一株納豆芽孢桿菌的產酶條件優化
2013-02-13 08:15:34吳慶紅陳義倫
食品科學 2013年3期
關鍵詞:優化
李 妍,吳慶紅,陳義倫,*,周 波*
(1.山東農業大學食品科學與工程學院,山東 泰安 271018;2.泰山學院附屬中學,山東 泰安 271018;3.土肥資源高效利用國家工程實驗室,山東農業大學生命科學學院,山東 泰安 271018)
血栓病威脅著人類的健康與生命,溶解血栓是治療這類疾病的首選方法;當前的溶栓劑,如尿激酶、鏈激酶、重組組織型纖溶酶原激活劑等,存在半衰期短、副作用大、價格昂貴等缺點[1-3]。納豆激酶(nattokinase,NK)是一種在納豆發酵過程中由納豆菌(Baciuus natto)或納豆枯草桿菌(B.subtilis natto)產生的絲氨酸蛋白酶[4-5],研究表明[6-8]納豆激酶成本低廉,作用迅速,藥效時間長,安全性好,具有很強的纖溶活性,不但能直接作用于纖溶蛋白,而且還能激活體內纖溶酶原,從而增加內源性纖溶酶的量與作用,有望稱為新型的溶栓藥物。響應面分析法(response surface methodology,RSM)是能通過綜合實驗設計和數學建模得到準確有效結論的實驗設計方法[9-10],被廣泛應用于各類發酵條件的優化[11-15]。
本課題組在前期的研究中篩選到一株納豆芽孢桿菌,采用Plackett-Burman法和響應面分析法的方法對該菌產納豆激酶的液體發酵條件進行優化,以期獲得較高的納豆激酶產量,對該酶進一步的產業化應用提供理論依據。
1 材料與方法
1.1 材料與培養基
1.1.1 材料與試劑
菌種為山東農業大學資源與環境微生物實驗室保藏的納豆芽孢桿菌。
凍干人纖維蛋白原 上海萊士血制品有限公司;凝血酶 美國Sigma公司;尿激酶 阿拉丁公司。
1.1.2 培養基
斜面培養基(g/L):胰蛋白胨10、酵母抽提物5、NaCl 10,pH7.0;種子培養基:純牛乳100mL、葡萄糖2.5g;發酵培養基:胰蛋白胨、乳糖、Na2HPO4、NaH2PO4、CaCl2、MgSO4,pH7.0。
1.2 方法
1.2.1 粗酶液的制備
納豆芽孢桿菌菌株經過斜面培養基活化24h后,轉接到種子培養基中,30℃、180r/min培養12h,按一定的接種量接到發酵培養基中,30℃、180r/min培養56h后,取發酵液于4℃、8000r/min冷凍離心10min得到的上清液,即為粗酶液。
