作用日變化,生理生態因子由儀器記錄,測定時段為7:00-18:00,每隔1 h測定1次;于2010年5月9日9:00-11:30測定光合-光強響應,晴天,24~28℃,相對濕度40% ~60%,采用人工光源,光合有效輻射分別設定為 0、20、50、80、100、200、300、600、1 000、1 500、2 000 μmol· m-2·s-1。光響應曲線采用光響應曲線新模型[1]:

式中:Pn是凈光合速率,I是光合有效輻射,α,β,γ是3個系數,Rd是暗呼吸速率。
數據分析和圖表采用EXCEL和DPS完成。
2 結果與分析
2.1 光合作用日變化
從圖1可以看出,2種野海棠在5月份的Pn日變化趨勢均表現為典型的雙峰曲線,秀麗野海棠的首峰出現較中華野海棠早,出現在8:00左右,峰值約為 3.60 μmol·m-2·s-1,中華野海棠要晚約1 h,首峰出現在9:00左右,峰值約為4.82 μmol·m-2·s-1,隨后均呈下降趨勢,于 12:00左右二者均達到最低值,分別為 2.73 μmol·m-2·s-1和 1.57 μmol·m-2·s-1,中華野海棠最低值要高于秀麗野海棠,隨后二者同步平、緩上升,次峰則大致同步,均出現在14:00左右,峰值分別為 4.02 μmol·m-2·s-1和 2.34 μmol·m-2·s-1,之后呈逐漸下降,但中華野海棠下降較快,18:00 左右下降到約 0.29 μmol·m-2·s-1,秀麗野海棠下降緩慢,18:00左右約為1.24 μmol·m-2·s-1,2種野海棠的次峰均明顯低于首峰,兩峰之間有一低谷,說明二者均有明顯的光合“午休”現象,這可能與中午溫度高、空氣濕度低、CO2虧缺、氣孔部分關閉及RuBP羧化酶活性降低等生態、生理和生化因子有關[2-4]。
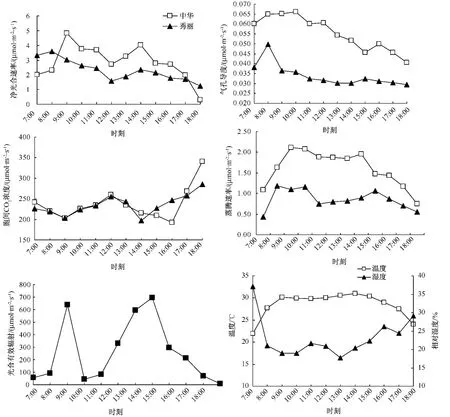
圖1 中華野海棠和秀麗野海棠光合特性的日變化
2.2 光合日變化和主要生理生態因子的關系
中華野海棠的Pn與氣孔導度 (Gs)變化趨勢在7:00-18:00基本保持一致 (圖1),與蒸騰速率 (Tr)在7:00-9:00,10:00-12:00保持一致,9:00-10:00和12:00-15:00變化趨勢相反,15:00后又保持一致;胞間CO2濃度日變化為逆向雙峰曲線,首谷和首峰出現時間與Pn變化較一致,但次谷延續到16:00才出現,之后呈快速增加趨勢。秀麗野海棠的Gs和Tr在7:00-8:00逐漸上升,在8:00達到峰值,和Pn一致,Gs在8:00-9:00迅速下降,之后緩慢平穩下降,Tr則呈現雙峰曲線趨勢,但低谷出現要比Pn約早1 h,而次峰則要晚1 h,Gs變化也是逆向雙峰曲線,首谷出現時間比Pn變化要晚約1 h,其余時間段變化趨勢較為一致。I變化成雙峰趨勢,7:00-9:00迅速上升至首峰,隨后10:00左右迅速下降至最低值,隨后緩慢上升于14:00左右達次峰,隨后迅速下降。溫度變化為類似鐘形曲線,7:00-9:00逐漸上升,之后在30℃左右保持平穩,14:00達最大值31℃,隨后緩慢下降,而相對濕度在7:00-8:00迅速下降,之后在20%上下波動,15:00后開始上升。
經相關分析,中華野海棠的Pn與Gs、Tr、I、溫 度 的 相 關 系 數 分 別 為 0.597 7*、0.908 9**、0.652 9*、0.720 1**,均達到顯著正相關,其中與Tr、溫度為極顯著正相關,與胞間CO2濃度、相對濕度的相關系數分別為 -0.744 1**和 -0.639 9*,為極顯著和顯著負相關;秀麗野海棠的Pn與Gs相關系數為0.846 3**,為極顯著正相關,與胞間CO2濃度相關系數為 -0.754 2**,為極顯著負相關,與I、Tr、溫度、相對濕度的相關性均不明顯。
