豬源性乙型腦炎病毒分離株全基因序列測定和分析*
2012-01-24 07:44:44趙靈燕謝榮輝朱函坪楊章女姚蘋蘋周小龍朱智勇
中國人獸共患病學報 2012年1期
關鍵詞:分析
趙靈燕,謝榮輝,朱函坪,徐 芳,楊章女,姚蘋蘋,周小龍,朱智勇
豬源性乙型腦炎病毒分離株全基因序列測定和分析*
趙靈燕1,謝榮輝2,朱函坪2,徐 芳2,楊章女2,姚蘋蘋2,周小龍2,朱智勇2
目的了解豬源性乙型腦炎病毒(乙腦病毒)的基因特征。方法對豬源性乙腦病毒株(JX61)進行全基因組序列測定和基因進化特性的分析。結果豬源性乙腦病毒JX61株全基因組全長均為10 964個核苷酸,含有一個開放閱讀框架,編碼3 432個氨基酸。與GenBank中選擇的41株乙腦病毒全基因序列比較發現,其核苷酸同源為83.6%~99.3%,氨基酸總體同源性為94.9%~99.6%。通過全基因序列進行系統進化分析均顯示該毒株屬于基因I型乙腦病毒。結論豬源性乙腦病毒JX61株屬于基因I型,與國內分離株XJP613關系最為接近。提示豬體內的乙腦病毒基因型別有了新的變化,這也是浙江省在豬體第一次分離基因I型乙腦病毒。
乙腦病毒;序列分析;種系發生
乙型腦炎病毒又稱日本腦炎病毒(JEV),簡稱乙腦病毒,是引起流行性乙型腦炎的病原體,具有較強的傳染性和致病性,其傳播地域廣,病死率高,嚴重威脅著人類的健康[1]。乙腦病毒通常在蚊一豬—蚊等動物間循環,其中豬被認為是乙腦最重要的傳染源和乙腦病毒傳播的主要宿主,在乙腦的流行環節上起重要的作用。因此加強豬體中乙腦病毒的流行狀況及變異研究對預防和控制乙腦流行具有重要的意義。目前國內從豬體內分離到乙腦病毒除了HN0701株以外均為基因III型乙腦病毒,較少見到基因I型乙腦病毒,但本實驗室2008年從浙江省嘉興地區分離到的3株乙腦病毒均為基因I型乙腦病毒,提示豬體內乙腦病毒基因型別可能發生了新的變化。為了更好的了解乙腦病毒在豬體內的變異狀況,本研究測定了豬源性乙腦病毒分離株(JX61株)全基因組序列并對其分子生物學特征進行了分析,現報道如下。
1 材料與方法
1.1 病毒株 2008年采自浙江嘉興某豬場,經BHK-21分離獲得并經熒光定量RT-PCR鑒定為乙腦病毒,編號為JX61,-80℃保存備用。
1.2 主要試劑和菌種 RNA提取試劑盒式購Qiagen公司,M-MLV逆轉錄酶購自Promega公司;Taq DNA聚合酶、DNA純化回收試劑盒、質粒提取試劑盒購自Takara公司。大腸桿菌DH5α感受態細胞購自Takara公司,PGEM-T克隆載體購自Promega公司。
1.3 引物設計和合成 根據GenBank中乙腦病毒中乙腦病毒的全基因序列設計了9對相互重疊片段的引物,引物由上海生工生物工程有限公司合成(表1)。

表1 實驗中所用引物Table 1 Primer used in the study
1.4 病毒RNA的提取 將病毒接種到長滿單層的BHK-21細胞瓶中,接種3~4d,觀察病變。當75%以上的細胞發生病變時,收獲病毒,用Qiagen公司的RNA提取試劑盒進行抽提。
1.5 RT-PCR和克隆 使用Promega公司的 MMLV,按照說明書操作,用引物合成cDNA,然后用Taq酶分段進行PCR,PCR產物進行瓊脂糖凝膠電泳后,切下目的條帶,用膠回收試劑盒回收目的片段。并將目的片段克隆于T載體,轉化DH5α感受態細胞,在含有Amp、X-gal和IPTG的LB平板培養基上培養18h。選取菌落進行培養,堿裂解法提取質粒,進行陽性克隆株的PCR鑒定。
1.6 序列測定和分析 篩選的陽性克隆經PCR鑒定,選2株送上海生工生物工程有限公司測序,用DNAStar和Clustal X生物軟件對其進行序列分析。
1.7 中和試驗 將SA14-14-2免疫人體后得到的陽性血清以1∶10、1∶20、1∶40和1∶80稀釋后分別與等體積的100TCID50病毒稀釋液(JX61株病毒)混合,37℃水浴作為1h后,將其接種于96孔細胞板于37℃細胞培養箱培養1h后,棄去液體后補加100μL放置于37℃細胞培養箱繼續培養并觀察病變,每個血清稀釋度平行做4孔。同時設病毒對照組和乙腦病毒陰性血清對照組。
2 結 果
2.1 乙腦病毒浙江分離株JX61的基因組結構JX61由10 964個核苷酸組成(登錄號GU556217),5非編碼區由96個核苷酸組成,3非編碼區含596個核苷酸組成。97至10 392位編碼區,共10 296個核苷酸,編碼3 432個氨基酸。JX61株與基因Ⅲ、Ⅱ、Ⅳ型和基因I型的KV1899和K94P05乙腦病毒序列比較分析表明,JX61在5′NTR的91位存在1個核苷酸的插入(位點以P3株為準,以下皆同),且JX61和其他基因I型乙腦病毒一樣在10 395~10 407均存在著13個核苷酸的缺失,可作為基因I型乙腦毒株的分子標記之一。本文中引用的國內基因I型和豬源性乙腦病毒分離株病毒名稱、分離時間、分離地點及標本來源背景資料見表2。