1.2.2 酶活力的測定方法
納豆激酶酶活力測定采用纖維蛋白平板法[10],以尿激酶繪制標準曲線。
1.2.3 產酶條件的單因素試驗
在發酵培養基的基礎上選擇不同的碳源、氮源以及營養鹽之比進行發酵產酶[16-17],確定菌株的產酶最優培養基組成;并且在產酶最優培養基的基礎上進行發酵條件的優化,包括種齡、接種量、發酵時間、培養溫度、裝液量。
1.2.4 Plackett-Burman篩選影響產酶的重要因素設計
實驗中選用n=12的PB設計,把每個因素設計成高(+1)和低(-1)2個水平。
1.2.5 響應面優化試驗
采用響應面分析法中的Box-Behnken試驗設計[18-20],對PB實驗篩選到的關鍵因子做進一步的分析,獲得影響該菌發酵產酶的最優發酵條件。
1.3 數據分析
采用Design Expert統計軟件分析數據。
2 結果與分析
2.1 納豆芽孢桿菌產酶條件的優化結果
2.1.1 最適發酵培養基的優化結果
2.1.1.1 碳源對產酶的影響
采用的基礎培養基(g/L)為:胰蛋白胨20.0、Na2HPO45.0、NaH2PO41.0、CaCl20.2、MgSO40.5,分別以葡萄糖、乳糖、麥芽糖、蔗糖、麩皮為碳源,添加量為10g/L,進行發酵產酶實驗,研究碳源對產酶的影響,結果見圖1。乳糖作為碳源時,最有利于產酶,其次是葡萄糖;麩皮和麥芽糖作為碳源,產酶量不高,當蔗糖作為碳源時,發酵過程則不產生納豆激酶,這與已往的研究存在差異[16],可能是因為菌株的差異造成的結果。

圖1 碳源對產酶的影響Fig.1 Effect of carbon source on the activity of nattokinase
2.1.1.2 氮源對產酶的影響
采用的基礎培養基(g/L)為:乳糖10.0、Na2HPO45.0、NaH2PO41.0、CaCl20.2、MgSO40.5,分別以胰蛋白胨、酵母粉、牛肉膏、硫酸銨、磷酸二氫銨作為氮源,添加量為20g/L,進行發酵產酶實驗,研究氮源對產酶的影響,結果見圖2。最適氮源為牛肉膏,其次是胰蛋白胨和酵母粉,而硫酸銨和磷酸二氫銨作為氮源不能滿足大量產酶的需求,出于實驗操作的考慮,本實驗采用胰蛋白胨作為氮源。

圖2 氮源對產酶的影響Fig.2 Effect of nitrogen source on the activity of nattokinase
2.1.2 最適發酵條件的優化結果
