東方山羊豆脫水蛋白基因的克隆及初步分析
2012-08-20 04:06:14李玉坤王學敏王贊李俊VladimirNikolay孫桂枝高洪文
草業學報 2012年1期
關鍵詞:植物
李玉坤,王學敏,王贊,李俊,Vladimir C,Nikolay D,孫桂枝,高洪文*
(1.中國農業科學院北京畜牧獸醫研究所,北京100193;2.俄羅斯瓦維洛夫全俄植物栽培研究所,圣彼得堡190000)
干旱、高鹽、低溫等非生物脅迫嚴重影響了植物的生存和產量,它們均可造成植物細胞缺水,產生干旱信號,誘導植物的抗旱反應。晚期胚胎發生豐富蛋白(late embryogenesis abundant protein,LEA)是一種脫水保護劑,能夠在水分脅迫下保護生物大分子。LEA蛋白可分為許多不同的家族[1,2],脫水蛋白(dehydrin,DHN)屬于LEAⅡ或LEA D-11蛋白家族[2]。DHN可在胚胎發生晚期或者在干旱、低溫、鹽脅迫和ABA的誘導下表達[3]。脫水蛋白一級結構保守性強,具有K,S和Y等3個高度保守區域,通常以YSK的順序結合在一起[3]。Y片段為(V/T)DEYGNP,位于N末端。K片段(EKKGIMDKIKEKLPG)由15個氨基酸組成。S片段是一個可磷酸化的絲氨酸簇,一般位于K片段之間,或緊跟K片段。高度極性的φ片段的存在增加了極性氨基酸片段的比例,加強了脫水蛋白結構的親水性。由于K片段和φ片段的存在,脫水蛋白能夠通過形成一個保護層而保護變性的高分子,在各種組織中都起到穩定細胞結構的作用[3,4]。
Koag等[5]從玉米(Zea mays)中分離純化得到DHN1,證實在受到脅迫的植物中,脫水蛋白在穩定液泡和細胞膜的結構中起到非常重要的作用。在水稻(Oryza sativa)中,已報道了多個脅迫誘導的脫水蛋白,其中RAB21/16A,RAB16B,16C,16D,RAB25和 WSI724已經證實是滲透脅迫誘導的[6-9]。此外,LEA 蛋白在植物和酵母中超表達可以提高對滲透脅迫的抵抗力[10-12]。
東方山羊豆(Galega orientalis)作為一種新型優良豆科牧草,在我國主要種植在西北地區。東方山羊豆品質好、產量高、抗病抗逆性強、使用年限長[13],與其他牧草資源相比,東方山羊豆目前的相關研究較少[14]。中國農業科學院北京畜牧獸醫研究所牧草資源研究室自2007年以來對引進東方山羊豆的抗逆基因開展了研究,目前已經取得一定進展。李鑫等[15]從東方山羊豆中分離得到液泡膜Na+/H+逆向轉運蛋白基因,推測該基因在轉錄水平的調節可能是決定東方山羊豆耐鹽能力的重要因子。Chen等[16]克隆了東方山羊豆的RAV基因,實時熒光定量PCR(real-time PCR)分析表明,GoRAV 基因受外源脫落酸(abscisic acid,ABA)、干旱、低溫和鹽脅迫誘導,證明該基因在依賴ABA信號途徑的抗逆性調控中起到一定的作用。
目前,抗逆基因工程主要集中在逆境條件下表達的某些基因和抗逆代謝過程中某些酶的研究[17]。隨著生物技術的發展和廣泛應用,克隆植物抗逆相關基因并利用轉基因技術將一些與抗性密切相關的外源目的基因導入目標植物中來提高其抗逆性是解決逆境脅迫的一條有效途徑[18]。本研究首次從東方山羊豆中克隆DHN基因,初步探索了DHN基因在東方山羊豆抗逆性方面的作用,為深入研究該種牧草的抗逆機理,進行抗逆性改良奠定了一定基礎。
1 材料與方法
1.1 實驗材料
1.1.1 植物材料 東方山羊豆為中國農業科學院北京畜牧獸醫研究所牧草資源研究室提供。
