RGMa在創傷后精神應激障礙大鼠內側前額皮質的表達變化
2012-12-03 08:09:06溫莉莉韓芳石玉秀
中國醫科大學學報 2012年12期
溫莉莉,韓芳,石玉秀
(中國醫科大學基礎醫學院組織學與胚胎學教研室,沈陽110001)
創傷后精神應激障礙(post-traumatic stress disorder,PTSD)是由異常精神心理創傷所致的以再度體驗創傷并伴有易激惹情緒和回避行為的臨床表現為特征的長期持續性精神障礙。研究表明PTSD患者腦部的海馬,杏仁核都有減小,其內有神經細胞的凋亡[1,2],而排斥性導向分子 A(repulsive guidance molecule A,RGMa)是一種新的軸突生長抑制蛋白,在中樞神經生長發育過程中起介導排斥性軸突導向信號和神經管閉合、調控神經細胞的增殖、分化、存活、誘導生長錐塌陷而阻斷受損神經再生等作用[3]。大鼠前額皮質(medial prefrontal cortex,mPFC),是認知、記憶和情感的重要腦區[4~5]。mPFC在情緒反應中起重要作用[6],文獻報道PTSD腦萎縮可能由神經元細胞凋亡所致[7],從RGMa在PTSD mPFC的表達變化有可能揭示PTSD大鼠mPFC神經元凋亡發生機制。RGMa的激活可破壞生長錐,使軸突回縮、生長抑制[3],研究RGMa在PTSD大鼠的表達變化可能揭示PTSD mPFC縮小的發病機制之一。本研究利用國際認定的PTSD-SPS模型,采用免疫組織化學、免疫印跡方法檢測PTSD大鼠mPFC神經元RGMa表達變化,為研究PTSD致細胞凋亡的機理提供實驗依據。
1 材料與方法
1.1 實驗動物及分組
由中國醫科大學實驗動物中心提供成年健康雄性Wistar大鼠45只,體質量180~220 g,分籠喂養,自由飲食,實驗室內通風良好,室溫保持在18~25℃,先將大鼠在實驗室條件適應性喂養1周后,將其隨機分為對照組、PTSD-SPS組(7 d和14 d組),每組15只。
1.2 PTSD-SPS大鼠模型建立與行為學驗證
采用2005年日本文部省召開的“基礎和臨床研究進展”國際會議確定的關于大鼠PTSD模型-SPS[8]。將大鼠連續進行下述步驟處理:禁錮2 h;后強迫游泳20 min(水深40 cm,水溫25℃);休息15 min后乙醚麻醉至意識喪失;無干擾常規喂養直至取材前通過行為學驗證確認符合PTSD后取材。
1.3 制備光鏡標本
對SPS 7 d、14 d及正常對照組大鼠分別進行心臟灌流固定。步驟如下述:大鼠經2%戊巴比妥腹腔注射麻醉后,暴露心臟,左心室插管,剪開右心耳,先以生理鹽水300 mL快速灌沖血道,后以4%多聚甲醛進行灌注固定,取出腦組織,并浸入4%多聚甲醛續固定3 h,然后浸入Holt′s液(用0.01 mol/L PBS配制的40%蔗糖)中至沉瓶底。于恒冷箱切片機(萊卡)行冠狀切片,片厚8 μm,用于免疫組化染色。
1.4 RGMa免疫組織化學SABC法染色
冰凍切片經吹干,依次用3%H2O2-dH2O漂洗10 min,PBS(含0.03%Triton X-100) 漂洗10 min×3次,5%BSA封閉20 min后,滴加兔多克隆抗體RGMa(工作濃度為1∶200,Santa Cruz Bio-technology)0.01 mol/L,PBS代替一抗作陰性對照,4 ℃孵育過夜,滴加生物素化羊抗兔IgG37℃,30 min,滴加SABC37℃,20 min,以上各步間均用0.01 mol/L PBS漂洗,之后DAB(武漢博士德公司)顯色,常規脫水,透明,中性樹膠封片,以PBS液代替一抗作陰性對照,光學顯微鏡 (OLYMPUS,BX60,Japan) 觀察mPFC的RGMa表達,攝片。同倍率下,觀察5張連續切片,每張切片隨機選取3個視野進行陽性細胞計數。用形態學圖像分析系統軟件,記錄陽性細胞吸光度(absorbance,A),對以上免疫組化顯色切片進行半定量分析。
1.5 免疫印跡(Western blot)
脫頸法處死大鼠,冰上快速取出正常對照組及SPS 7 d、14 d組大鼠mPFC,組織經勻漿、超聲粉碎后高速低溫離心,12000 r/min,取上清;采用考馬斯亮藍法測定蛋白濃度;每例標本提取蛋白50 μg,10%SDS PAGE變性凝膠電泳,濃縮膠90 V、30 min和分離膠110 V、120 min;恒壓轉移到PVDF膜上,4℃過夜,5%脫脂奶粉封閉90 min;加入兔多克隆多抗體RGMa(工作濃度為1∶200)中孵育,4℃過夜;IgG抗體-HRP多聚體孵育2 h;ECL發光,以目的條帶與內參照GAPDH的平均光密度(OD)的比值表示相對表達水平,進行半定量分析。
2 結果
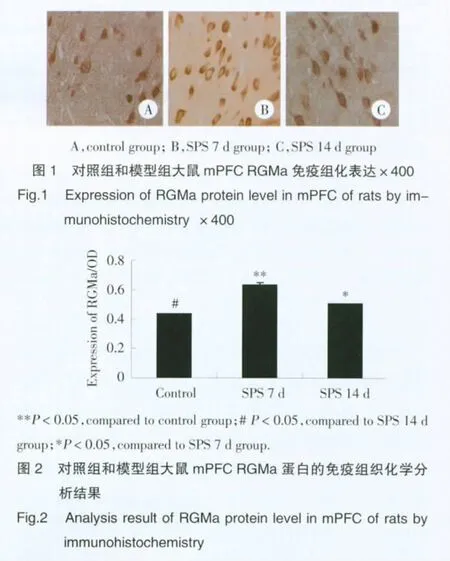
2.1 RGMa免疫組織化學染色
顯色結果表明,RGMa在正常對照組和模型組均有表達,且陽性信號表達在神經元和突起內,見圖1。分析結果顯示,SPS 7 d組和14 d組高于對照組,差異有統計學意義(P<0.05);3組陽性細胞OD值分別是(0.44±0.002)、(0.63±0.015)和(0.51±0.009),SPS 7 d組RGMa的表達增多,SPS14 d組RGMa表達稍有降低,但較對照組仍明顯增高,差異有統計學意義(P<0.05),見圖2。