2.1.2.1 種齡對產酶的影響
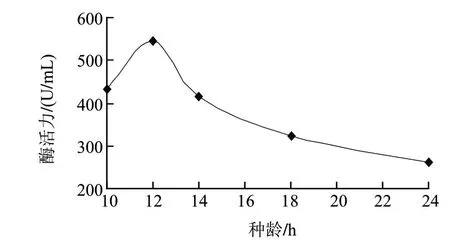
圖3 種齡對產酶的影響Fig.3 Effect of inoculation age on the activity of nattokinase
在最適發酵培養基的基礎上,將種齡分別為10、12、14、18、24h的種子活化液接入搖瓶進行發酵實驗,結果見圖3。最初隨著種齡的增加,細菌數量的增多,產酶量逐漸提高,當用種齡為12h的活化液進行發酵,最有利于產酶。當種齡超過12h后,隨著細菌生長活力的下降,產酶量開始降低,至24h時活化液非常不利于產酶。
2.1.2.2 接種量對產酶的影響
在最適發酵培養基的基礎上,將種齡為12h的種子活化液按1%、2%、3%、4%、5%、6%的接種量接入搖瓶進行發酵實驗,結果見圖4。接種量為2%時,最有利于產酶。接種量過高或過低都不利于產酶,過低影響細菌數量,導致產酶不足;過高則因為培養基的營養成分被大量的微生物的生長而快速消耗,影響了發酵后期的產酶。
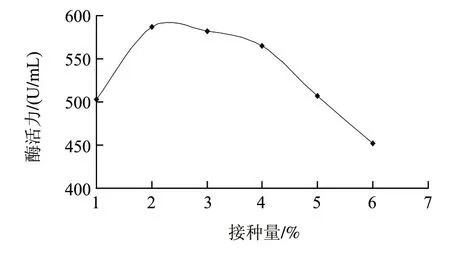
圖4 接種量對產酶的影響Fig.4 Effect of inoculum size on the activity of nattokinase
2.1.2.3 發酵時間對產酶的影響
在最適發酵培養基的基礎上,選定不同發酵時間12、24、36、48、56、64、72h進行酶活力測定,結果見圖5。隨著發酵時間的增加,產酶量逐漸提高,當發酵時間超過50h,產酶量趨于穩定,因此將56h作為最適發酵時間。

圖5 發酵時間對產酶的影響Fig.5 Effect of fermentation time on the activity of nattokinase
2.2 Plackett-Burman設計篩選影響產酶的重要因素
在單因素試驗的基礎上選取各因素的高、低水平,按表1設計進行了3輪重復試驗,取樣測定納豆激酶的酶活力,取3次測量的平均值,結果見表1。計算各因素效應,評價其重要性,各因素所代表的參數、水平見表2。這個模型是顯著的,胰蛋白胨添加量、MgSO4添加量、接種量及發酵溫度是影響產酶的重要因素,其中胰蛋白胨添加量、MgSO4添加量、接種量呈現顯著的正效應,發酵溫度則呈現負效應,因此應當提高胰蛋白胨添加量、MgSO4添加量、接種量,降低發酵溫度。根據P值大小,選擇3個因素:胰蛋白胨添加量、MgSO4添加量及發酵溫度進行響應面優化試驗。