1.1.2 藥品與試劑 Trizol試劑為Invitrogen公司產品,cDNA Synthesis Kit為Promega公司產品,SMARTTMRACE cDNA Amplification Kit為 Clontech公司產品,克隆載體pMD18T、Taq酶、限制性內切酶、Alkaline Phosphatase、T4DNA連接酶均為TaKaRa公司產品。
1.2 實驗方法
1.2.1 材料的處理與總RNA的提取 于2009年7月開始植物材料的處理,首先將東方山羊豆的種子用氯氣消毒24h,播種于鋪有濾紙的培養皿中,在光照培養箱(溫度為24℃,光照時間為12h/d)中培養至種子發芽。然后將發芽后的種子移至蛭石與珍珠巖比例為3∶1的花盆中繼續培養30d。幼苗用0.25mol/L NaCl溶液處理2,6,10h,Trizol法提取總RNA后用于克隆DHN 基因的全長cDNA序列[19]。再用0.25mol/L NaCl溶液、20%PEG(聚乙二醇)溶液、0.1×10-3mol/L ABA溶液分別處理幼苗0,2,4,8,12和24h,分別提取總 RNA,用于 Real-time PCR分析。
1.2.2 DHN 基因3′末端序列、5′末端序列和全長cDNA序列的克隆 用Trizol法分別提取0.25mol/L NaCl溶液處理2,6,10h后的幼葉總RNA后,按照質量比為1∶1∶1混合,用混合RNA為模板反轉錄成cDNA。根據中國農業科學院北京畜牧獸醫研究所牧草資源研究室通過構建SSH cDNA文庫得到的DHN基因的EST序列,用Primer Premier 5.0引物設計軟件分別設計3′RACE和5′RACE特異性引物,3′RACE:GGTTATGGAACAACTGGGTATGGTGG;5′RACE:CCAGTGCTTCCAGTTCCACCATACCC。根據 SMARTTMRACE cDNA Amplification Kit說明書分別PCR擴增出3′末端和5′末端序列,分別測序后,結合EST序列用DNAMAN 5.0軟件拼接出全長cDNA序列,找到開放閱讀框ORF,在ORF兩端分別設計全長引物,DHN-ORF-F:CAATGTCTCAGTATAATCAAGGTCA;DHN-ORF-R:AGAGATACTATATGATCTAGTGTCCAG。經 PCR 擴增,測序后得到DHN基因全長cDNA序列。
核酸及氨基酸序列分析、開放閱讀框ORF的查找和翻譯用DNAStar 6.13軟件進行分析,利用ScanProsite服務器(http://www.expasy.org/tools/scanprosite/)對 DHN蛋白進行功能預測。從 GeneBank上下載其他植物中的18種DHN蛋白,利用MEGA4軟件Neighbor-Joining法構建系統進化樹。
1.2.3 Real-time PCR檢測DHN基因的組織表達特異性 分別提取東方山羊豆的根、莖、葉總RNA,反轉錄為cDNA。根據東方山羊豆DHN基因全長cDNA序列設計特異性引物(預期擴增片段大小為270bp),DHNRT-F:GGTCAATACGGTCAACAAACACG;DHN-RT-R:CACCTGTACCTGTCTGGGTTCC。以 Actin 基因做內參,設計特異性引物(預期擴增片段大小為200bp),Actin-RT-F:GGACAAGTTATCACCATCGG;Actin-RT-R:TCAGGAATACCTGGAAACATAG。參考TaKaRa公司的SYBR Premix Ex TaqTM試劑盒說明書,使用 ABI PRISM 7500Real-time PCR System(ABI,USA),采用兩步法進行 Real-time PCR擴增,第1步:95℃預變性30s;第2步:95℃5s,60℃34s,40個循環。每個cDNA樣品做3次重復。
