鉛鋅尾礦區優勢草本植物體內鉛及氮、磷、鉀含量變化特征
2012-08-20 04:06:10張樹金李廷軒鄒同靜熊劍
草業學報 2012年1期
張樹金,李廷軒,鄒同靜,熊劍
(四川農業大學資源與環境學院,四川 成都611130)
隨著工業的發展和農業生產的現代化,環境問題越來越受到人們關注,重金屬引起的土壤污染也日益成為環境、土壤科學工作者研究的熱點問題。鉛(Pb)已成為環境中最普遍的重金屬污染物之一,其主要來源于人類活動如采礦、冶煉、焚燒、含鉛汽油、顏料及化肥和殺蟲劑的施用等[1]。目前,國內外用來降解或消除土壤重金屬的基本方法分為物理法、化學法和生物修復法三大類,但前2類方法往往投資昂貴,需用復雜的設備,大多數只能暫時緩解重金屬危害,而且有可能造成二次污染[2,3]。近年來,包括微生物、植物和動物3種方法在內的生物修復技術成為土壤污染修復的熱點研究領域[4,5]。1983年,美國科學家Chaney首次提出了利用某些能夠富集重金屬的植物來清除土壤重金屬污染的設想——植物修復技術的設想,與傳統方法相比,這項技術以其高效、經濟和生態協調性等優勢顯示出巨大的生命力,很快成為國內外研究的熱點[6-8]。
國際上報道的超富集植物已有400多種,但其富集重金屬的種類還比較有限,其中超富集鎳的植物約占73%[9]。但有關植物對鉛超富集或富集以及其生物學過程則報道較少。通過對我國植物進行廣泛的篩選和調查,首次發現我國一種原生鉛富集植物——東南景天(Sedum alfredii),其地上部最高鉛濃度可達1 182 mg/kg[10]。但現已發現的超富集植物通常生物量低,生長緩慢,修復效率低,難以形成較完善的植物修復技術以直接大規模應用于修復實踐[11]。因此,各國學者就提高超富集植物的生物產量和積累重金屬的能力作了大量研究,主要通過采取改良土壤環境、增加土壤肥力等農藝措施提高超富集植物的生長速度、生物產量以及地上部的重金屬積累量[12,13]。
氮、磷、鉀是植物必需的大量營養元素,在物質組成和代謝過程中有重要的作用,是植物體內蛋白質、核酸等重要物質的合成和代謝過程中不可或缺的組分[14]。植物缺乏大量營養元素的一種或幾種均會導致物質組成和代謝的紊亂,進而引起植物品質、生物量和產量的下降。已有研究表明,氮、磷、鉀均能顯著影響超富集植物對鉛的吸收能力[15,16]。但對重金屬與大量營養元素分布特點及其交互關系的研究不多,而從植物整個生長時期,研究植物重金屬與大量營養元素的相關性甚少[17]。