利用RAPD技術構建四倍體苜蓿遺傳連鎖圖譜
2012-08-20 04:06:12劉曙娜于林清周延林吉仁花陳世茹孫娟娟么婷婷
草業學報 2012年1期
劉曙娜,于林清,周延林,吉仁花,陳世茹,孫娟娟,么婷婷
(1.內蒙古大學生命科學學院,內蒙古 呼和浩特010021;2.中國農業科學院草原研究所,內蒙古 呼和浩特010010;3.內蒙古農業大學林學院,內蒙古 呼和浩特010019;4.內蒙古農業大學農學院,內蒙古 呼和浩特010019;5.東北師范大學植被生態教育部重點實驗室,吉林 長春130024)
苜蓿(Medicago)屬豆科多年生草本植物,是世界上分布、栽培面積廣,適應性廣泛,品質優良的牧草。具有產草量高、品質優良、適口性好的特點,有很高的農業價值和商業價值。由于栽培苜蓿的同源四倍體特性使得其連鎖圖譜的構建相對困難。目前,公認的已發表四倍體苜蓿連鎖圖共有3個,Brouwer和Osborn[1]利用2個回交群體各101株個體,以單劑量位點(SDAs)方法將88個RFLP(restriction fragment length polymorphism,限制性內切酶片段長度多態性)標記構成7個連鎖群,總遺傳距離為443cM,飽和度相對較低。Julier等[2]利用168個F1雜交重組群體構建了遺傳距離為2 649cM的父本連鎖圖譜和遺傳距離為3 045cM的母本連鎖圖譜,同時利用SSR(simple sequence repeat,簡單重復序列)標記構建了遺傳距離為709cM 的整合圖譜。Musical等[3]利用 AFLP(amplified fragment length polymorphism,擴增片段長度多態性)和 RAPD(random amplified polymorphic DNA,隨機擴增多態性)標記構建了四倍體苜蓿遺傳圖譜。Eujayl等[4]于2005年利用來自紫花苜蓿(M.sativa)×黃花苜蓿(M.falcata)的F1回交群體,以EST(expressed sequence tag,表達序列標簽)-SSR、SSR(SSR的分子標記代替首例圖譜中的88個RFLP)構建了比較飽滿的遺傳圖譜,總遺傳距離為624cM。構建苜蓿分子遺傳圖譜可以進行目的基因的連鎖分析、定位重要農藝性狀相關基因位點(QTL)及分子標記輔助育種(molecular marker assisted selection,MAS)[5]。由于苜蓿遺傳圖譜共享機制不完善和我國苜蓿種質資源群體的獨特性,構建擁有自主產權的苜蓿遺傳圖譜成為首要任務。但是,目前國內尚未構建出一套完整的四倍體苜蓿遺傳連鎖圖譜。魏臻武[6]利用SSR、ISSR(inter-simple sequence repeat)和 RAPD技術構建了苜蓿的指紋圖譜。
