水稻ERECTA基因組DNA的克隆及植物表達載體構建
2013-04-29 12:17:00韓同凱王盈盈林紅珍等
山東農業科學 2013年6期
關鍵詞:水稻
韓同凱 王盈盈 林紅珍等
作者簡介:韓同凱(1987-),男,碩士研究生,研究方向為作物遺傳育種。
通訊作者:陳翠霞(1964-),女,教授,研究方向為作物遺傳育種。
謝先芝(1972-),女,副研究員,研究方向為植物發育生物學。
摘要:根據水稻ERECTA的cDNA序列(GenBank中的登陸號:Os06g0203800),利用NCBI的Blast工具找到OsERECTA所對應的基因組DNA序列(gERECTA),根據該序列設計2對特異引物,對gERECTA進行分段PCR擴增,并對前后兩部分片段進行克隆、測序和拼接。本研究得到的gERECTA序列為8 901 bp,其中包括約17 kb的 5′區域和約300 bp 的3′區域。將拼接起來的gERECTA序列插入到pCAMBIA1390載體,經酶切和測序鑒定,證明重組表達質粒中帶有gERECTA基因片段,表明gERECTA植物表達載體pCAMBIA1390-gERECTA已成功構建。
關鍵詞:水稻;OsERECTA基因;克隆;植物表達載體
中圖分類號:Q78 文獻標識號:A 文章編號:1001-4942(2013)06-0004-07
水稻ERECTA基因是植物中編碼富含亮氨酸重復序列的類受體蛋白激酶基因家族(Leucine-rich repeat receptor-like kinase gene family)中的一員。Torii等(1996)[1]首次克隆得到擬南芥ERECTA基因。擬南芥ERECTA基因在頂端分生組織、植株幼嫩的或正在生長的地上部分包括生長發育中的葉片和花器官中表達,在成熟器官和根中不表達,且在植株發育早期的頂端分生組織中表達較弱,但在營養生長向生殖生長過渡階段表達水平提高[2]。
隨著對模式植物擬南芥ERECTA基因研究的深入,發現ERECTA基因通過協調細胞分裂和細胞增殖調節包括花在內的氣生器官結構(如節間和花梗的伸長和近軸-遠軸極性)和氣孔模式(如氣孔密度和分布)[3~6]。已有報道證明擬南芥ERECTA基因調控某些生理過程如二磷酸核酮糖羧化酶(Rubisco)的羧化率、電子傳遞能力、植株的蒸騰作用和水分利用效率等[6]。且ERECTA基因在植株對生物和非生物環境如熱脅迫、干旱和細菌感染的響應中起作用[6~8]。