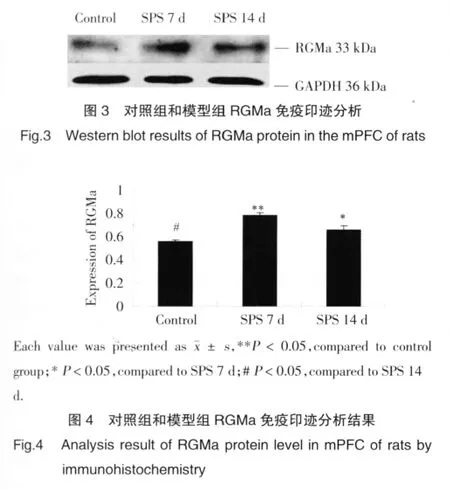
2.2 SPS大鼠mPFC組織蛋白提取及免疫印跡法測定RGMa
免疫印跡測試顯示,在33 kDa處有陽性條帶。分析結果表明,對照組的mPFC RGMa有一定表達,SPS 7 d組表達量明顯增多,差異具有統計學意義(P<0.05),SPS14 d組表達較7 d組減少,較正常組增多(圖3,圖 4)。
3 討論
現代社會壓力日趨增大,巨大自然災害(海嘯、地震、洪災等)近年頻發,故此PTSD受到國內外極大關注[9]。PTSD-SPS大鼠模型是2005年日本文部省召開的“基礎和臨床研究進展”國際會議確定的關于大鼠PTSD模型[8],該模型以環境條件刺激(強迫性游泳)作為應激刺激,SPS大鼠能表現出PTSD相似的臨床行為學改變[10],更適于PTSD的研究。目前國外研究發現PTSD患者MRI檢查見海馬體積縮小、萎縮[11,12]。所以推測記憶的異常可能與海馬體積減小有關。杏仁核活躍,PTSD大鼠杏仁核神經細胞樹突增多,推測恐怖增強可能與此有關[2,13]。國內外的學者對PTSD的研究主要集中在行為學和內分泌的改變,對細胞通路的研究較少。RGM是分子量為33/35 kDa的排斥導向分子,其家族成員有RGMa、RGMb RGMc,其中 RGMa、在成年大鼠神經元有表達,在腦損傷時表達增強[11]。RGMa的高親和力受體neogenin主要表達于發育中和成熟的中樞神經系統[14],而RGMa可在創傷條件下與其高親和力受體結合,通過激活RhoA-ROCK(RhoA-Rhoassociated kinase)而使神經元生長錐崩塌,抑制大腦神經元軸突生長[11,12,15]。因此,RGMa 參與神經元的再生,mPFC在情感反應中起到核心作用。研究RGMa在mPFC的表達對揭示PTSD凋亡機制有一定意義。
本實驗觀察到RGMa在PTSD大鼠mPFC的表達增加,RGMa在SPS后表達增高尤其是SPS 7 d表達量最多,提示 RGMa參與PTSD的應激過程。RGMa是一種新的軸突生長抑制蛋白,可誘導生長錐塌陷進而阻斷受損神經再生等作用[3],致中樞神經系統受到損傷后神經元的再生困難,在哺乳動物腦和脊髓損傷后其神經元中誘導抑制性或排斥性導向分子表達增多[14]。研究報導阻斷RGMa蛋白后能使neogenin與RGMa分離,促使neogenin與netrin-1結合,促進軸突生長[16]。我們推測PTSD后情感改變、恐懼增強,與創傷后RGMa表達增多有關,RGMa表達增多可能使神經元生長錐塌陷抑制神經元再生,使PTSD神經元萎縮、凋亡。RGMa在創傷條件下與其高親和力受體結合可通過激活RhoAROCK(RhoA-Rho-associated kinase)而使神經元生長錐崩塌,抑制腦神經元軸突生長[15],進而神經細胞萎縮甚至凋亡。
綜上所述,本實驗結果顯示RGMa在mPFC的細胞凋亡時有很強的過表達,我們推測RGMa/neogenin信號有可能是PTSD細胞凋亡的重要調控信號。SPS大鼠mPFC中RGMa表達增高,可能通過RhoA-ROCK(RhoA-Rho-associated kinase)通路抑制神經元再生和修復,加重PTSD大鼠mPFC神經元凋亡的發生。本研究為闡明PTSD發病機制提供了部分實驗依據。
[1]Hull AM.Neuroimaging findings in post-traumatic stress disorder systematic review[J].Br J Psychiatry,2002,181:102-110.
[2]Xiao B,Han F,Shi Yu-xiu,et al.Single-prolonged stress induces apoptosis by activating cytochrome c/caspase-9 pathway in a rat model of posttraumatic stress disorder[J].Cell Mol Neurobiol,2011,31(1):37-43.
[3]Hata K,Fujitani M,Yasuda Y,et al.RGMa inhibition promotes axonal growth and recovery after spinal cord injury [J].J Cell Biol,2006,173(1):47-58.
[4]Mottaghy FM,Gangitano M,Sparing R,et al.Segregation of areas related to visual working memory in the prefrontal cortex revealed by rTMS[J].Cereb Cortex,2002,12(4):369-375.