因此,在2007年對三合鉛鋅尾礦區的調查基礎上,從22種草本植物中篩選出9種優勢植物,并于2008年對其氮、磷、鉀和鉛含量進行分析研究。旨在探討不同生長時期植物鉛及氮、磷、鉀元素分布情況及其相關性,弄清大量營養元素與鉛的交互關系,以期為尾礦區重金屬污染的治理和修復提供理論依據。
1 材料與方法
1.1 研究區域概況
研究區位于四川省雅安市滎經縣三合鄉境內(102°31′E,29°47′N),屬中緯度亞熱帶濕潤氣候區,年均氣溫15.3℃,雨量充沛。滎經縣礦產資源豐富,各類礦產規模有64處,其中鉛鋅礦14處,探明貯量共50萬t。三合鉛鋅礦為該地區內具有開采價值的一處,平均含鉛7.62%。采樣區域為2003年停用的尾礦區,面積約1hm2,海拔1 358~1 445m。區域內土壤類型為黃壤。尾礦區土壤表面被礦渣覆蓋,植物以草本植物為主。
1.2 采樣方法
2008年在生長前期(春季,5月)、生長旺盛期(夏季,8月)和生長后期(秋季,10月)3個時期進行采樣。9種優勢植物分別屬于7科(表1),每種植物至少采集3個重復,每個重復取6株以上混合樣。
1.3 樣品分析
采集的植物樣品分為地下部和地上部,分別用自來水充分沖洗,去除粘附于植物樣品上的泥土和污物,再用去離子水沖洗,在105℃下殺青30min,然后在70℃下于烘箱中烘至恒重,稱重。植物樣品粉碎后備用。植物樣品采用HNO3+HClO4(5∶1,V∶V)在160℃用密閉的聚四氟乙烯罐消煮至澄清。消煮后的溶液洗至100mL容量瓶中,用高純水定容。樣品待測液分別用原子吸收分光光度計測定鉛含量[18]。植物樣品通過H2SO4+H2O2消煮法消煮,采用蒸餾法測定全氮,鉬銻抗比色法測定全磷,火焰光度計法測定全鉀[18]。
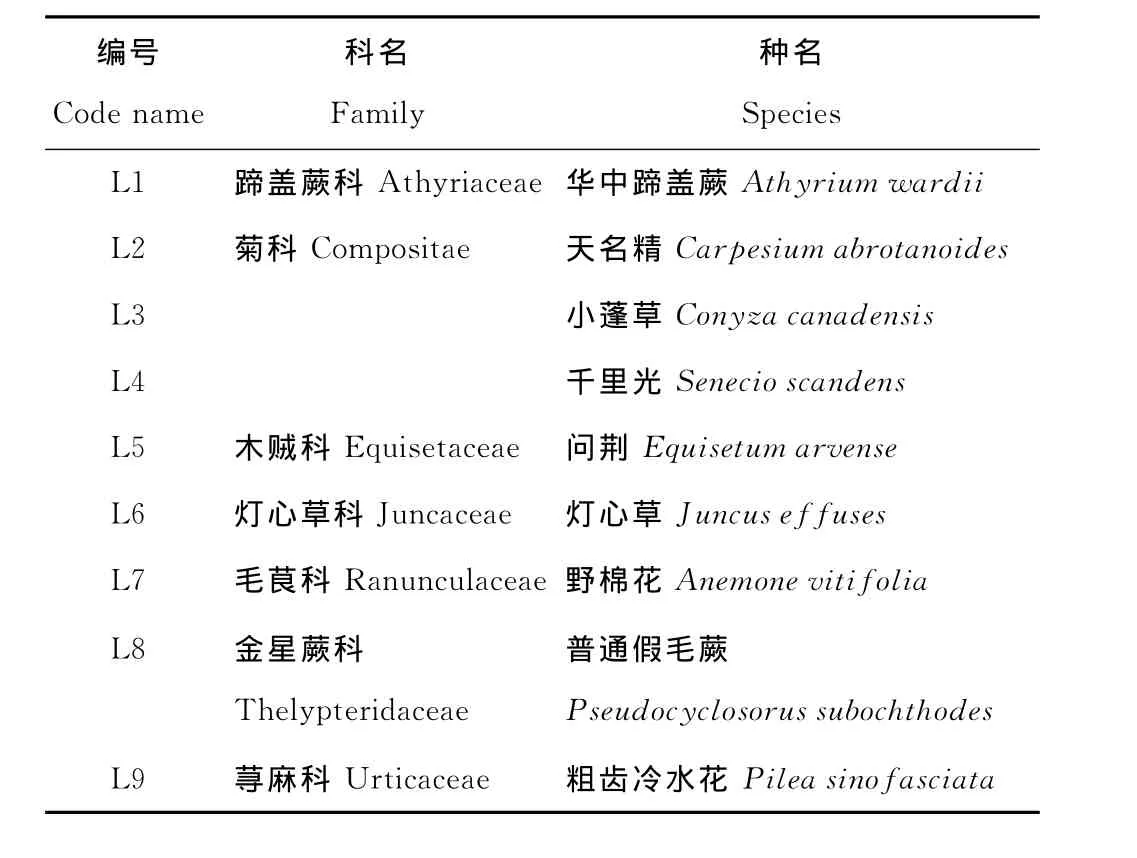
表1 鉛鋅尾礦區優勢草本植物種類Table 1 Family and species composition of the predominant herbaceous plants investigated in the lead-zinc mining tailing
1.4 數據分析
數據用Excel 2003和SPSS 13.0軟件進行統計分析。
2 結果與分析
2.1 植物鉛含量變化特征
