扣帶回腦區與癲癇大鼠學習記憶損害相關性分析
2012-06-01 06:18:16宋石磊王莉趙勇張萬琴
中國實用醫藥 2012年19期
宋石磊 王莉 趙勇 張萬琴
癲癇是常見的中樞神經系統疾病之一。目前對于癲癇的研究主要還是采用動物模型,其中海仁酸(KA)所致的癲癇模型已廣泛用于癲癇發作的研究,被認為是經典的癲癇動物模型[1]。長期以來對于癲癇所致學習記憶障礙的研究主要集中在海馬腦區,并且肯定了海馬腦區與癲癇學習記憶障礙的相關性,但是對于扣帶會在這方面的研究甚少。本研究通過對難治性癲癇大鼠腦內扣帶回腦區C-Jun免疫反應性變化的觀察,初步探討扣帶回與難治性癲癇空間學習記憶障礙的相關性。
1 材料與方法
1.1 制備及分組 KA誘發癲癇急性發作及癲癇持續狀態取20只大鼠,一次頸部皮下注射驚厥劑量(10 mg/kg,2 ml/kg)的KA誘發SD大鼠出現癲癇急性發作及癲癇持續狀態。癲癇的嚴重程度按Racine[2]描述的標準進行1~5級分級,在本實驗中有12只大鼠有4級以上發作并呈持續狀態,其中有5只死亡,7只存活大鼠被采用為癲癇模型即模型組。剩余未達到四級以上發作的10只大鼠有4只死亡,6只存活大鼠作為模型對照組,另取同批SD大鼠(n=6)頸部皮下注射等容積(2.0 ml/kg)的生理鹽水即為空白對照組。
1.2 Morris水迷宮行為學測定 SD大鼠在分組完成后30 d采用Morris水迷宮通過隱藏平臺獲得實驗及空間搜索實驗對大鼠的學習記憶能力進行測定。
1.3 灌流固定
1.3.1 4%水合氯醛400 mg/kg腹腔注射麻醉大鼠,開胸暴露心臟。
1.3.2 16 g注射針頭刺入左室尖部,止血鉗固定,剪開左心耳,先后以0.9%氯化鈉注射液200 ml和4%多聚甲醛400 ml先快速后緩慢進行灌注,至肝臟、四肢及頭尾變硬。
1.3.3 斷頭取腦,于4℃4%多聚甲醛中后固定。
1.4 腦片制備及免疫組織化學實驗 將固定好的鼠腦放入含30%蔗糖的磷酸緩沖液中4℃過夜,振動切片機切取50 μm厚的冠狀腦片,PBS漂洗10 min×2,與1%BSA孵育30 min;加入第一抗體(C-jun-Ab,1∶500)4℃孵育過夜,PBS漂洗10 min×2;加入生物素化羊抗兔(1∶400)室溫振蕩孵育1 h,PBS漂洗10 min 2;與卵白素生物素復合物A:B:PBS(1∶1∶400)室溫振蕩孵育2 h,二氨基聯苯胺(DAB)法顯色;適時用0.01 m PBS終止反應;明膠裱片,風干;梯度酒精脫水:70%、80%、90%、95%、100%、100%酒精各脫水 5 min;二甲苯透明,中性樹膠封片、晾干。
1.5 圖像分析 采用HPIAS系列彩色病理圖文分析系統對C-Jun免疫陽性細胞進行定量計數,采用Olympus-BX51型顯微攝像系統對目標腦區進行攝像。分析方法:選取扣帶回為目標腦區,分別在扣帶回5個不同層面分別選取4個寬度 和高度分別為213、289 μm的計數區,在顯微鏡下進行觀察,計數C-Jun陽性細胞數目,取其平均值。并分別攝片。
1.6 統計學方法 實驗采用SPSS 11.5軟件進行統計處理。免疫組化數據采用均數±標準差,各組腦片之間的免疫活性差異采用單因素方差分析;不同處理組間的兩兩比較采用單因素方差分析S-N-K法;檢驗標準為P<0.05。
2 實驗結果
2.1 Morris水迷宮行為學測定結果
2.1.1 隱匿平臺獲得實驗 模型組大鼠的尋找潛伏期在各個時間段都明顯長于其它2組大鼠。并且其在學習測試的4 d中每天的尋找潛伏期沒有明顯差異(P>0.05)。模型對照組與空白對照組的尋找潛伏期在各個時間段差異無統計學意義(P>0.05)。
2.1.2 空間探索試驗 模型組大鼠穿越平臺的次數平均為0.71,而其它2組分別為2.33次、3.29次,差異有統計學意義(P<0.05),模型對照組與空白對照組之間的差異無統計學意義(P>0.05)。
2.2 免疫組織化學結果 在C-jun的免疫組化結果中,模型組大鼠扣帶回腦區C-Jun免疫陽性細胞數目較空白對照組組和模型對照組大鼠明顯降低,有顯著差異(P<0.05)。而模型對照組扣帶回腦區C-Jun免疫陽性細胞數目與空白對照組相比則無明顯差異(P>0.05)。(圖1、表1)。
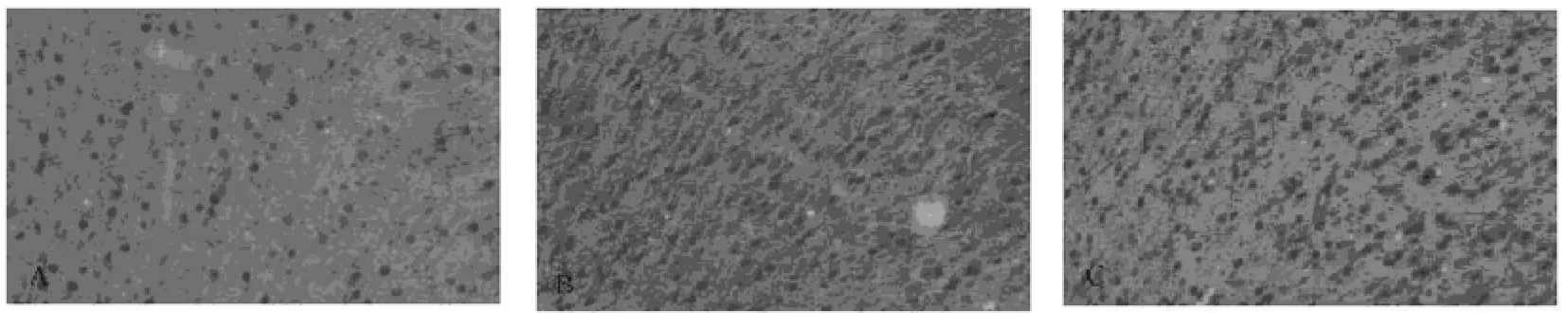
圖1 A模型組B模型對照組C空白對照組(200)

表1 各組大鼠扣帶回腦區C-Jun陽性細胞數目
3 討論
C-Jun是一種即刻早期基因(IEGs),它的表達受控于突觸的活動,有研究指出其在突觸地可塑性機制(LTP)中發揮重要的作用,而LTP是目前公認的記憶形成基礎[3]。在經過記憶訓練的大鼠腦內可以觀察到C-Jun蛋白表達的升高[4],這些研究表明了C-Jun參與了學習記憶的形成過程。通過抑制C-Jun氨基末端激酶(JNKs)造成C-Jun mRNA和蛋白質合成的損傷,可以改善短時程記憶而阻斷長時程記憶[5],也說明了C-Jun與學習記憶的相關性。