本研究以篩選出的高抗(抗寒——秋眠級為1)黃花苜蓿與高產(每年產量——鮮重6.45kg/m2)[7]紫花苜蓿雜交產生F1代個體,并且由F1代個體自交構建F2群體。對F1的親本紫花、黃花苜蓿和F2個體DNA樣本分別進行RAPD[8]分子標記的遺傳作圖研究。旨在優化苜蓿分子遺傳學實驗體系,構建苜蓿遺傳連鎖框架圖,為進一步構建飽和的苜蓿遺傳連鎖圖譜及開展苜蓿分子育種研究奠定基礎。
1 材料與方法
1.1 植物材料
2007年選擇早期收集的黃花苜蓿與紫花苜蓿的特殊類型遺傳材料,包括國內、美國農業部和俄羅斯共計25份材料(均為2代以上選育的四倍體材料)。對所選的18份紫花苜蓿、7份黃花苜蓿進行正、反交產生了大量的F1代。2008年春在中國農業科學院草原研究所呼和浩特市南郊實驗場對雜交成功的F1育苗并人工套袋自交,于2009年建立了F2群體。2010年對雜交成功的94個單株雜交子代進行形態與農藝指標的檢測。
1.2 DNA提取與純化
1.2.1 基因組DNA提取 對苜蓿親本(本研究中均為高產紫花苜蓿、高抗黃花苜蓿)、F1和F2代個體葉片分別進行取樣,并存儲于-80℃的超低溫冰箱中待用。本實驗采用植物基因組提取試劑盒(由上海生物工程有限公司提供)進行DNA提取。
1.2.2 基因組DNA檢測 紫外分光光度法測定DNA的濃度、純度,計算樣品濃度:DNA濃度(μg/μL)=OD260×N×50/1000[9],式中,OD260為核酸吸光度,N 為樣品稀釋倍數。
電泳檢測DNA:將提取的DNA在0.8%的瓊脂糖凝膠(含GoodView核酸染料)上電泳,檢測其完整性和片斷大小。
1.3 RAPD實驗體系
反應在96孔PCR板上進行,總體積為20μL,其中包括模板DNA 1μL(大約20ng);50μmol/L的引物1 μL;2×Taq Master Mix 10μL;ddH2O 8μL。擴增條件為94℃下預變性3min;進入循環,94℃下變性30s;36℃下復性30s;72℃下延伸60s,共40個循環;最后在72℃下延伸10min,結束后于4℃下保存。
擴增結束后,取下樣品,在制備好的1.5%瓊脂糖膠(含GoodView核酸染料)上點樣。在1×TBE電泳緩沖液中電泳,功率為100W。電泳時間為45min。電泳結束后取出膠在UVI凝膠成像系統(英國UVI公司Fire-Reader系列)下觀察、拍照記錄。