植物吸收鉛的能力隨著生長環境的變化而變化,影響植物吸收鉛的因素是多方面的。在不同生長時期,不同植物鉛含量差異很大。植物地下部表現出鉛含量高,變異程度大的特點,且明顯高于地上部(表2)。在3個生長時期,植株地上部鉛含量均小于1 000mg/kg,未達超富集植物的標準。除L1和L8外,其余植物均在生長旺盛期地下部和地上部鉛含量同時達到最大值。
在生長前期,L1、L5、L8地下部鉛含量超過500mg/kg,與其他植物達到極顯著差異。其中L1地下部鉛含量為最高,達到15 542.1mg/kg,為其余植物的21.4~251.1倍;地上部仍以L1為最高,達到656.8mg/kg,為其余植物的2.3~16.1倍。在生長旺盛期,多數植物地下部和地上部鉛含量均有所增加。地下部鉛含量以L1為最高,達到10 720.1mg/kg,為其余植物的25.8~68.8倍;地上部鉛含量以L9為最高,達475.4mg/kg。在生長后期,多數植物地下部和地上部鉛含量表現出下降的趨勢,可能是由于隨著生長期的繼續延長,其生物量增大,產生稀釋效應所致。其地下部和地上部鉛含量均以L1為最高,分別達4 132.2和188.0mg/kg,極顯著高于其余植物的鉛含量。
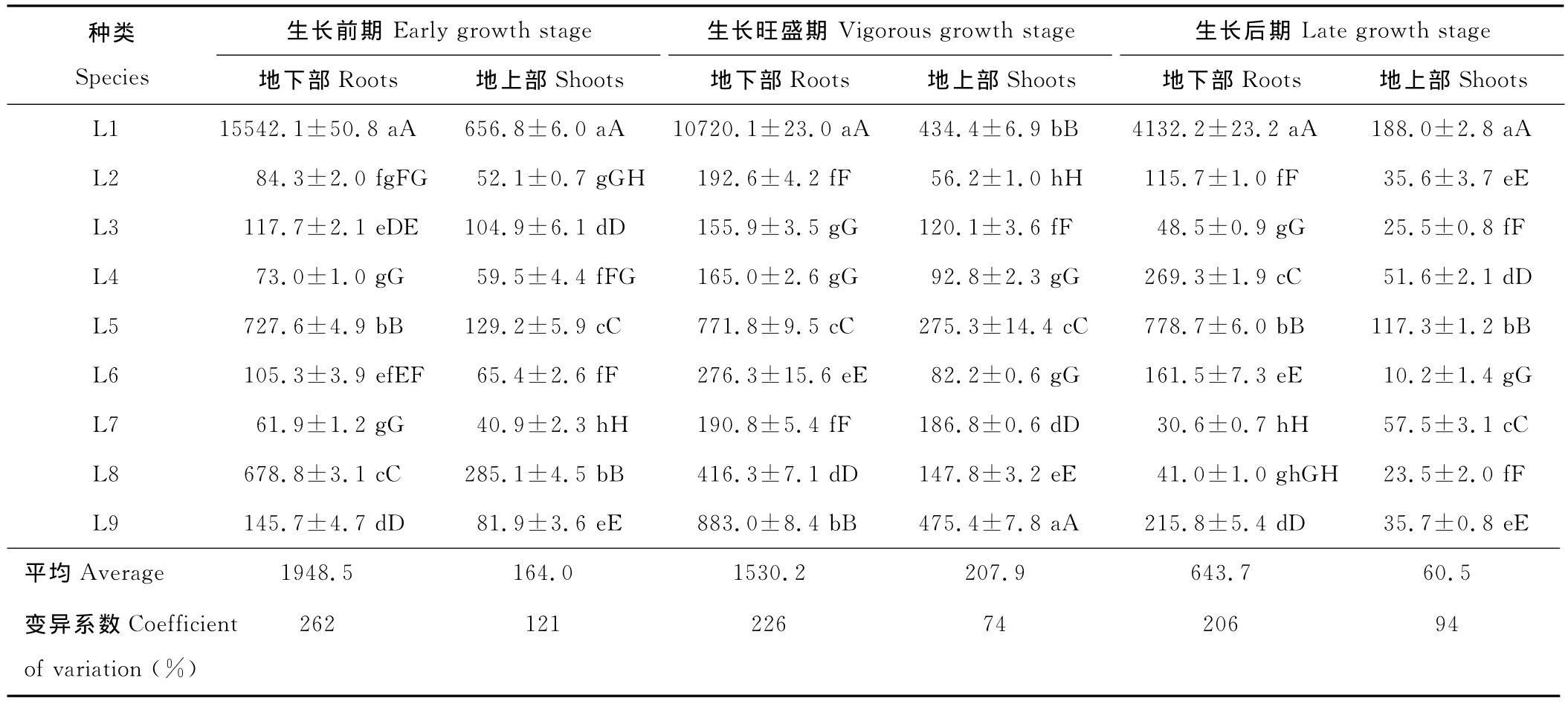
表2 9種優勢草本植物3個生長時期鉛含量差異Table 2 Lead concentration in the roots and shoots of 9predominant herbaceous plant species at three growing stagesmg/kg
2.2 植物氮含量變化特征
氮被稱為生命的元素,它是蛋白質、核酸和磷脂的主要成分,在生命活動中有特殊作用[19,20]。在不同生長時期,不同植物氮含量差異較大(表3)。總體上表現出地上部氮含量高于地下部,在3個生長時期,植物地上部平均氮含量分別為地下部的1.99,2.00和1.74倍。多數植物地下部和地上部氮含量在生長前期達到最大值,隨生長時期的延長,氮含量總體上呈現出下降的趨勢。在生長前期,地下部氮含量為5.52~10.92g/kg,平均為7.93 g/kg。地上部則表現出氮含量相對較高,變異程度大的特點,平均氮含量為15.82g/kg。在生長旺盛期,多數植物氮含量呈現出降低的趨勢,其地下部和地上部平均氮含量分別為5.81和11.62g/kg,與生長前期相比均降低27%。在生長后期,植物氮含量繼續下降,地下部氮含量為2.44~6.76g/kg,平均為4.46g/kg;地上部氮含量為5.64~10.79g/kg,平均為7.77g/kg。與生長前期相比,其地下部和地上部平均氮含量分別下降44%和51%。