現在共識的記憶形成的分子機制假說是:刺激發動長時程記憶的相關基因的表達進而啟動分子級聯反應,級聯反應又可以調控IEGs的表達,最終達到記憶相關區域持久的結構和功能的改變[6]。在這個過程中隨著各種類型的細胞興奮,C-Jun會被誘導并表達,C-Jun的免疫反應活性可作為神經元興奮的標志[7]。在大鼠學習過程中,學習記憶相關腦區如果C-Jun表達降低,就很有可能是該腦區的學習記憶機制受到了損害。
本實驗中通過免疫組化實驗對各組大鼠的扣帶回腦區進行的觀察發現實驗組大鼠扣帶回腦區內C-Jun有著相對較低的表達,與此相應的是在行為學實驗中,模型組的學習記憶能力與空白組和實驗對照組相比明顯降低,而模型對照組與空白組相比學習記憶能力無明顯差異。結合以上對C-Jun在學習記憶形成過程中作用的探討,有理由認為實驗組大鼠扣帶回腦區內的學習記憶機制發生了損害。所以癲癇大鼠學習記憶能力的改變很可能與扣帶回腦區學習記憶機制損害相關。
[1]Sperk G.Kainic acid seizures in therat.Prog Neurobiol,1994,42:1.
[2]Racine RJ.Modification of seizure activity by electrical stimulation.Ⅱ.Motor seizure.Electroencephalogr Clin Neurophysiol,1972,32:281.
[3]Herdegen T,Skene P,Bahr M.The c-Jun transcription factorbipotentialmediator of neuronal death,survival and regeneration.Trends Neurosci,1997,20:227-231.
[4]Lisa A,Teather,Mark G,Packard,et al.Differential induction of c-Jun and Fos-like proteins in rat hippocampus and dorsal striatum after training in two water maze tasks.Neurobiology of Learning and Memory,2005,84(2):75-84.
[5]Bevilaqua LR,Kerr DS,Medina JH,et al.Inhibition of hippocampal Jun N-terminal kinase enhances short-term memory but blocks long-term memory formation and retrieval of an inhibitory avoidance task.Eur J Neurosci,2003,17,897-902.
[6]P Goelet VF,Castelucci S,Schaecher,et al.The long and the short of long-term memory-a molecular framework.Nature,1986,322:419-422.
[7]J I Morgan,T Curran.Stimulus-transcription coupling in the nervous system:Involvement of the inducible proto-oncogenes fos and jun.Annual Review of Neuroscience,1991,14:42-451.
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
中國民間療法(2021年5期)2021-06-09 09:21:04
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
作文周刊·小學一年級版(2016年27期)2017-06-03 23:21:17
飲食科學(2017年5期)2017-05-20 17:11:53
新湘評論·下半月(2016年4期)2016-05-05 22:12:41
新湘評論·下半月(2016年4期)2016-05-05 22:12:41
海外文摘(2016年4期)2016-04-15 22:28:55
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