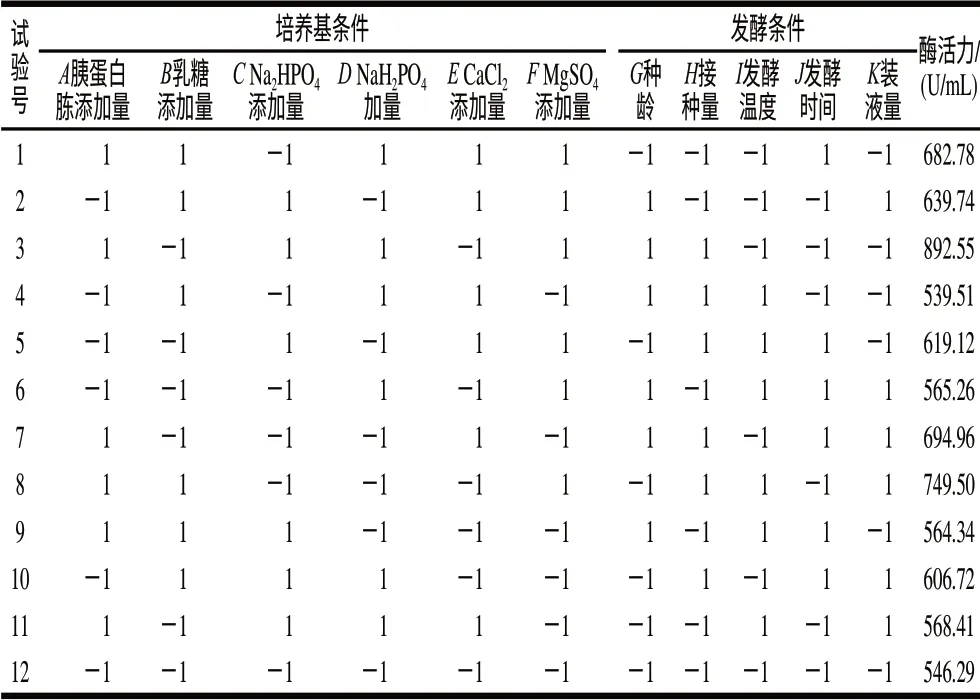
表1 Plackett-Burman試驗設計與結果(n=12)Table 1 Design and results of Plackett-Burman tests (n=12)
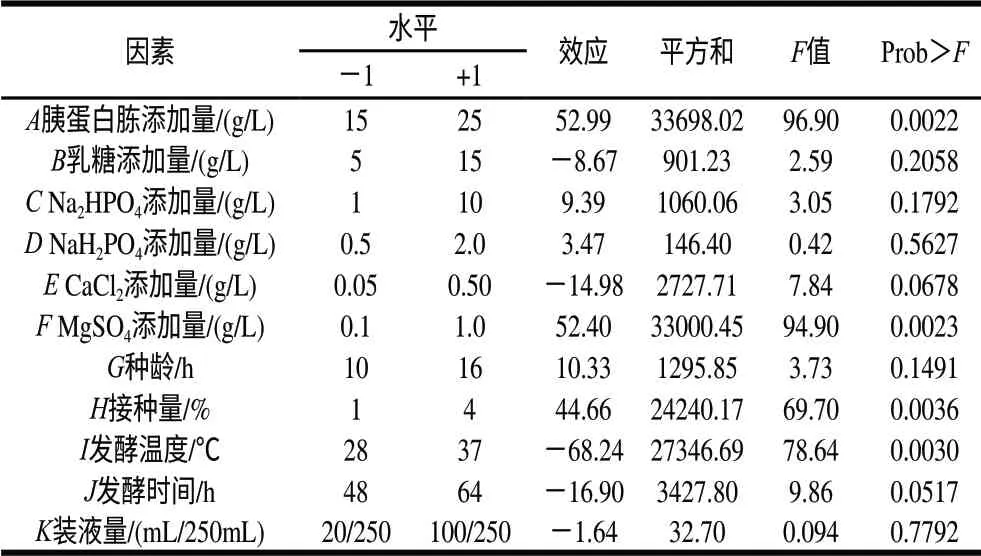
表2 Plackett-Burman試驗因素、水平及其效應Table 2 Factors, levels and effects of Plackett-Burman tests
2.3 中心組合試驗
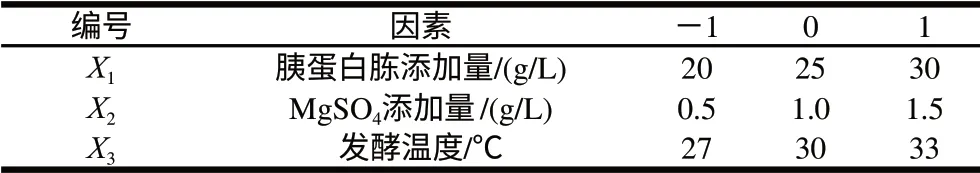
表3 中心組合試驗的自變量及其水平Table 3 Variables and levels of central composite tests
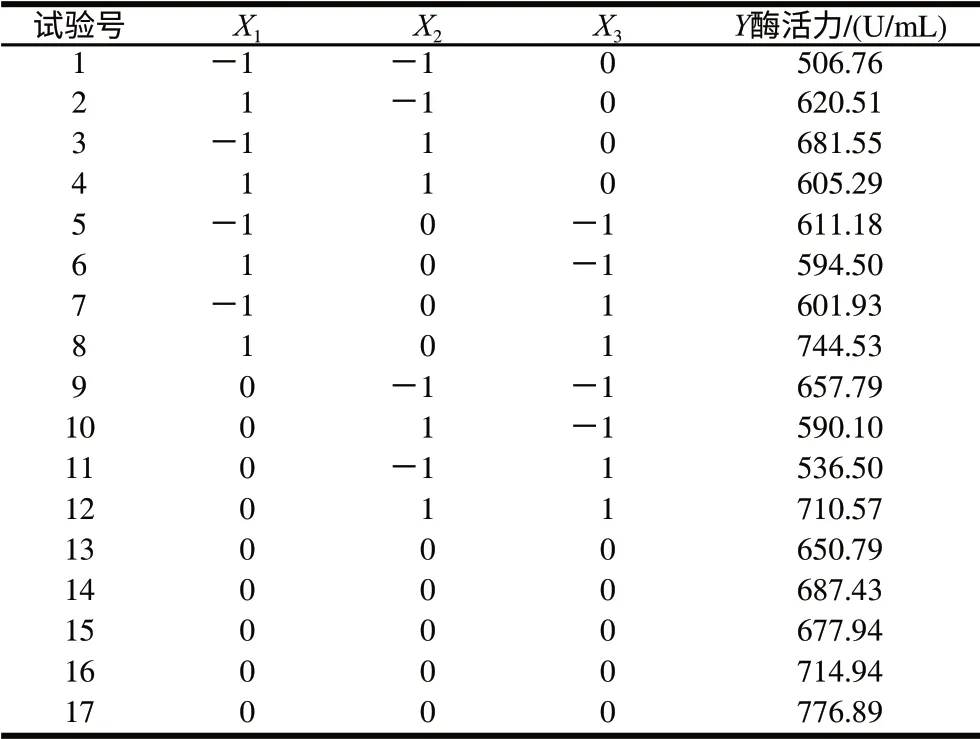
表4 中心組合試驗設計及結果Table 4 Design and results of central composite tests
利用Box-Behnken進行響應面試驗設計,對PB試驗篩選出3個顯著因素進行優化,以酶活力為響應值,對應于因變量Y。各自變量水平見表3,試驗設計及結果見表4。
通過Design Expert1軟件對試驗結果進行分析,獲得回歸方程如下:
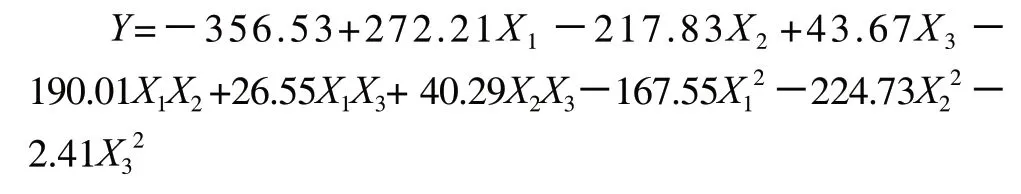
通過對上述二次回歸方程進行方差分析,驗證回歸模型及各參數的顯著度,結果見表5。

表5 酶活力二次多項模型的方差分析表Table 5 Analysis of variance for quadratic model of nattokinase activity
由表5可見,模型Prob>F值<0.0500,該模型是顯著的,確定系數R2=96.3%,表明該模型與實際情況擬合很好。因此該模型可用于分析和預測納豆芽孢桿菌液體發酵產納豆激酶的酶活力。