2.3 植物磷含量變化特征
磷素有“能量元素”之美譽,是植物正常生長發育不可或缺的營養元素之一,在植物整個生命歷程中起著極其重要的作用[21]。尾礦區9種優勢草本植物磷含量與氮含量的變化趨勢相似(表4),在不同生長時期,不同植物磷含量差異較大。在3個生長時期,總體上表現出地上部磷含量高于地下部,其平均磷含量分別為地下部的1.67,1.85和1.28倍。隨著生長時期的延長,磷含量呈現下降的趨勢。在生長前期,植物地下部磷含量為0.81~1.85 g/kg,平均磷含量為1.28g/kg。地上部則表現出磷含量相對較高,變異程度大的特點,為0.80~3.05g/kg,平均磷含量為2.14g/kg。在生長旺盛期,植物磷含量的變異程度增大,可能是由于在生長旺盛期,不同植物對磷的需求差異較大,導致其選擇吸收和積累的量有差別所致。但其平均磷含量仍略有降低,與生長前期相比,磷含量分別降低16%和7%。在生長后期,植物地下部和地上部平均磷含量分別為1.02和1.31g/kg,與生長前期相比,分別降低20%和39%。
2.4 植物鉀含量變化特征
鉀在植物的生長發育以及產量形成過程中都有十分重要的作用。鉀并不直接參與植物體內組織、器官的形成,而是通過促進植物體內生理、生化活動及代謝反應來調節植物的生長發育及產量形成[22]。在不同生長時期,不同植物鉀含量差異較大,總體上表現出地下部鉀含量低于地上部(表5)。植物地下部和地上部鉀含量均在生長前期或生長旺盛期時達到最大值,隨著生長時期的繼續延長,鉀含量呈現出下降的趨勢,并在生長后期達到最小值。在生長前期,植物地下部鉀含量為2.78~16.04g/kg,平均為10.81g/kg;地上部鉀含量為10.63~32.59 g/kg,平均為19.89g/kg,是地下部平均鉀含量的1.84倍。在生長旺盛期,植物地下部和地上部平均鉀含量分別為10.75和18.95g/kg,后者為前者的1.76倍。在生長后期,植物鉀含量處于較低水平,地下部和地上部平均鉀含量分別為8.42和13.96g/kg,與生長前期相比,分別下降22%和30%。