實驗用于RAPD的隨機引物共192個,購自上海Sangon公司。
1.4 苜蓿RAPD引物篩選
用F1親本及隨機抽取的F2個體的DNA樣品篩選在親本中呈多態性而在子代中呈分離的擴增譜帶的引物。
1.5 RAPD擴增作圖及數據采集
利用篩選的引物對包括F1的親本、F1及F2個體共97個樣本的DNA進行擴增反應。記錄在親本中存在多態性而在F2代個體中發生分離的譜帶,某一譜帶出現的個體在該位點的基因型記作“1”;該譜帶不出現的個體在該位點的基因型記作“0”;有些帶型不易確定連同缺失的個體,一同按缺失數據處理記作“-”。按通用格式將所有數據輸入計算機保存成文本文件備用。
1.6 RAPD標記的分離檢測、連鎖分析與圖譜構建
將原始數據文件轉換成符合 MAPMAKER 3.0(For PC)作圖軟件中要求的作圖數據文件,然后采用MAPMAKER 3.0軟件并結合JionMap 4.0軟件在計算機上構建連鎖圖譜[10]。通過兩點連鎖分析,計算所有成對座位的重組率和最大LOD值,推測可能的連鎖群。根據需要,不同的連鎖群可以定義不同的LOD值,以達到最佳的分群效果。然后對每個連鎖群中的標記作多點分析,并采用Kosambi函數,將重組率轉換成圖距單位——厘摩(centimorgan,cM)。分析模型選用自交(F2)類型。經過連鎖分析,最后得到連鎖框架圖。
2 結果與分析
2.1 RAPD引物篩選
用親本DNA和F2代個體DNA對192個隨機引物進行篩選,共獲得在親本中呈多態性,在F2代個體中有分離的隨機引物72個,有120個引物(占62.50%)沒有檢測出多態性標記。引物入選率為37.50%。
2.2 親本的多態性檢測
從隨機引物篩選的結果中發現有37.50%的引物揭示了親本間的遺傳多態性,表明兩親本在基因組水平上存在一定的差異。
2.3 RAPD標記分析
用篩選的72個引物對F1雙親、F1及94個F2群體樣本DNA進行擴增(圖1),共記錄了74個RAPD標記的分離。F2中多態分離標記來自母本黃花苜蓿的有29個(占39.19%),來自父本紫花苜蓿的有45個(占60.81%),表明F2群體基本上分別來自雙親的50%,略偏父本。