尹偉倫課題組克隆得到Populus nigra×(Populus deltoids×Populus nigra)ERECTA(PdERECTA)基因并培育了轉基因擬南芥植株(35S∶ PdERECTA)。PdERECTA過表達導致苗期初生根變長、葉面積變大尤其是長期水分利用率(Long-term water use efficiency)大大增強,這揭示出ERECTA基因具有提高作物水分利用率的潛在作用[9]。朱錦程等利用RT-PCR技術從擬南芥中克隆得到ERECTA基因并構建真核表達載體pBin438-ERECTA,轉化番茄,結果證實轉基因番茄植株的水分利用率得到提高[10]。
大多數高等真核生物基因都含有內含子,其轉錄后的序列在剪接過程中從mRNA前體上切除。但很多情況下,內含子對基因的功能是非常重要的,可能同選擇性剪接有關或者內含子本身包含了重要的非編碼RNAs和ORFs。內含子具有增強基因表達的作用,例如,在過渡相實驗(Transient assay)中,包含內含子的玉米乙醇脫氫酶1(Alcohol dehydrogenase-1)的表達量提高了50~100倍[11]。Karve等發現擬南芥ERECTA的表達強烈依賴內含子的存在,內含子是ERECTA基因mRNA積累所必需的[12]。因此,我們推測OsERECTA基因的表達同樣需要內含子存在,本研究構建了包含內含子的ERECTA基因組DNA的植物表達載體。
1 材料與方法
11 試驗材料
111 植物材料 水稻品種日本晴(Oryza sativa L cv Nipponbare)。樣品為溫室中生長2周的幼苗葉片,液氮速凍后置于-70℃保存,用于全基因組DNA提取。
112 試驗試劑 限制性內切酶、DNA Ligase、DNA片段分子量標記(λ-EcoT 14 I digest)、BAP均購于TaKaRa公司;質粒小提試劑盒購于Bioteke公司;KOD-Plus-Neo DNA聚合酶購于ToYoBo公司;DNA膠回收試劑盒、卡那霉素(Kan)、氨芐青霉素(Amp)購于上海生工生物工程有限公司。
113 載體及菌株 克隆載體pMD18-T Vector、pTA2-T vector購于TaKaRa公司;表達載體pCAMBIA1390(CAMBIA, Canberra, Australia)由本實驗室保存;E coli DH 5α由本實驗室保存。pMD18-T Vector、pTA2-T vector具有Amp抗性;pCAMBIA1390具有Kan抗性。