表3 9種優勢草本植物不同生長時期氮含量差異Table 3 Nitrogen concentration in the roots and shoots of 9predominant herbaceous plant species at three growing stagesg/kg
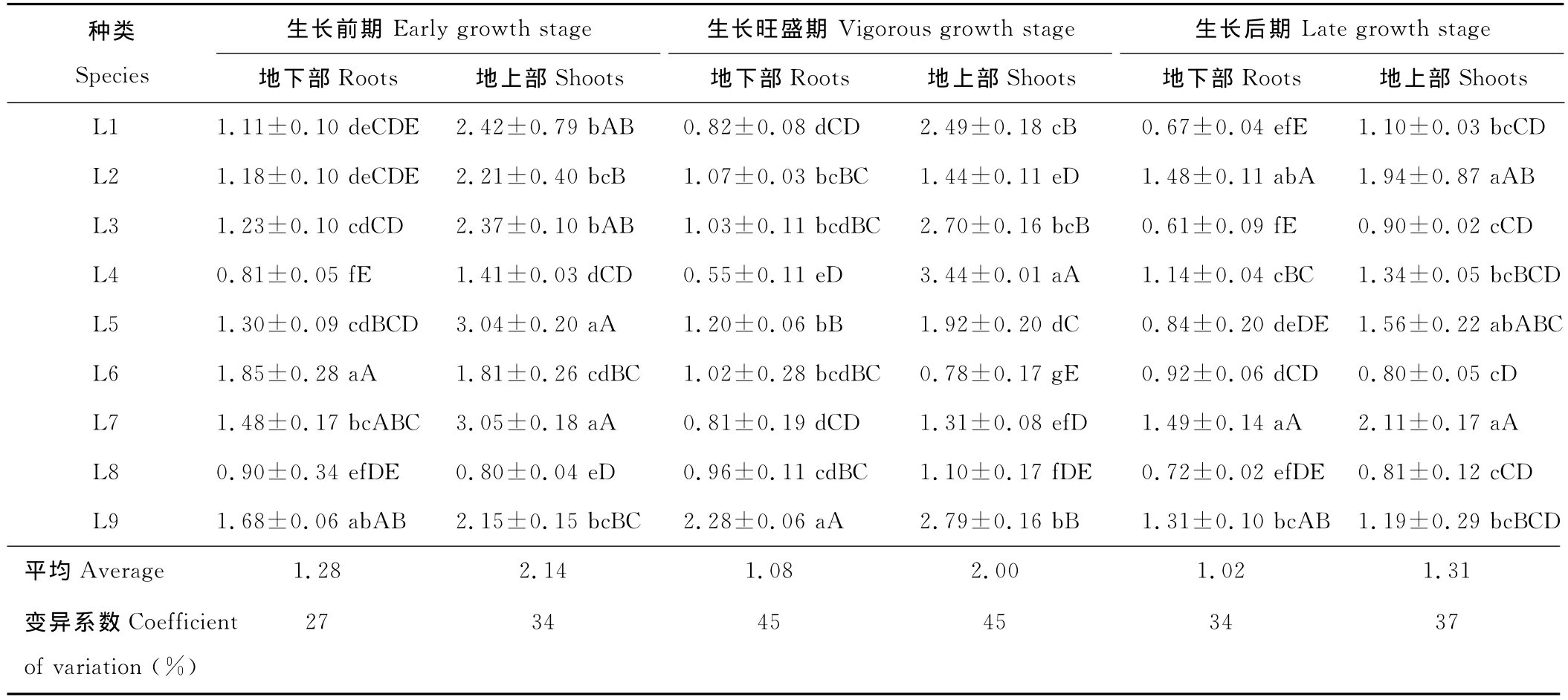
表4 9種優勢草本植物3個生長時期磷含量差異Table 4 Phosphorus concentration in the roots and shoots of 9predominant herbaceous plant species at three growing stagesg/kg

表5 9種優勢草本植物3個生長時期鉀含量差異Table 5 Potassium concentration in the roots and shoots of 9predominant plant herbaceous species at three growing stagesg/kg
3 討論
3.1 植物鉛與氮、磷、鉀含量變化特征
李裕紅等[17]在對泉州灣海岸植物研究指出,鉀為含量最高的大量營養元素,高含量的鉀有助于增強植物的抗逆性。自然環境下生長的泉州灣海岸植物的氮、磷、鉀含量與其銅、鋅、鉛含量之間無顯著相關關系。本研究得出,鉀含量最高,氮次之,磷最低。尾礦區9種優勢草本植物總體上表現出地上部氮、磷、鉀含量高于地下部,并隨著生長時期的延長,呈現出下降的趨勢。這可能是由于隨著生長時期的延長,植物發生木質化,并主要以糖類、纖維素等碳水化合物的形式增加生物量,導致氮、磷、鉀的含量相對降低。植物鉛含量則是地上部小于地下部,可能是由于鉛為植物非必需營養元素,凱氏帶的阻隔將大量的鉛滯留在地下部,這也是至今尚未發現真正意義上的鉛超富集植物的根本原因所在[23]。
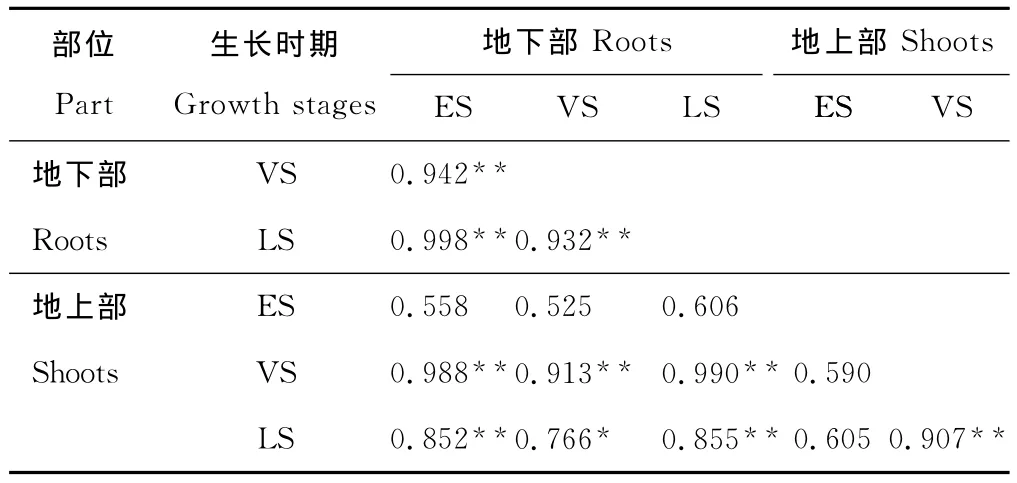
表6 9種優勢草本植物地上部和地下部鉛含量在3個生長時期的相關性分析Table 6 Relationship between lead concentration of root and shoot of 9predominant herbaceous plant species at three growth stages