參與連鎖分析的RAPD分子標記共計74個。其中,51個標記位點分別定位在8個連鎖群上(表1),覆蓋長度為1 261.5cM,平均標記數為6.38個,2個標記間平均圖距為24.73cM,另有23個標記未與這8個連鎖群連鎖。標記位點較多的連鎖群為LG1、LG2、LG3和LG8,分別包含18,8,5和5個標記,標記數最少的連鎖群只有3個標記。連鎖群LG1遺傳距離最長,為461.2cM,遺傳距離最短的為51.3cM,平均圖距最大的連鎖群為LG3,間距為38.16cM,平均圖距最小的連鎖群為LG4,間距為14.23cM。來源于紫花苜蓿的45個多態RAPD標記中有32個標記被分配到8個連鎖群上,占43.24%。有13個標記未被連鎖到任何一個連鎖群上;來源于黃花苜蓿的29個多態RAPD標記中有19個標記被分配到8個連鎖群上,占25.68%。有10個標記未被連鎖到任何一個連鎖群上。
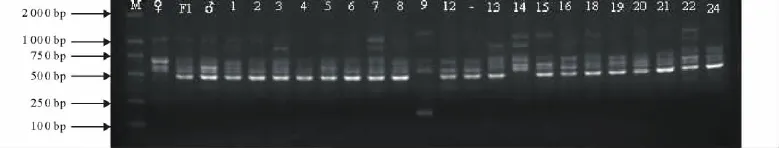
圖1 S1013對親本及個別F2個體DNA電泳圖Fig.1 PCR amplification patterns by primer S1013in parents and F2
2.4 連鎖分析與圖譜構建
通過對每個連鎖群中的標記作多點分析,并將重組率轉換成圖距單位(cM)。經過連鎖分析,共得到8條連鎖群(圖2),其中,每條連鎖群左側給出了2個標記位點間的圖距(cM),右側給出了呈多態性的標記名稱。
由于作圖軟件的局限性和總標記數目太少,本研究所獲得的連鎖標記不多。隨著實驗條件的逐步完善,可以預期,隨著標記數目的增加,連鎖標記會越來越多,遺傳圖譜越來越密集。
本研究中,定位到圖譜上的分離標記共51個,來自于49個RAPD引物(表2)。其中,引物L16、S43均有2個分離標記。
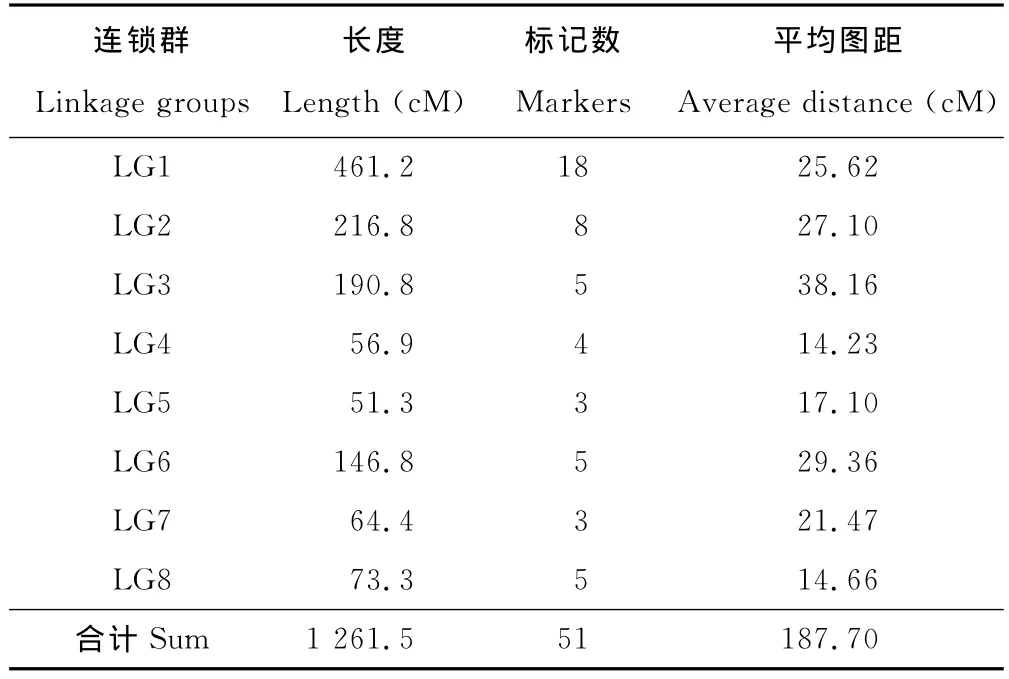
表1 分子標記在各連鎖群的分布情況Table 1 Molecular marker in the distribution of a linkage groups
3 討論
本研究以F2群體作為作圖群體,利用RAPD技術構建了四倍體苜蓿的遺傳圖譜。以期為苜蓿遺傳育種工作提供參考與基礎材料。
建立一個完善的遺傳作圖群體直接關系到圖譜構建的質量高低。由于苜蓿自身的細胞學特性、遺傳研究水平和現有構圖技術限制,在短時間內不可能像農作物那樣建立高世代作圖群體。目前真正比較理想的作圖群體比較少,對親本的一些基本信息還了解不多,在作圖中將會帶來很多問題[2]。用于苜蓿遺傳圖譜構建的作圖群體,主要有下列幾種類型:F2群體、回交群體、F1群體等。在國外,目前公布的3個四倍體苜蓿圖譜中的作圖群體分別是回交群體(BC1為基礎)[1]和F1雜交重組群體[2,4]。在國內,張麗芳[11]以2個差異較大的蒺藜苜蓿親本W5160和Jemalong進行雜交,獲得雜交組合W5160×Jemalong,進而構建了蒺藜苜蓿的F2群體。