114 引物設計 利用Primer Premier 50軟件,選擇得分較高、GC含量適中、無引物二聚體和發夾結構的引物對用于擴增水稻gERECTA基因序列;引物合成和基因測序由Invitrogen公司完成。
12 實驗方法
121 PCR擴增 以全基因組DNA為模板進行PCR反應。使用KOD-Plus-Neo高保真酶(ToYoBo, Code: KOD-401),按照下列組成配成反應液(50 μl體系),反應液在冰上配制。PCR反應體系為:10× PCR Buffer for KOD- Plus- Neo 5 μl、2 mmol/L dNTPs 5 μl、25 mmol/L MgSO4 3 μl、20 μmol/L引物 (each) 075 μl、模板(Genomic DNA) 200 ng、KOD -Plus-Neo (1 U/μl) 1 μl,加ddH2O至總體積為50 μl。PCR反應條件如下:94℃預變性2 min;98℃變性10 s,(引物Tm值)退火30 s,68℃延伸3 min,35個循環。用10%(W/V)的瓊脂糖凝膠電泳檢測PCR擴增結果,并用DNA膠回收試劑盒回收目的片段。
122 重組克隆質粒的構建與鑒定 利用TaKaRa公司的pMD18-T Vector和pTA2-T Vector試劑盒將PCR產物與T載體連接,進行目的片段的克隆與鑒定。連接反應液包含1 μl pMD18-T Vector(或者pTA2-T Vector)、4 μl Insert DNA和5 μl Solution I。反應液混勻后于16℃連接反應45 min。將重組質粒轉化E coli DH 5α感受態細胞,涂布于含Amp(100 mg/L)的LB平板培養基,37℃培養過夜,經抗性篩選后挑取單菌落。搖菌提取質粒后進行酶切鑒定,鑒定為陽性的質粒進行測序。
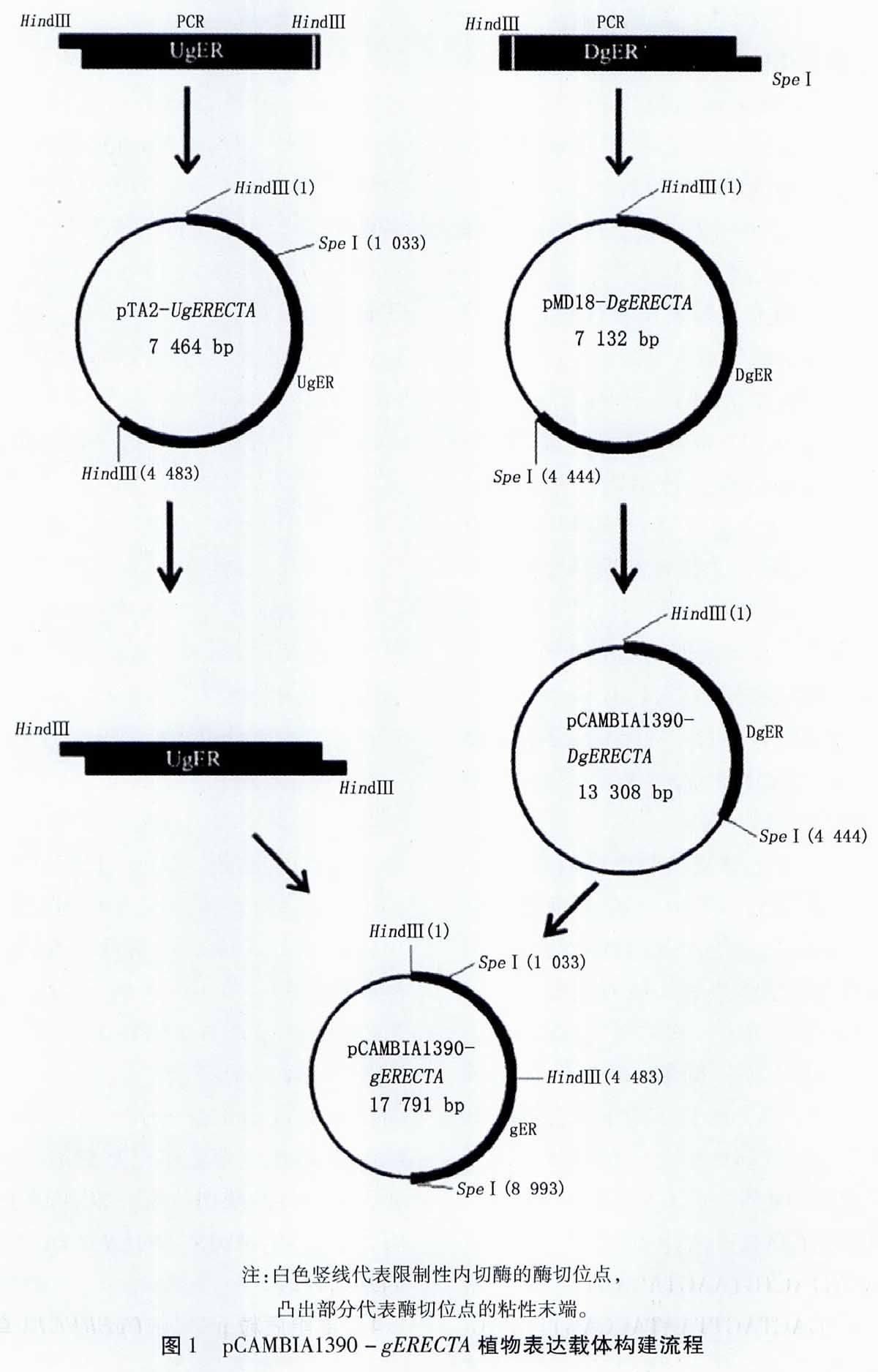
123 植物表達載體pCAMBIA1390-gERECTA的構建與鑒定 本實驗構建表達載體的流程圖見圖1。
(1)載體pCAMBIA1390-DgERECTA的構建與鑒定:Hind Ⅲ和Spe Ⅰ雙酶切陽性pMD18-DgERECTA,10%(W/V)瓊脂糖凝膠電泳分離并回收目的片段。利用DNA連接酶將目的片段定向連接到經同樣處理的植物表達載體pCAMBIA1390中,轉化E coli DH 5α感受態細胞,涂布于含Kan(50 mg/L)的LB平板培養基,37℃培養過夜,挑取單菌落。搖菌提取質粒后進行質粒雙酶切鑒定,鑒定為陽性的質粒進行測序。