根據得到的Ct值,利用Kenneth和Thomas[20]報道的2-△△Ct方法,分別計算DHN基因在根、莖、葉中的表達量。把根中表達量設為對照,莖、葉分別設為2個不同處理,利用公式:

分別計算出根、莖、葉的2-△△Ct(表達水平)值,式中,CtTarget為目標基因達到設定閾值所經歷的循環數;CtActin為內參基因達到設定閾值所經歷的循環數;(CtTarget-CtActin)處理為處理樣品中目標基因經Actin校正后的循環數;(CtTarget-CtActin)對照為對照樣品中目標基因經Actin校正后的循環數;△△Ct為處理樣品中目標基因與對照樣品中目標基因循環數的差值。利用Excel繪出柱形圖。
1.2.4 Real-time PCR檢測DHN 基因在不同逆境脅迫下的表達量 按照1.2.1中用于Real-time PCR分析的材料處理方法處理東方山羊豆幼苗,分別提取葉片總RNA,反轉錄為cDNA。同1.2.3的方法,進行不同逆境脅迫下的表達量分析。每個cDNA樣品做3次重復。
將0h設為對照,2,4,8,12和24h分別設為5個不同處理,按照1.2.3的方法,分別計算出2-△△Ct值,利用Excel繪出柱形圖。
1.2.5 構建植物表達載體 設計帶有BglII酶切位點的引物,DHN-F:GAAGATCTAATGTCTCAGTATAATCAAGGTC;DHN-R:GAAGATCTGTGTCCAGTACAAGATCCAG。以pMD18T-DHN重組質粒為模板,擴增出帶有酶切位點的DHN基因的ORF。用BglII限制性內切酶酶切擴增產物和pCAMBIA1302植物表達載體,對酶切后的p1302載體進行去磷酸化后,用T4DNA連接酶4℃連接過夜,轉化大腸桿菌,涂布含有卡那霉素的LB(Luria-Bertani培養基)平板,經抗生素篩選后挑菌,搖菌,抽提質粒,經BglII酶切鑒定為陽性克隆的,送北京三博遠志公司測序。
2 結果與分析
2.1 DHN基因全長cDNA的克隆與分析
以東方山羊豆鹽脅迫cDNA為模板,分別利用3′RACE和5′RACE特異性引物擴增出3′末端和5′末端片段,其中3′末端片段經測序為397bp(圖1A),5′末端片段經測序為813bp(圖1B)。利用DNAMAN 5.0軟件拼接后的全長cDNA為1 169bp。利用DHN-ORF特異性引物,擴增出全長cDNA的ORF經測序為843bp(圖1C)。將該基因命名為GoDHN,NCBI登錄號為HM777019。

圖1 DHN基因的3′末端片段、5′末端片段和全長ORF擴增結果Fig.1 Amplification results of 3′RACE,5′RACE and full-length ORF of DHNgene

圖2 DHN基因的核苷酸序列和氨基酸序列Fig.2 The nucleotide sequence and amino acid sequence of DHNgene
利用DNAStar 6.13軟件對DHN基因cDNA序列進行分析,開放閱讀框編碼281個氨基酸,所編碼的蛋白質分子量為28.71kDa。通過ScanProsite服務器對DHN蛋白的氨基酸序列分析,該氨基酸序列包含1個Y片段,3個K片段(圖2)。