圖6~8為響應面分析的曲面圖,可直觀地看出各變量與響應值,變量與變量之間的關系,而且回歸模型確實存在最大響應值。該模型可預測的最大值為739.818U/mL,此時極值點坐標:X1=26.6g/L,X2=1.35g/L,X3=33℃。為了檢驗模型預測的準確性,在優化條件下進行發酵驗證實驗,即發酵培養基條件為:胰蛋白胨26.6g/L、乳糖10.0g/L、Na2HPO45.0g/L、NaH2PO41.0g/L、CaCl20.2g/L、MgSO41.35g/L;發酵條件為:種齡12h、接種量2%、發酵溫度33℃、發酵時間56h、裝液量50mL/250mL時,所得納豆激酶酶活力為743.65U/mL,比優化前提高了232.33U/mL,可見該模型能較好的預測實際產酶情況。
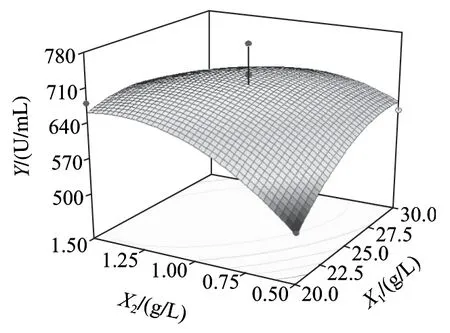
圖6 Y=f(X1, X2)響應面圖Fig.6 Response surface plot for Y = f(X1, X2)
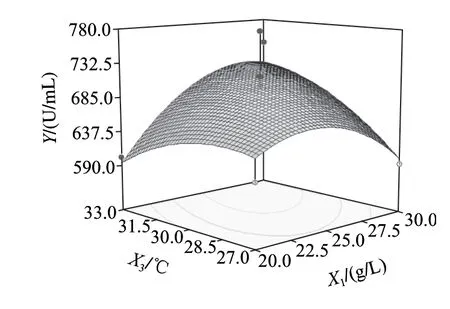
圖7 Y=f(X1, X3)響應面圖Fig.7 Response surface plot for Y = f(X1, X3)
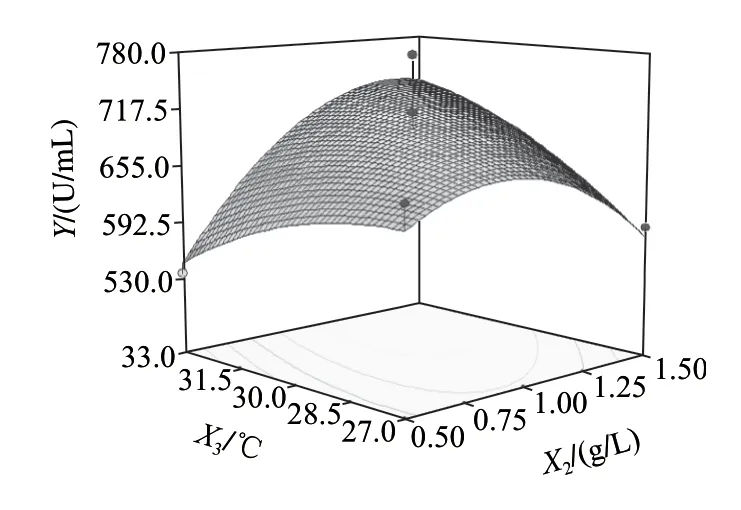
圖8 Y=f(X2, X3)響應面圖Fig.8 Response surface plot for Y = f(X2, X3)
3 結 論
本研究在單因素試驗的基礎上,利用Plackett-Burman法篩選出影響產納豆激酶的重要因素有胰蛋白胨添加量、MgSO4添加量和發酵溫度;然后采用Box-Behnken實驗設計對這3個重要因素做進一步分析,獲得最優的發酵產酶條件為:發酵培養基(g/L)條件:胰蛋白胨26.6、乳糖10.0、Na2HPO45.0、NaH2PO41.0、CaCl20.2、MgSO41.35;發酵條件:種齡12h、接種量2%、發酵溫度33℃、發酵時間56h、裝液量50mL/250mL;此時實際產酶活力為743.65U/mL,與回歸模型預測值存在較小的誤差,驗證了回歸模型的準確性;在此發酵條件下,納豆激酶的酶活力較優化前提高了232.33U/mL。
[1]SHERRY S.Recombinant tissue plasminogen activator(rt-PA)∶ is it the thrombolytic agent of choice for an evolving acute myocardial infarction[J].Am J Cardiol, 1987, 59(9)∶ 984-989.