相關性分析得出,在植物生長前期,其地上部鉛含量與地下部鉛含量以及與生長旺盛期和生長后期的地上部或地下部鉛含量均不存在顯著的相關關系,而在生長旺盛期和生長后期,同一時期地上部和地下部鉛含量之間以及不同時期地上部與地下部鉛含量之間均存在顯著或極顯著的相關關系(表6)。這是因為植物主要通過根系吸收重金屬鉛,并從地下部往地上部運輸,但鉛在植物體內移動性小,不易轉移。由于不同的植物生長速度和轉移能力不同[24],導致在生長前期,9種優勢草本植物地上部吸收和累積鉛的能力存在明顯差異,故無顯著的相關關系。因此,在尾礦區篩選鉛植物修復材料時,應避開在植物生長前期進行采樣。但不同類型的礦區及氣候條件對植物重金屬的吸收累積存在差異,篩選時期可能存在一定的差異。
3.2 大量營養元素在植物修復中的作用
現已發現的超富集植物通常生物量低,生長緩慢,修復效率低,阻礙了植物修復技術以直接大規模應用于修復實踐[25]。因此,前人的研究致力于通過施加外源物質等來 提 高 修 復 效 果[26]。 李 繼 光 等[27,28]研 究 表明,在外界生長環境的氮素含量未達到東南景天的最適臨界點情況下,適當施加氮肥顯著促進東南景天的生長,提高其干物質產量,同時促進了東南景天對鎘的吸收及向地上部分的轉運。孫琴等[12]研究得出,適當增磷顯著促進東南景天的生長,提高其干物質產量,同時促進了東南景天對鋅的吸收及鋅向地上部分的運轉和積累。聶俊華等[16]在對單一營養元素氮、磷、鉀對植物富集鉛的研究中得出,少量的氮和鉀可促進富集植物干質量的增加,促進植物對鉛的吸收,隨著氮和鉀水平的增加,植物對鉛的吸收能力降低,但鉀的抑制作用不如氮的顯著;土壤供磷會降低植物對鉛的吸收,且鉛含量下降達極顯著水平。相關性分析得出,9種優勢草本植物地下部氮、磷、鉀含量與鉛含量均不存在顯著的相關關系(表7),這可能是由于植物吸收氮、磷、鉀的方式與吸收鉛的方式不同及鉛在植物體內的移動性相對較小所致。在生長后期,9種優勢植物地上部氮和鉀含量與鉛含量存在顯著的正相關關系(0.750*和0.773*),表明氮和鉀可能對解重金屬鉛的毒害起到一定的作用。
華中蹄蓋蕨(L1)是一種常見的多年生蕨類植物,這種蕨類植物根系發達、根部鉛吸收能力強、轉運能力弱,是一種新型的鉛穩定修復材料,具有潛在的應用價值[29]。本研究得出,華中蹄蓋蕨能夠將大量鉛囤積在地下部,在3個生長時期,其根部累積量分別為32.64,41.81和21.49mg/株,并維持正常的生長,這可能與其在3個生長時期地上部、地下部氮含量均處于較高水平(表2),大量蛋白質與重金屬結合降低其毒性有關。然而,能否通過增施氮肥,提高華中蹄蓋蕨的穩定修復效率仍需進一步的研究。