用MEGA 4軟件構建系統進化樹(圖3),分析結果顯示,GoDHN蛋白與豆科植物親緣關系較近,與禾本科和十字花科等的親緣關系較遠。其中與豌豆PsDHN蛋白親緣關系最近,其次是大豆GmMAT1和GmDHN5蛋白,均屬于YKn亞型,但與豆科植物紫花苜蓿MsCAS15A蛋白和大豆GmSRC1蛋白親緣關系較遠,這2個蛋白屬于KnS亞型。
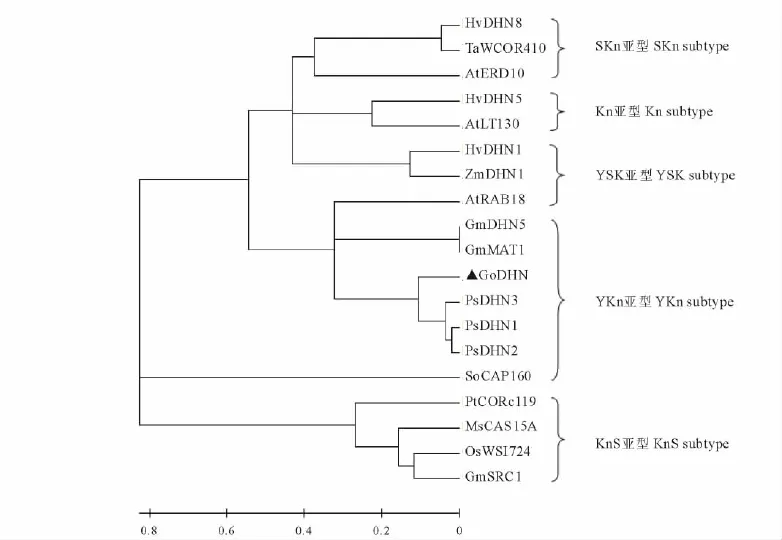
圖3 東方山羊豆DHN蛋白與其他植物中18種DHN蛋白的系統進化樹Fig.3 The phylogenetic tree of dehydrins in G.orientalis and another eighteen plants
2.2 DHN基因的組織特異性表達分析
根據1.2.3中的公式,計算DHN基因在東方山羊豆不同組織中的相對表達量(圖4)。結果表明,GoDHN在根中表達量最少,莖中最多,葉中次之。以東方山羊豆根中DHN基因的表達量為對照,莖中的表達量為根中的6倍,葉中表達量為根中的5.5倍。
2.3 DHN基因在不同逆境脅迫下的表達分析
根據1.2.4中的公式,計算了DHN基因在分別受到ABA、NaCl和PEG脅迫0,2,4,8,12和24 h的相對表達量(圖5)。結果顯示,DHN基因在ABA脅迫誘導下,4h時表達量達到一個小高峰,然后下降,在24h誘導后表達量猛增到4h表達量的2倍(圖5A)。在NaCl的脅迫誘導下,DHN基因的表達量逐步增加,在24h達到最高峰,為對照(0 h)的6 500倍(圖5B)。在PEG誘導12h后,DHN基因的表達量達到一個最高峰,為對照(0h)的42 000倍,誘導24h后,表達量相對12h時有所下降,但仍然是對照(0h)的35 000倍(圖5C)。
2.4 植物表達載體的構建及鑒定
用BglII限制性內切酶分別酶切帶有BglII酶切位點的DHN基因ORF和pCAMBIA1302載體后,經過連接、轉化,挑取菌落,抽提質粒,經BglII酶切鑒定,得到與預期大小一致的目的片段(圖6)。鑒定為陽性的質粒經測序序列完全正確,表明植物表達載體pCAMBIA1302-DHN構建成功。
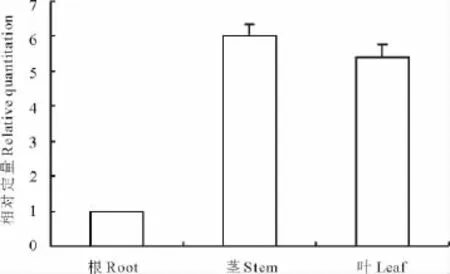
圖4 DHN基因在東方山羊豆的不同組織中的相對表達量Fig.4 The relative expression of DHNgene in different tissues of G.orientalis
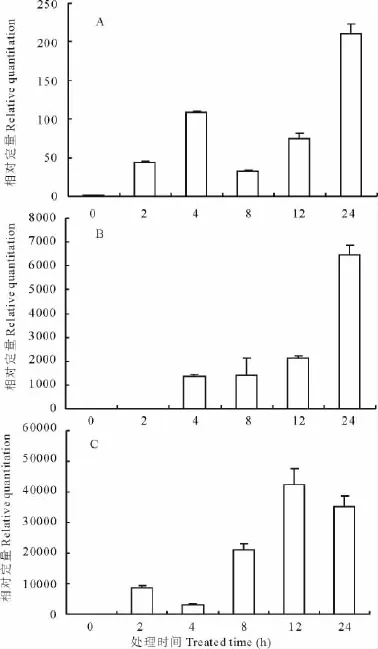
圖5 DHN基因在受到ABA、NaCl和PEG誘導后的相對表達量Fig.5 The relative expression of DHNgene induced by ABA,NaCl and PEG
3 討論
本研究得到的GoDHN基因的氨基酸序列中(圖2),在 N末端有一個 Y片段(VDQYGNP),Close[3]認為該片段的存在與DHN基因在植物和細菌中的核定位有關。在該氨基酸序列的中間部位和C末端包含有3個 K片段(EE/D/KKGIMDKIKEKIPG),Soulages等[19]通過二級結構預測證明K片段可能形成α螺旋結構,參與脫水蛋白的親水或疏水作用。純化的玉米脫水蛋白G50中15%的氨基酸形成了α螺旋結構,但G50中沒有任何疏水位點,卻能參與疏水作用,推測K片段形成的α螺旋結構,賦予了G50參與疏水相互作用的潛力[21]。脫水蛋白通過K片段可能參與部分變性蛋白質及膜的疏水相互作用來防止蛋白質和膜進一步變性[19]。在某些植物的脫水蛋白中,如擬南芥(Arabidopsisthaliana)RAB18,還含有一個 S保守片段,S片段是一個可磷酸化的絲氨酸簇,該片段的磷酸化可能也與核定位信號有關,可以參與脫水蛋白的入核過程[3]。