[5]Meeldinger A,Gruenewald C,Beseon M,et al.Separable neuronal circuitries for manipulable and non-manipulable objects in working memory[J].Cereb Cortex,2002,12(11):lll5-1123.
[6]Bechara A,Damasio H,Damasio AR.Emotion,decision making and the orbitofrontal cortex[J].Cerebral Cortex,2000,10(3):295-307.
[7]Li XM,Han F,Liu DJ,et al.Single-prolonged stress iduced mitochondrial-dependent apoptosis in hippocampus in the rat model of post-traumatic stress disorder[J].J Chem Neuroanat,2010,40(3):248-255.
[8]Liberzon I,Krstov M,Young EA.Stress restress:effects on ACTH and fast feedback[J].Psychoneuroendocrinology,1997,22(6):443-453.
[9]Li S,Yukihisa M,Wang M,et al.The effects of chronic valproate and diazepam in a mouse model of posttraumatic stress disorder[J].Pharmacol Biochem Behav,2006,85(2):324-331.
[10]lmanaka A,Mofinobu S,Toki S,et a1.Importance of early environment in the development of post-traumatic stress disorder-like behaviors[J].Behav Brain Res,2006,173(1):129-137.
[11]Apfel BA,Ross J,Neylan TC,et al.Hippocampal volume differences in gulf war veterans with current versus lifetime posttraumatic stress disorder symptoms[J].Biol Psychiatry,2011,69(6):541-548.
[12]Wang Z,Neylan TC,Muellea SG,et al.Magnetic resonance imaging of hippocampal subfields in posttraumatic stress disorder[J].Arch Gen Psychiatry,2010,67(3):296-303.
[13]Liu H,Wang HT,Han F,et al.Activity of5-HT1A receptor is involved in neuronal apoptosis of the amygdala in a rat model of posttraumatic stress disorder[J].Mol Med Rep,2011,4(2):291-295.
[14]Mueller BK,Yamashita T,Schaffar G,et al.The role of repulisive guidance molecules in the embryonic and adult vertebrate central nervoussystem[J].PhiloTransRSocLondBBioSci,2006,361(1473):1513-1529.
[15]Bae GU,Yang YJ,Krauss RS.Neogenin regulates skeletal myofiber size and focal adhesion kinase and extracellular signal-regulated kinase activities in vivo and in vitro[J].Mol Biol Cell,2009,20(23):4920-4931.
[16]Manitt C,Thompson KM,Kennedy TE,et al.Developmental shift in expression of netrin receptors in the rat spinal cord:predominance ofUNC-5homologuesinadulthood[J].Neurosci Res,2004,77(5):690-700.
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
光學精密工程(2016年6期)2016-11-07 09:07:19