4 結論
尾礦區9種優勢草本植物以鉀為含量最高的大量營養元素,氮次之,磷最低,且植物地上部氮、磷、鉀含量均高于地下部,并隨著生長時期的延長呈下降趨勢。植物鉛含量差異很大,在不同生長時期其含量變化也較明顯,表現出地下部明顯高于地上部。9種優勢草本植物均未表現超富集鉛的能力,但華中蹄蓋蕨地下部能夠囤積大量鉛,并保持正常生長,進一步證實其能作為穩定修復的先鋒植物,應進一步研究探討其富集忍耐重金屬的機理。
尾礦區9種優勢草本植物地下部氮、磷、鉀含量在3個生長時期均與鉛含量無顯著的相關關系。在生長后期,地上部氮、鉀含量與鉛含量存在顯著的正相關關系,表明氮和鉀可能對植物解重金屬鉛的毒害起到一定的作用,其機理目前尚不清楚,還有待進一步研究。
在植物生長前期,其地上部鉛含量與地下部鉛含量以及與生長旺盛期和生長后期的地上部或地下部鉛含量均不存在顯著的相關關系,而在生長旺盛期和生長后期,同一時期地上部和地下部鉛含量之間以及不同時期地上部與地下部鉛含量之間均存在顯著或極顯著的相關關系。因此,在尾礦區篩選鉛植物修復材料時,應避開在植物生長前期采樣。但不同類型的礦區及氣候條件對植物重金屬的吸收累積存在差異,篩選時期可能存在一定的差異。
[1] 王云,魏復盛.土壤環境元素化學[M].北京:中國環境科學出版社,1995.
[2] Gworek B.Lead inactivation in soils by zeolites[J].Plant and Soil,1992,143(1):71-74.
[3] Vangronsveld J,Colpaert J V,Van Tichelen K K.Reclamation of a bare industrial area contaminated by non-ferrous metals:Physico-chemical and biological evaluation of the durability of soil treatment and revegetation[J].Environmental Pollution,1996,94(2):131-140.
[4] 龍新憲,楊肖娥,倪吾鐘.重金屬污染土壤修復技術研究的現狀與展望[J].應用生態學報,2002,13(6):757-762.