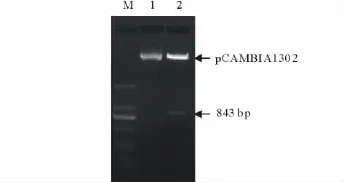
圖6 酶切鑒定Fig.6 Indentification of enzyme digestion
通過構建系統進化樹(圖3),將脫水蛋白分為5類[4]。結果表明,屬于同一亞型的脫水蛋白親緣關系有的近有的遠,如GoDHN蛋白與菠菜(Spinacia oleracea)SoCAP160蛋白雖然同屬于YKn亞型,但是親緣關系較遠;與豌豆(Pisum sativum)PsDHN蛋白親緣關系最近,而且同屬于YKn亞型。與擬南芥At-RAB18蛋白親緣關系雖然較近卻不屬于同一亞型。說明在脫水蛋白的進化過程中,分離較晚的蛋白有可能進化為不同的亞型,而較早分離的蛋白可能源于同一亞型,并且在進化過程中,一直保持不變。
YKn亞型脫水蛋白是一類酸性蛋白,對這一類蛋白的抗性研究主要集中在干旱和冷害環境脅迫下,如大豆(Glycine max)Mat1編碼一個Y2K型脫水蛋白,主要由干旱脫水誘導產生,而在ABA誘導下并無明顯表達[22]。菠菜CAP160編碼一個YK型脫水蛋白,主要受低溫誘導,在轉基因煙草中的高效表達可以提高其抗寒性[23]。桃樹 (Prunus persica)PpDhn1編碼一個Y2K2型脫水蛋白,也在低溫誘導時大量表達[24]。以上結果表明大部分YK型脫水蛋白在抗旱和耐寒方面可能發揮重要作用。本實驗增加了鹽脅迫方面的研究,通過模擬逆境處理后(圖5),GoDHN基因在NaCl、PEG和ABA脅迫條件下表達均上調,其中NaCl和PEG模擬的鹽脅迫和干旱環境,誘導GoDHN基因表達量成千上萬倍的提高,暗示GoDHN脫水蛋白與植物的抗鹽和抗旱性密切相關。ABA誘導GoDHN基因的表達量雖然低于NaCl和PEG,但是與對照相比仍然提高了200倍,表明外源激素ABA的存在也可以誘導脫水蛋白的表達量上調。
脫水蛋白參與細胞的滲透調節過程,可以提高植物的抗旱和抗鹽性[25]。向日葵(Helianthus annuus)脫水蛋白基因HaDhn1和HaDhn2在干旱脅迫下的轉錄活性顯著高于供水條件的轉錄活性,耐旱品種累積的轉錄物明顯高于敏感品種,并且其累積量與耐旱品種芽細胞膨壓的維持和干旱的適應性呈正相關[26]。樺樹(Betula platyphylla)脫水蛋白Peudhn1,在未脅迫的植株中僅有少量表達,而在PEG6000和NaCl處理后,該蛋白在根和葉中大量積累,并在復水后持續積累[27]。在模擬鹽脅迫下誘導24h后,GoDHN基因的表達量是對照的6 500倍,顯著高于對照;在模擬干旱脅迫下誘導12h后,GoDHN基因的表達量是對照的42 000倍,誘導24h后,表達量雖然有所下降,但仍然顯著高于對照(圖5)。GoDHN可能通過參與細胞內的滲透調節過程,來提高東方山羊豆的抗旱耐鹽能力。而高強度的誘導表達可能暗示著東方山羊豆具有非常強的抗旱耐鹽能力。
構建植物表達載體是有一定策略的,構建載體時目的基因與載體的連接可分為同源粘末端的連接、平末端的連接、定向克隆、接頭連接和同聚物加尾連接等[28]。本研究中,為了消除載體自身連接,采用堿性磷酸酶(alkaline phosphatase,CIAP)對載體進行去磷酸化。最后得到植物表達載體的重組質粒及菌株并通過酶切鑒定,確定連接正確。
