20個青菜品種的遺傳多樣性分析
2012-06-29 02:20:12馬曉靜李廣平鄭曉靜林冬枝董彥君
長江蔬菜 2012年22期
關鍵詞:分析
馬曉靜 ,李廣平 ,鄭曉靜 ,林冬枝 ,2,董彥君 ,2
(1.上海師范大學生命與環境科學學院,200234;2.上海師范大學遺傳研究所)
青菜(Brassica chinensisL.)屬十字花科蕓薹屬青菜種,別稱小白菜、青梗白菜等,是我國常見的綠葉蔬菜之一[1],在我國長江三角洲地區占有著重要地位,市場需求量大[2,3]。目前對青菜種質資源的鑒別主要依據形態學,而在DNA分子水平對其遺傳多樣性研究不多。李紅斌等[4]利用AFLP分子標記技術對浙江省主栽的10個青菜品種進行了遺傳多樣性及聚類分析。近年來,更多DNA分子標記的發展及應用[5~11],為種質資源遺傳多樣性的研究提供了更多手段,其中SSR分子標記技術廣泛應用于種質資源遺傳多樣性方面的研究[12~15],但未見利用SSR分子標記技術對青菜進行遺傳多樣性分析的報道。本研究利用10對SSR分子標記技術對來自中國和日本的20個青菜品種進行遺傳多樣性分析,探索其親緣關系,以期為青菜種質改良和創新提供分子水平上的理論依據。
1 材料與方法
1.1 試驗材料
供試材料為來自我國長江三角洲地區主栽的8個青菜品種以及從日本引進的12個青菜品種,具體見表1。2012年4月1日將其播種于上海師范大學奉賢校區生物基地內,30 d后取其植株的新鮮幼嫩葉片,-70℃保存備用。

表1 供試青菜品種
1.2 DNA模板的提取
用改良的CTAB法[16]提取青菜葉片基因組DNA,-20℃保存。
1.3 引物篩選
本研究選用88對隨機引物對樣品進行PCR擴增,篩選出譜帶清晰、位點數量及產物大小適中且適用于20個青菜品種遺傳多樣性分析的引物,對樣品進行正式擴增。
1.4 PCR擴增及聚丙烯酰胺凝膠電泳
PCR反應在eppendorf擴增儀上進行,反應體系為 10 μL, 其中 1 μL 10×Buffer with Mg2+,0.2 μL 2.5 mmlo/L dNTP,0.25 μL 1 μmol/L 引 物 ,0.25 μL 5 U/μL Taq 酶,5 μL 模板 DNA,最后用無菌水補足至 10 μL。 反應條件如下:94℃預變性 4 min,94℃變性 30 s,50℃退火 30 s,72℃延伸 60 s,35 個循環;最后72℃延伸10 min,產物4℃保存備用。擴增產物在8%的聚丙烯酰胺非變性凝膠上電泳分離3.5 h(恒定功率30 W),以銀染法檢測電泳結果。
1.5 數據統計與分析
PCR擴增產物在銀染過的聚丙烯酰胺凝膠上都表現為譜帶的“有”和“無”,分別將其賦值,有帶的記為1,無帶的記為0,建立所用材料分子標記的0、1矩陣。根據表征矩陣,統計擴增產物的條帶總數和多態性條帶數量。
利用NTSYS 2.10e軟件分析材料間的Nei's遺傳相似系數。根據遺傳相似系數矩陣,用非加權類平均法(UPGMA法)進行聚類分析,建立聚類圖。
2 結果與分析
2.1 引物篩選
從88對隨機引物中篩選出譜帶清晰、位點數量及產物大小適中且適用于遺傳多樣性分析的10對引物(表2)。其擴增產物的條帶總數和多態性條帶結果見表3。由表3可知,其擴增的條帶總數為26條,其中多態性條帶有20條,占擴增條帶總數的76.9%,說明供試青菜品種間具一定的遺傳差異。
2.2 聚類分析
由圖1可知,以遺傳相似系數0.708為界,可將20個青菜品種劃分為3類,其中第Ⅰ類包括CR夏專科、夏八仙、長江;第Ⅱ類包括大葉黑大頭、嘉興矮腳青、上海五月慢、特矮蘇州青、上海矮抗青;第Ⅲ類包括長陽、上海四月慢、夏三味、敦煌、夏賞味2號、NiHAO 夏、陽夏、一江、華王、陽太、抗熱 605、紫冠一號(表 4)。