2.2 與疫苗株SA14-14-2 ORF編碼區的差異分析JX61株與疫苗株SA14-14-2 ORF碼區存在1 201個核苷酸差異,核苷酸同源性88.7%,但核苷酸大部分突變屬于同義突變,僅引起89個氨基酸核苷酸變異,氨基酸同源性為97.4%。其中在結構蛋白中共有26個氨基酸差異,7個在衣殼蛋白(C)區段,4個在膜蛋白(M)區段,15個在囊膜蛋白(E)區段,其中以衣殼蛋白的差異率為最高,達到5.5%。在非結構蛋白共有63個氨基酸差異,NS1蛋白區段有12個氨基酸差異,NS2蛋白區段有14個氨基酸差異,NS3蛋白區段有11個氨基酸差異。NS4蛋白區段有5個氨基酸差異,NS5基因區段有21個氨基酸差異。
2.3 新分離病毒JX61株與其他乙腦病毒毒株全序列比較和基因組進化樹的構建 JX61與41株乙腦毒株的全基因核苷酸及氨基酸序列進行同源性比較分析,JX61株與其他毒株的核苷酸總體同源性為83.6%~99.3%,JX61株與其他毒株氨基酸同源性均為94.9%~99.6%。JX61株與SH17M-07和XJP613株核苷酸同源性最高,均為99.3%,與JKT6468株核苷酸同源性最低,為83.6%。JX61與SH17M-07和Mie40株的氨基酸同源性最高,均為99.6%,與JKT6468株的氨基酸同源性最低,為94.9%。JX61株與減毒活疫苗株SA41-14-2及P3株的核苷酸同源性只有88.6%和89.1%,但氨基酸同源性分別為97.4%和97.8%。

表2 本研究中使用乙腦病毒毒株信息Table 2 Details of JEV strains from China used for analysis in this study
2.4 新分離株JX61 E基因比較分析 采用減毒活疫苗株SA14-14-2為標準,對新分離的乙腦病毒JX61的E區段核苷酸及氨基酸進行分析。與疫苗株之間核苷酸同源性為87.7%,氨基酸同源性分別為97.0%。新分離株與減毒活疫苗SA14-14-2株存在15個氨基酸位點差異,分別位于Domain I區的E-138,E-176,E-177位,Domain II區的 E-107、E-129、E-222、E-244、E-264、E-279 位,Domain III區的 E-315、E-327、E-366,還有3個位于這3個活性區外的 E-433,E-439和 E-447,而影響病毒抗原抗體結合的E304和E335二個位點的半胱氨酸則沒有發生變異。新分離株與國內其他基因I型病毒的核苷酸同源性在97.5%~99.3%。氨基酸同源性為98.4%~99.8% (結果末列)。通過對幾十株基因I型乙腦病毒比較結果表明:基因I型乙腦病毒E222位都為S,而基因III乙腦病毒則為A。對GenBank上中國豬源性乙腦病毒分離株進行E蛋白序列比較發現,大多數分離株屬于基因III型乙腦病毒,而本實驗室分離的JX61、JX66、JX67和河南分離株HN0701則屬于基因I型乙腦病毒,表明近年來我國在豬體內的基因型別已經有了新的變化。JX61株與HN0701核苷酸同源性為98.4%,與本實驗室分離的豬源性乙腦病毒分離株JX66及JX67的核苷酸同源性則為100%。而與國內其他豬源性分離株的核苷酸同源性均在90%以下,而氨基酸同源性則均在97%以上,表明豬源性乙腦病毒核苷酸雖然差異較大,但大多數屬于沉默突變,氨基酸差異較小。分析表明近年來國內分離的豬源性乙腦病毒可分為基因I型和基因III型乙腦病毒,國內豬源性基因III型乙腦病毒E基因序列比較顯示各毒株之間核苷酸同源性最低為99.2%、最高為100%。國內豬源性基因I型乙腦病毒E基因序列比較顯示各毒株之間的核苷酸同源性最低為98.4%,最高為100%。而豬源性基因I型與基因III型乙腦病毒E基因核苷酸同源性較低,均在90%以下,但氨基酸同源性較高,均在97%以上。
2.5 JX61株進化分析 對JX61株與41株乙腦病毒全基因組序列進行系統進化分析,結果顯示明顯的4個基因型別,其中基因I型11株,基因II與IV型各1株,基因III型29株。新分離乙腦病毒JX61株屬于基因I型乙腦病毒,與2007年分離株XJP613和SH17M-07最接近。
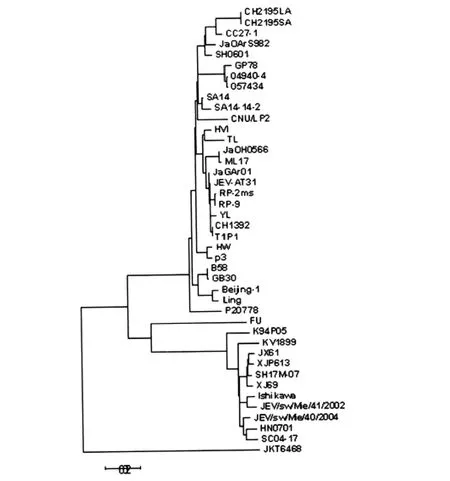
圖1 分離自不同國家的42株乙腦病毒全基因序列系統進化分析Fig.1 Phylogenetic trees based on full-length genome of genetic relationships among 42 virus strains
2.6 現行減毒活疫苗免疫機體后產生抗體與JX61株的中和狀況 結果顯示當陽性血清1∶10和1∶20稀釋后與等體積的100TCID50病毒量(JX61株)混合作用后,接種于BHK-21細胞4d后4孔均末發生病變,而當陽性血清1∶40與等體積的100TCID50病毒量(JX61株)混合作用后,接種于BHK-21細胞4d后有3孔則發生了病變。病毒對照組和陰性血清對照組則均發生了病變。表明疫苗株SA14-14-2株免疫人體后產生的抗體能中和JX61株,提示現行疫苗對JX61株具有保護作用。