[2]FUJITA M, HONG K, ITO Y, et al.Thrombolytic effect of nattokinase on a chemically induced thrombosis model in rat[J].Biol Pharm Bull,1995, 18(10)∶ 1387-1391.
[3]SUMI H, HAMADA H, NAKANISHI K.et al.Enhancement of the fibrinolytic activity in plasma by oral administration of nattokinase[J].Acta Haematol, 1990, 84∶ 139-143.
[4]SUMI H, HAMADA H, TSUSHIMA H, et a1.A novel fibrinolytic enzyme(nattokinase)in the vegetable cheeseNatto∶ a typical andpopular soybean food in the Japanese diet[J].Experientia, 1987,43(10)∶ 1110-1111.
[5]陳志文, 徐爾尼, 肖美燕.納豆激酶的研究進展[J].食品科學, 2002,23(10)∶ 130-134.
[6]杜平中.溶栓藥物的研究進展[J].國外醫藥∶ 合成藥、生化藥、制劑分冊, 1998, 19(2)∶ 67-71.
[7]鄒和昌.溶栓劑的發展及研究[J].中國藥學雜志, 1997, 32(5)∶ 263.
[8]遲東升, 阮新民.新型溶栓劑∶ 納豆激酶[J].心血管病學進展, 2007,28(4)∶ 545-550.
[9]慕運動.響應面方法及其在食品工業中的應用[J].鄭州工程學院學報, 2001, 22(3)∶ 91-94.
[10]ASTRUP T, MULLERTZ S.The fibrin plate method for estimating fibrinolytic activity[J].Arch Biochem Biophys, 1952, 40(2)∶ 346-351.
[11]盧可, 婁永江, 周湘池.響應面優化楊梅果醋發酵工藝參數研究[J].中國調味品, 2011, 36(2)∶ 57-60.
[12]董超, 李楠, 程輝彩, 等.納豆激酶發酵條件的優化[J].食品科學,2009, 30(1)∶ 151-154.
[13]王樂.納豆激酶高產菌株篩選及發酵工藝優化研究[M].濟南∶ 山東輕工業學院, 2009.
[14]王萍, 杜連祥, 路福平, 等.溶栓納豆芽抱桿菌的篩選鑒定及產納豆激酶條件的研究[J].食品與發酵工業, 2006, 32(2)∶ 74-77.
[15]梅樂和, 胡升, 許靜, 等.納豆中納豆枯草桿菌的篩選和納豆激酶的分離過程研究[J].浙江大學學報∶ 工學版, 2004, 38(10)∶ 1355-1360.
[16]王成濤, 籍保平, 張麗萍.納豆激酶高活性菌株的篩選及其發酵條件的優化[J].食品科學, 2004, 25(4)∶ 71-74.
[17]桂春燕, 陳義倫, 謝曉平, 等.黃河三角洲耐高滲堿性纖維素酶產生細菌YRD-19的誘變育種和產酶條件優化[J].食品工業科技, 2010,31(10)∶ 163-168.
[18]STOWE R A, MAYER R P.The application of PB method in bacteria research[J].Industrial and Engineering Chemistry, 1996, 58(2)∶ 36-40.
[19]胡升, 梅樂和, 姚善涇.響應面法優化納豆激酶液體發酵[J].食品與發酵工業, 2003, 29(1)∶ 13-17.
[20]張健, 高年發.利用響應面法優化丙酮酸發酵培養基[J].食品與發酵工業, 2006, 32(8)∶ 52-55.
猜你喜歡
房地產導刊(2022年5期)2022-06-01 06:20:14
能源工程(2022年1期)2022-03-29 01:06:28
建材發展導向(2021年12期)2021-07-22 08:06:48
建材發展導向(2021年7期)2021-07-16 07:07:52
中學生數理化(高中版.高二數學)(2021年12期)2021-04-26 07:43:48
中學生數理化(高中版.高考數學)(2021年12期)2021-03-08 01:28:50
今日農業(2020年16期)2020-12-14 15:04:59
消費導刊(2018年8期)2018-05-25 13:20:08
家庭影院技術(2018年4期)2018-05-09 07:07:41
電子制作(2017年20期)2017-04-26 06:57:45