目前國內還未見關于四倍體栽培苜蓿群體構建的報道,今后有必要加強相關的研究[12]。而在構建遺傳圖譜中采用的分子標記主要包括RFLP、RAPD、AFLP、STS(sequence tagged sites,序列標志位點)、SSR 等。各種方法均各具優缺點。RAPD技術是建立在PCR基礎上的一種可對整個未知序列的基因組進行多態性分析的分子標記技術[13]。相比之下,RAPD技術方法簡便、高效、成本低,得到了廣泛的應用。目前已被分子遺傳學家應用到各個學科、領域,如品種鑒定[14-16]、遺傳多樣性分析[17-20]、遺傳連鎖分析和基因定位、分子標記輔助育種[21]等。目前已發表的多個苜蓿圖譜由于材料與所選分子標記的不同還不能重合。隨著各種遺傳標記方法的逐步完善和發展,目前已經進入綜合應用各種遺傳標記進行基因組研究的新階段。作圖群體的大小直接影響到作圖的精度和應用范圍。從已發表的圖譜來看,大多數所采用的群體樣本數在100以內,少數研究者采用300個以上。本研究中采用了94個樣本可檢測到的重組最小圖距為3.2cM,表明在樣本數量上可以滿足構建連鎖框架圖的需要。本研究在構建遺傳圖譜時選擇了 MAPMAKER/EXP(3.0)與JionMap 4.0軟件來進行分群,MAPMAKER/EXP(3.0)在作物、蔬菜中已被廣泛應用,但在牧草中還尚未廣泛應用。分群過程中可以根據分群效果選用適宜的LOD值,以期獲得最佳分群效果。
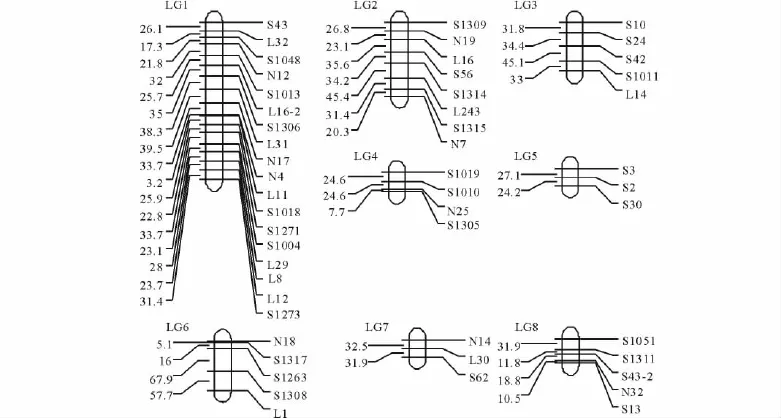
圖2 苜蓿連鎖框架圖Fig.2 RAPD map of tetraploid alfalfa
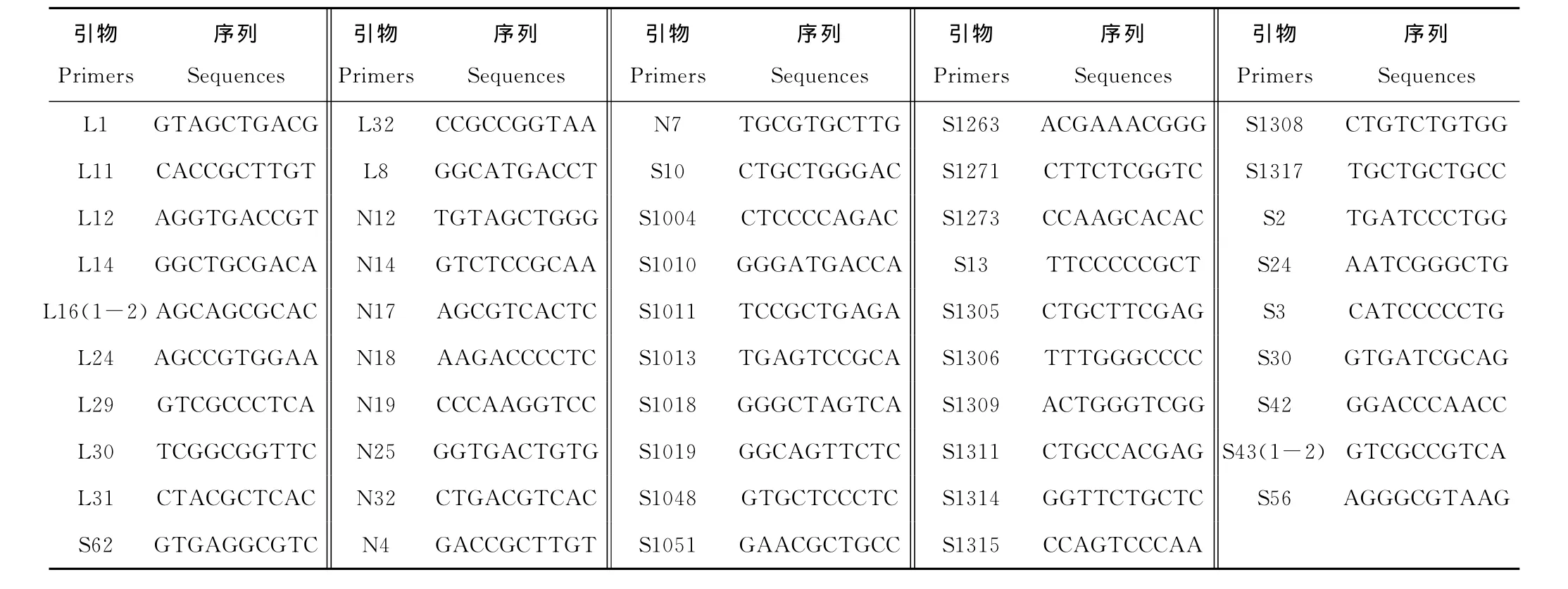
表2 49個RAPD引物序列Table 2 Sequences of 49RAPD primers
由于在過去缺乏對苜蓿經典遺傳學和細胞遺傳學等方面的研究,目前大多數已構建的遺傳連鎖圖譜僅僅是一個框架圖,大都沒有和染色體相對應,構建的圖譜離應用還有一段距離;因此在今后的一段時間內,在開展分子標記研究的同時必須加強一些基礎研究。
[1] Brouwer D J,Osborn T C.A molecular marker linkage map of tetraploid alfalfa(Medicago sativa L.)[J].Theoretical and Applied Genetics,1999,99:1194-1200.