3 討 論
乙腦病毒通常在蚊-豬-蚊等動物間循環,乙腦病毒在豬群的長時間貯存與循環可促進病毒的變異,增加環境中該病毒的濃度,而使人感染乙腦病毒的概率提高。因此加強對豬機體內乙腦病毒的研究有助于該類疾病的預防和控制。目前國內外學者已成功地從豬體內分離到多株乙腦病毒并對其進行研究,研究結果顯示在日本、韓國等國家豬源性乙腦病毒既有基因I型乙腦病毒又有基因III乙腦病毒,但在中國豬源性乙腦病毒通常為基因III型乙腦病毒[2-5]。而本研究2008年從浙江嘉興某豬場采集的豬血清中分離到的JX61則屬于基因I型乙腦病毒,提示基因I型乙腦病毒在我國部分地區的豬體內存在。以往研究表明近年來國內分離到的豬源性乙腦病毒大多數為基因III型乙腦病毒,且同一地域、豬源性乙腦病毒毒株之間核苷酸和氨基酸均具有較高的同源性,但JX61株則與國內豬源性乙腦病毒的同源性較低,而與蚊蟲中分離的基因I型乙腦病毒具有較高的同源性,說明基因I型乙腦病毒在自然界中蚊-豬-蚊循環途徑中的各種媒介宿主中存在并進行傳播,但基因I型乙腦病毒何時在豬體中流行有待于進一步研究。
系統進化分析顯示疫苗株SA14-14-2和分離株JX61株屬于不同基因型乙腦病毒,但中和試驗表明SA14-14-2減毒活疫苗免疫機體后產生的抗體能中和JX61株,提示現行疫苗對目前流行的基因I型乙腦病毒同樣具有保護作用。與SA14-14-2株ORF區核苷酸和氨基酸的比較分析顯示存在1 201個核苷酸差異,核苷酸同源性為88.7%,但核苷酸大部分突變屬于同義突變,僅引起89個氨基酸變異,氨基酸同源性為97.4%。此外SA14-14-2株和JX61株在主要保護性抗原E蛋白氨基酸對比結果顯示:(1)JX61株與SA14-14-2在 E337-345,E377-E382、E397-403三個中和位點的氨基酸均相同[3];(2)JX61株和SA14-14-2株在E304和E335均為半胱氨酸,而這二個半胱氨酸形成的二硫鍵是抗原抗體的必要結構[6];(3)二者的氨基酸同源性高達97.0%。以上這些結果從分子水平上表明現行疫苗對我國分離的不同基因型乙腦病毒都會有很好的免疫保護作用。
目前常采用Pr M/C,E基因片段對乙腦病毒進行基因分型,不能夠完全反映乙腦病毒基因組的變異程度,存在一定的不足之處[7-8]。但乙腦病毒株的全基因組序列進化分析最能夠準確地描述乙腦病毒株之間的進化關系。JX61與其他毒株相比較,核苷酸 總 體 差 異 率 為0.7% (XJP613、SH17M-07)-16.4% (JKT6468),氨 基 酸 差 異 率 為 0.4%(SH17M-07、Mie40)-5.1%(JKT6468)。從全基因序列進化分析得各JX61株與浙江蚊子分離株XJP613最接近,同屬于基因I型乙腦病毒。本實驗室自2007年以來從浙江省蚊子和豬體內相繼分離到多株基因I型乙腦病毒,提示在浙江省自然界中流行基因I型乙腦病毒,而基因I型乙腦病毒流行的增多很可能對當地乙腦的發病、流行特征產生新的影響,因而需要密切地監測乙腦病毒在自然界中的進化,為更好預防和控制乙腦提供理論依據。
[1]Tsai TF.New initiatives for the control of Japanese encephalitis by vaccination:minutes of a WHO/CV1 meeting,Bangkok,Thailand,13-15 october 1998[J].Vaccine,2000,26:1-25.
[2]Tang WF,Ogawa M,Eshita Y,et al.Molecular evolution of Japanese encephalitis virus isolates from swine in Oita,Japan during 1980-2009[J].Infect Genet Evol,2010,10(2):329-336.
[3]Yang DK,Kim BH,Kwon JH,et al.Molecular characterization of full-length genome of Japanese encephalitis virus(KV1899)isolated from pigs in Korea[J].J Vet Sci,2004,5(3):197-205.
[4]Nidaira M,,Maira K,Okano S,et al.Survey of Japanese encephalitis virus in pigs on Miyako,Ishigaki,Kume,and Yonaguni islands in Okinawa,Japan[J].Jpn J infect Dis,2009,62(3):220-224.
[5]王環宇,梁國棟.中國基因3型乙型腦炎病毒E基因分子特征[J].中華微生物學和免疫學雜志,2010,30(5):387-391.