2.3 光合-光強響應
在短期的 0 ~ 2 000 μmol·m-2·s-1光強條件下,中華野海棠出現光抑制現象,光強超過1 000 μmol·m-2·s-1時光合速率明顯下降,而秀麗野海棠光抑制現象不明顯。經新光合模型擬合得到中華野海棠的光補償點約為 11.5 μmol·m-2·s-1,光飽和點約為 769 μmol·m-2·s-1,最大凈光合速率約為 4.88 μmol·m-2·s-1(R2=0.991 5),相比之下,秀麗野海棠光補償點更低,約為2.04 μmol·m-2·s-1,而光飽和點遠高于中華野海棠,約為2 270 μmol·m-2·s-1,最大凈光合速率約為4.18 μmol·m-2·s-1(R2=0.996 0)。由此可見,與白鶴芋[5]等觀葉植物相比,中華野海棠和秀麗野海棠的光補償點更低,說明二者均具有較強的耐蔭性;中華野海棠的光飽和點也驗證了其光抑制現象,表明中華野海棠不適宜強光下生長,而秀麗野海棠的光飽和點相比美麗異木棉[6]、琯溪蜜柚[7]等喜光植物均要高,表明其具有陽生性,適應能力更強,適合在更大范圍內生長。但2種野海棠的最大凈光合速率均較低,這可能是二者生長緩慢的重要原因之一。中華野海棠的Tr、胞間 CO2濃度以及Gs隨光強增加的變化趨勢和光合速率變化均較為一致;秀麗野海棠的胞間CO2濃度與Pn變化較一致,而Gs和Tr隨I增加開始迅速上升,但隨后變化較為波動,與Pn變化不一致 (圖2)。

圖2 2種野海棠凈光合速率、氣孔導度、胞間CO2濃度和蒸騰速率對光合有效輻射的響應
3 小結與討論
中華野海棠光補償點均較低,光飽和點較秀麗野海棠低,因此在今后的引種栽培過程中應采取適當遮蔭的措施,以利于其生長。
中華野海棠和秀麗野海棠的光合“午休”現象出現的主要原因是與高溫、低濕有關,特別是氣溫或葉溫過高引起了氣孔的部分關閉,蒸騰減弱,因此,在今后的引種栽培實際中,特別是在夏季高光強下出現的干熱天氣,要注意降溫、保濕,如采用生草法、人工覆草、葉面噴霧等降低周圍小環境的溫度,減弱光合抑制的程度。
植物的光合作用受生理生態因子的綜合影響,簡單的相關性分析方法顯得有些片面,并且因子間的互作 (如溫度與胞間CO2濃度)對光合作用的影響不是很清楚,因此生理生態因子與光合作用的關系還有待今后進一步探求。
[1] 葉子飄.一個新的光響應曲線模型在超級雜交稻組合:П優明86中的應用 [J].生態學雜志,2007,26(增刊):1323-1326.
[2] 葉慶生,潘瑞熾,丘才新.墨蘭葉片結構及光合作用的研究 [J].植物學報,1992,34(10):771-776.
[3] 許大全.光合作用效率 [M].上海:上海科學技術出版社,2002:39-52.
[4] 蘇培璽,杜明武,張立新,等.日光溫室草莓光合特性及對CO2濃度升高的響應 [J].園藝學報,2002,29(5):423-426.
[5] 江如藍,劉偉.觀葉植物白鶴芋的光合特性研究 [J].廣東農業科學,2001(5):25-27.
[6] 吳吉林,李永華,葉慶生.美麗異木棉光合特性的研究[J].園藝學報,2005,32(6):1061-1064.
[7] 易干軍,姜小文,霍合強,等.琯溪蜜柚光合特性的研究[J].園藝學報,2003,30(5):519-524.
S 865.99
A
0528-9017(2011)03-0529-03
文獻著錄格式:錢仁卷,劉洪見,黃建,等.2種野海棠屬植物的光合特 [J].浙江農業科學,2011(3):529-531,535.
2011-03-04
基于生態園林城市建設的花境植物引選及其應用技術研究 (2006C22076)
錢仁卷 (1981-),男,浙江平陽人,碩士,主要從事花卉生理與生物技術研究工作。
(責任編輯:張才德)