[5] 丁佳紅,劉登義,儲玲,等.重金屬污染土壤植物修復的研究進展和應用前景[J].生物學雜志,2004,21(4):6-9,20.
[6] Chaney R L,Malik M,Li Y M,et al.Phytoremediation of soil metals[J].Current Opinion in Biotechnology,1997,8(3):279-284.
[7] 李源,李金娟,魏小紅.鎘脅迫下蠶豆幼苗抗氧化能力對外源NO和H2O2的響應[J].草業學報,2009,18(6):186-191.
[8] 李影,王友保.4種蕨類草本植物對Cu的吸收和耐性研究[J].草業學報,2010,19(3):191-197.
[9] González R C,González-Chávez M C A.Metal accumulation in wild plants surrounding mining wastes[J].Environmental Pollution,2006,144(1):84-92.
[10] He B,Yang X E,Ni W Z,et al.Sedum alfredii:A new lead-accumulating ecotype[J].Acta Botanica Sinica,2002,44(11):1365-1370.
[11] Lelie D V D,Schwitzguébel J P,Glass D J,et al.Peer Reviewed:Assessing phytoremediation’s progress in the United States and Europe[J].Environmental Science & Technology,2001,35(21):446-452.
[12] 孫琴,倪吾鐘,楊肖娥,等.磷對超積累植物——東南景天生長和積累鋅的影響[J].環境科學學報,2003,23(6):818-824.
[13] 李新博,謝建治,李博文,等.鎘對紫花苜蓿不同生長期生物量的影響及飼用安全評價[J].草業學報,2009,18(5):266-269.
[14] 安志裝,王校常,施衛明,等.重金屬與營養元素交互作用的植物生理效應[J].土壤與環境,2002,11(4):392-396.
[15] 聶俊華,劉秀梅,王慶仁.Pb超富集植物對營養元素N、P、K的響應[J].生態環境,2004,13(3):306-309.
[16] 聶俊華,劉秀梅,王慶仁.營養元素N、P、K對Pb超富集植物吸收能力的影響[J].農業工程學報,2004,20(5):262-265.
[17] 李裕紅,袁建軍,陳懷宇,等.泉州灣若干海岸植物銅、鋅、鉛及氮、磷、鉀元素分布特征[J].中國生態農業學報,2008,16(3):630-634.
[18] 魯如坤.土壤農業化學分析方法[M].北京:中國農業科技出版社,2000.
[19] 王忠.植物生理學[M].北京:中國農業出版社,2000.
[20] 趙平,孫谷疇,彭少麟.植物氮素營養的生理生態學研究[J].生態科學,1998,17(2):37-42.
[21] 陸景凌.植物營養學(上冊)[M].北京:中國農業大學出版社,2003.
[22] 曲婷婷.不同低鉀耐性大豆葉片顯微結構及光合特性差異[D].沈陽:沈陽農業大學,2009.
[23] Meyers D E R,Auchterlonie G J,Webb R I,et al.Uptake and localisation of lead in the root system of Brassica juncea[J].Environmental Pollution,2008,153(2):323-332.
[24] Huang H,Li T,Tian S,et al.Role of EDTA in alleviating lead toxicity in accumulator species of Sedum alfredii H.[J].Bioresource Technology,2008,99(14):6088-6096.
[25] 楊肖娥,龍新憲,倪吾鐘,等.古老鉛鋅礦山生態型東南景天對鋅耐性及超積累特性的研究[J].植物生態學報,2001,25(6):665-672.
[26] 傅華,王玉梅,周志宇,等.施用污泥對黑麥草草坪綠地土壤理化性質和重金屬元素含量的影響[J].草業學報,2003,12(2):82-86.
[27] 李繼光,朱恩,李廷強,等.氮對鎘脅迫下東南景天根系形態及鎘積累的影響[J].環境污染與防治,2007,29(4):271-275.
[28] 李繼光,李廷強,朱恩,等.氮對超積累植物東南景天生長和鎘積累的影響[J].水土保持學報,2007,21(1):54-58.
[29] Zou T,Li T,Zhang X,et al.Lead accumulation and tolerance characteristics of Athyrium wardii (Hook.)as a potential phytostabilizer[J].Journal of Hazardous Materials,2011,186(1):683-689.
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