表2 引物篩選結果
3 結論與討論
本研究所篩選出的10對SSR分子標記中絕大多數的多態性比率較高(表3),20個青菜品種的遺傳相似系數在0.461 5~0.923 1,變異范圍較大,說明其遺傳多樣性較高,同時也說明,SSR分子標記可作為青菜分子育種中有力的輔助工具。
另外,利用10對SSR分子標記的聚類分析結果表明,供試的20個青菜品種可分為三大類 (表4),其中第Ⅰ類中CR夏專科、夏八仙、長江均來自日本;第Ⅱ類中大葉黑大頭、嘉興矮腳青、上海五月慢、特矮蘇州青、上海矮抗青則均來自我國;而第Ⅲ類中除上海四月慢、抗熱605、紫冠一號以外,其余均為日本品種。本試驗中將來自中國的青菜品種五月慢和抗熱605歸屬為不同類群,與李紅斌等[4]的結果一致。總之,本試驗結果顯示絕大部分供試青菜品種,來自同一國家的聚在一起,說明其親緣關系較近,而來自不同國家的青菜品種之間的親緣關系則較遠,這可能是因為長久缺乏種質交流而使絕大部分供試青菜品種呈現明顯的地域性。由此可見,地域限制可能是促成青菜種質資源遺傳分化的主要原因之一。因此,我們可以引入外來種質,將其與國內青菜品種進行雜交,實現性狀的改良,以滿足青菜品種的市場需求。

表3 10對引物的擴增結果
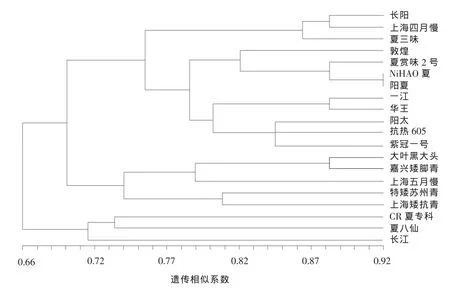
圖1 20個青菜品種的UPGMA聚類分析

表4 供試材料聚類結果
[1]郝春燕.上海地區青菜品種應用現狀及潛力品種推薦[J].長江蔬菜,2009(7):9-11.
[2]駱義成,戚立剛,沈玉興,等.青菜新品種夏帝引種示范試驗項目總結[J].上海蔬菜,2010(6):10.
[3]龔靜,朱玉英,吳曉光,等.青菜新品種“新矮青”的選育[J].上海農業學報,2002,18(1):39-42.
[4]李紅斌,王世恒,王宏.青菜品種親緣關系AFLP分析 [J].北方園藝,2008(8):39-40.
[5]Thomson M J,Septiningsih E M,Suwardjo F,et al.Genetic diversity analysis of traditional and improved Indonesian rice(Oryza sativaL.)germplasm using microsatellite markers[J].Theoretical and Applied Genetics,2007,114(3):559-568.
[6]Ma C Y,Ma H Y,Xu Z,et al.Isolation and characteristics of polymorphic microsatellite markers inPennahia argentatus[J].Biochemical Systematics and Ecology,2012,43:159-162.
[7]王娟,郭大龍,侯小改,等.不同花型牡丹品種親緣關系的SRAP 分析[J].中國農學通報,2011,27(28):167-171.
[8]魏玉杰,張金文,何慶祥,等.不同生態區罌粟種質的遺傳多樣性 ISSR 分析[J].植物遺傳資源學報,2012,13(2):239-243.
[9]王愛華,季祥彪,文曉鵬.16份野生半夏的遺傳多樣性分析[J].貴州農業科學,2012,40(1):15-19.
[10]劉麗,楊新龍,劉楚吾,等.南海海域常見龍蝦的遺傳多樣性分析[J].水產科學,2012,31(3):160-164.
[11]孫立夫,裴克全,張艷華,等.中國與歐洲高盧蜜環菌的遺傳多樣性[J].生物多樣性,2012,20(2):224-230.
[12]徐建欣,王云月,姚春,等.利用SSR分子標記分析云南陸稻品種遺傳多樣性 [J].中國水稻科學,2012,26(2):155-164.
[13]倪勝利,李興茂,楊德龍,等.國內外冬小麥品種(系)的遺傳多樣性分析[J].西北農業學報,2012,21(2):20-25.
[14]劉龍洲,翟文強,陳亞麗,等.設施用厚皮甜瓜品種SSR標記遺傳多樣性分析 [J].植物遺傳資源學報,2012,13(3):381-385.
[15]曾維英,梁江,陳淵,等.廣西野生大豆SSR標記的遺傳多樣性研究[J].江蘇農業科學,2012,40(3):22-25.
[16]Murray M G,Thompson W F.Rapid isolation of high molecular weight plant DNA[J].Nucleic Acid Res,1980,8:4 321-4 326.
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