[6]Wu SC,Lin CW.Neutralizing peptide ligands selected from phage displayed libraries mimic the conformational epitope on domain III of the Japanese encephalitis virus envelope protein[J].Virus Res,2001,76:59-69.
[7]Wang HY,Takasaki T,Fu SH,et al.Molecular epidemiological analysis of Japanese encephalitis virus in China[J].J Gen Virol,2007,88(3):885-894.
[8]Nga PT,Parquet M del Carmen,Cuong VD,et al.Shift in Japanese encephalitis virus(JEV)genotype circulating in northen Vietnam:implications for frequent introductions of JEV from Southeast Asia to East Asia[J].J Gen Virol,2004,85(6):1625-1631.
Complete sequence analysis of Japanese encephalitis virus JX61 strain from swine
ZHAO Ling-yan,XIE Rong-hui,ZHU Hai-ping,XU Fang,YANG Zhang-nu,YAO Ping-ping,ZHOU Xiao-long,ZHU Zhi-yong
(Centre of Animal Disease Control of Zhejiang Province,Hangzhou 310020,China)
In order to understand the genetic characteristics of Japanese encephalitis virus from swine,an JEV was isolated from swine,and its complete genome was sequenced and analyzed comparatively.The results of sequencing of the nucleotides demonstrated that the genome of strain JX61 was 10964 nucleotides in length with a single open reading frame encoding 3432 amino acids.Comparison of the complete genome sequences of different JEV isolates showed JX61 was 83.6%-99.3%nucleotide sequence homology among them,which resulted in 94.9%-99.6%amino acid sequence homology.Phylogenetic analysis through full-length genome showed that the JX61 strain belonged to genotypeⅠ.The phylogenetic analysis based on the entire genome demonstrated that JX61 strain has the nearest genetic relationship with XJP613 and the JX61 might belong to the genotype I.It was the first isolation of Japanese encephalitis virus genotype I from swine in Zhejiang province during recent years.
Japanese encephalitis virus;sequence analysis;phylogeny
R373.3
A
1002-2694(2012)01-0037-04
*浙江省醫藥衛生科學研究基金項目(2008B 39)
謝榮輝,Email:ghost_rhxie@hotmail.com
1.浙江省動物疫病預防控制中心,杭州 310020;2.浙江省疾病預防控制中心,杭州 310051
2011-04-06;
2011-06-21
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