[2] Julier B,Flajoulot S,Barre P.Construction of two genetic linkage maps in cultivated tetraploid alfalfa(Medicago sativa L.)using microsatellite and AFLP markers[J].BMC Plant Biology,2003,3:1-19.
[3] Musical J M,Lowe K F,Mackie J M,et al.DNA maker linked to yield,yield components,and morphological traits in autotetraploid lucerne(Medicago sativa L.)[J].Australian Journal of Agricultural Research,2006,57:801-810.
[4] Eujayl I,Sledge M K,Wang L,et al.Medicago truncatula EST-SSRs reveal cross-species genetic markers for Medicago spp.[J].Theoretical and Applied Genetics,2004,108:414-422.
[5] 王曉娟,孫月華,楊曉莉,等.苜蓿遺傳圖譜構建及其應用[J].草業學報,2008,17(3):119-127.
[6] 魏臻武.利用SSR、ISSR和RAPD技術構建苜蓿基因組DNA指紋圖譜[J].草業學報,2004,13(3):62-67.
[7] 于林清.苜蓿種質資源系統評價與遺傳多樣性分析[D].呼和浩特:內蒙古農業大學,2009.
[8] Wi1liams J G K,Kubelik A R,Livak K J,et al.DNA poly morphisms amplified by arbitrary primers are useful as genetic markers[J].Nucleic Acids Research,1990,18:6531-6535.
[9] 周延清.DNA分子標記技術在植物研究中的應用[M].北京:化學工業出版社,2005:4.
[10] 邢光南,趙團結,蓋鈞鎰.關于Mapmaker/Exp遺傳作圖中標記分群和排序操作技術的討論[J].作物學報,2008,34(2):217-223.
[11] 張麗芳.蒺藜苜蓿作圖群體的構建及其群體遺傳分析[D].蘭州:甘肅農業大學,2007.
[12] 何慶元,玉永雄.苜蓿作圖群體構建研究進展[A].第三屆苜蓿發展大會[C].北京:中國畜牧業協會,2010:23-25.
[13] 李惠英,婁燕宏,胡濤,等.中國高羊茅種質資源遺傳多樣性的RAPD分析[J].草業學報,2010,19(6):208-214.
[14] 林楠,翁頻,沈明山.RAPD鑒定遠緣雜交牧草蔗新品系94~42[J].廈門大學學報(自然科學版),2003,42(2):238-241.
[15] 高捍東,黃寶龍.板栗主要栽培品種的分子鑒別[J].林業科學,2001,37(1):64-71.
[16] Echt C S,Kidwell K K,Knapp S J,et al.Linkage mapping in diploid alfalfa(Medicago sativa)[J].Genome,1994,37:61-71.
[17] Chrochemore M L,Huyghe C,Ecalle C,et al.Structuration of alfalfa genetic diversity using agronomic and morphological characteristic-Relationship with RAPD markers[J].Agronomie,1998,18:79-94.
[18] 李擁軍,蘇加楷.苜蓿地方品種遺傳多樣性的研究:RAPD標記[J].草地學報,1998,6(2):105-114.
[19] 魏臻武,符昕,耿小麗,等.苜蓿遺傳多樣性和親緣關系的SSR和ISSR分析[J].草地學報,2007,15(2):118-123.
[20] 梁慧敏.不同居群狗牙根RAPD分析[J].草業學報,2010,19(1):258-262.
[21] 孟亞雄,仲軍,馬小樂,等.甘肅省小麥條銹病主要流行小種的RAPD分析及Sull-4小種SCAR標記建立[J].草業學報,2010,19(6):180-186.