目前對脫水蛋白的基因表達、結構和功能的研究已取得了很大的進展,但其具體的分子保護機制大多數只限于推測。例如,在逆境下脫水蛋白具有穩定細胞膜和保護蛋白質的作用雖然已被普遍認同,但其在植物體內具體是怎樣發揮作用的并不清楚。尤其是在牧草領域,對于脫水蛋白的研究很少。本實驗為進一步研究東方山羊豆脫水蛋白在逆境條件下的表達調控規律和生理生化機制奠定了一定理論基礎,也為牧草抗逆性改良提供了候選基因。
[1] Allagulova C R,Gimalov F R,Shakirova F M,et al.The plant dehydrins:Structure and putative functions[J].Biochemistry(Moscow),2003,68(9):945-951.
[2] Dure L III,Crouch M,Harada J,et al.Common amino acid sequence domains among the LEA polypeptides of higher plants[J].Plant Molecular Biology,1989,12:475-486.
[3] Close T J.Dehydrins:emergence of a biochemical role of a family of plant dehydration proteins[J].Physiologia Plantarum,1996,97(4):795-803.
[4] Close T J.Dehydrins:A commonalty in the response of plants to dehydration and low temperature[J].Physiologia Plantarum,1997,100(2):291-296.
[5] Koag M,Fenton R D,Wilkens S,et al.The binding of maize DHN1to lipid vesicles.Gain of structure and lipid specificity[J].Plant Physiology,2003,131:309-316.
[6] Kusano T,Aguan K,Abe M,et al.Nucleotide sequence of a rice rab16homologue gene[J].Plant Molecular Biology,1992,18:127-129.
[7] Mundy J,Chua N H.Abscisic acid and water-stress induce the expression of a novel rice gene[J].Embo Journal,1988,7(8):2279-2286.
[8] Takahashi R,Joshee N,Kitagawa Y.Induction of chilling resistance by water stress,and cDNA sequence analysis and expression of water stress-regulated genes in rice[J].Plant Molecular Biology,1994,26(1):339-352.
[9] Yamaguchi-Shinozaki K,Mundy J,Chua N H.Four tightly linked rab genes are differentially expressed in rice[J].Plant Molecular Biology,1990,14(1):29-39.
[10] Duan X,Wang B,Hong B,et al.Expression of a late embryogenesis abundant protein gene,HVA1,from barely confers tolerance to water deficit and salt stress in transgenic rice[J].Plant Physiology,1996,110:249-257.
[11] Imai R,Chang L,Ohta A,et al.A LEA-class gene of tomato confers salt and freezing tolerance when expressed in Saccharomyces cerevisiae[J].Gene,1996,170(2):243-248.
[12] Zhang L,Ohta A,Takagi M,et al.Expression of plant group 2and group 3lea genes in Saccharomyces cerevisiae revealed functional divergence among LEA proteins[J].Journal of Biochemistry,2000,127(4):611-616.
[13] 張清斌,楊志忠,賈納提,等.東方山羊豆引種研究初報[J].中國草地,2001,23(4):17-20.
[14] 沈禹穎,張自和,謝田玲.優良豆科牧草——東方山羊豆的研究與利用[J].草業學報,2003,12(6):105-109.
[15] 李鑫,王贊,王學敏,等.東方山羊豆液泡膜Na+/H+逆向轉運蛋白基因的克隆與分析[J].植物生理學通訊,2009,45(5):444-448.
[16] Chen X F,Wang Z,Wang X M,et al.Isolation and characterization of GoRAV,a novel gene encoding a RAV-type protein in Galegae orientalis[J].Genes & Genetic Systems,2009,84(2):101-109.
[17] 梁哲,姜三杰,未麗,等.三葉草基因工程研究進展[J].草業學報,2009,18(2):205-211.
[18] 周玲玲,繆建錕,祝建波,等.大葉補血草Na+/H+逆向轉運蛋白基因的克隆及序列分析[J].草業學報,2009,18(5):176-183.
[19] Soulages J L,Kim K,Arrese E L.Conformation of a group 2late embryogenesis abundant protein fromsoybean.Evidence of Poly(L-Proline)-typeⅡstructure[J].Plant Physiology,2003,131(3):963-975.
[20] Kenneth J K,Thomas D S.Analysis of relative gene expression data using real-time quantitutive PCR and the 2-ΔΔCtmethod[J].Methods,2001,25:402-408.
[21] Close T J,Kortt A A,Chandler P M.A cDNA-based comparison of dehydration-induced proteins(dehydrins)in barley and corn[J].Plant Molecular Biology,1989,13(1):95-108.
[22] Whitsitt M S,Collins R G,Mullet J E.Modulation of dehydration tolerance in soybean seedlings[J].Plant Physiology,1997,114(3):917-925.
[23] Kaye C,Neven L,Hofig A,et al.Characterization of a gene for spinach CAP160and expression of two spinach cold-acclimation proteins in tobacco[J].Plant Physiology,1998,116(4):1367-1377.
[24] Artlip T S,Callahan A M,Bassett C L,et al.Seasonal expression of a dehydrin gene in sibling deciduous and evergreen genotypes of peach(Prunus persica[L.]Batsch)[J].Plant Molecular Biology,1997,33(1):61-70.
[25] 張玉秀,王梓.脫水蛋白在逆境下的分子作用機制研究進展[J].自然科學進展,2007,17(1):1-10.
[26] Cellier F,Conejero G,Breitler J C,et al.Molecular and physiological responses to water deficit in drought tolerant and drought sensitive lines of sunflower[J].Plant Physiology,1998,116(1):319-328.
[27] Caruso A,Morabito D,Delmotte F,et al.Dehydrin induction during drought and osmotic stress in populus[J].Plant Physiology and Biochemistry,2002,40(12):1033-1042.
[28] 張茹,李金花,柴兆祥,等.Chi基因的克隆及轉基因馬鈴薯植株的獲得[J].草業學報,2009,18(6):51-58.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