(2)UgERECTA連接到pCAMBIA1390-DgERECTA載體:Hind Ⅲ酶切陽性pTA2-UgERECTA,10%瓊脂糖凝膠電泳分離并回收目的片段。利用T4連接酶將目的片段連接到經同樣酶切處理并經BAP(E coli C75)酶去磷酸化后的陽性pCAMBIA1390-DgERECTA載體中。轉化E coli DH 5α感受態細胞,涂布于含Kan(50 mg/L)的LB平板培養基,37℃培養過夜,挑取單菌落。搖菌提取質粒后進行兩次質粒單酶切鑒定,鑒定為陽性的質粒進行測序。測序正確的即為含有完整gERECTA片段的植物表達載體pCAMBIA1390-gERECTA。
2 結果與分析
21 特異性引物的設計
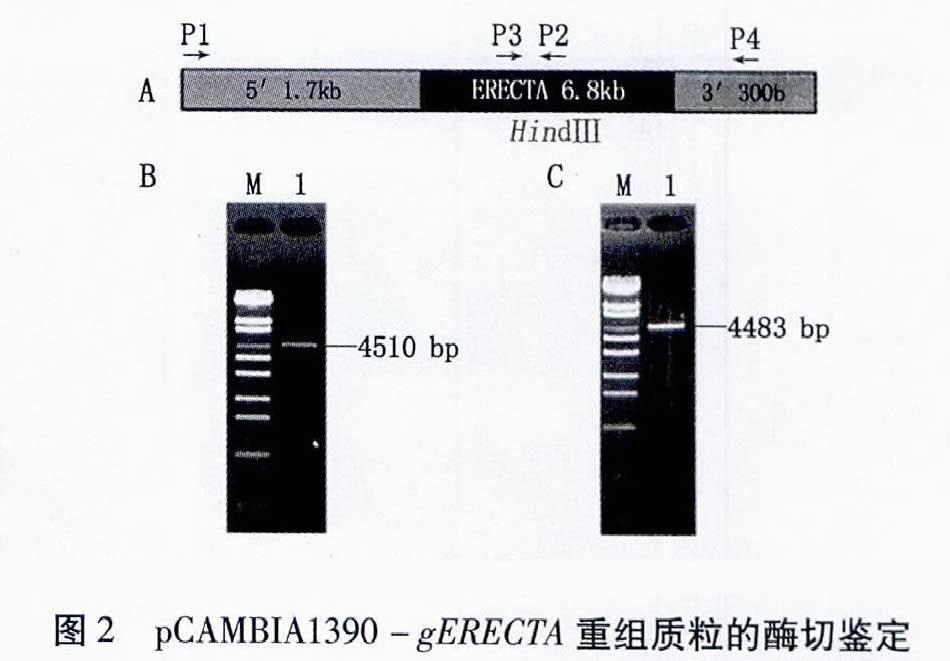
GenBank中的水稻OsERECTA基因cDNA序列所對應的基因組DNA序列(GenBank登陸號:AP0049913)全長為7 605 bp。利用Primer Premier 50軟件分別設計兩對特異引物,將gERECTA分為上游和下游兩部分(UgERECTA和DgERECTA)。為了便于后續與表達載體的連接,在部分引物中設計了酶切位點和保護堿基。擴增UgERECTA序列的上游引物P1: 5′-CCCAAGCTTCAATAAACAAAGGCTAATCTTG-3′,下游引物P2: 5′-GAATTAAAATATCTTGAAAGCTTCAT-3′,下劃線部分為Hind Ⅲ酶切位點(為gERECTA序列內固有的限制性酶切位點)。擴增DgERECTA序列的上游引物P3: 5′-AGGCTCTCGCTGTACTGTAAGTACAAT-3′,下游引物P4: 5′-AGACTAGTTAATACCAGTTCCAGCATTCCAATCCCAG-3′,下劃線部分為SpeⅠ酶切位點。兩對gERECTA基因特異引物設計示意圖見圖2A。
22 水稻DgERECTA和UgERECTA片段的分離
利用兩對特異性引物分別擴增UgERECTA和DgERECTA兩片段。根據gERECTA基因序列及兩對引物的設計位點,預測DgERECTA片段大小為4 510 bp,含有約300 bp的3′區域,UgERECTA片段大小為4 483 bp,含有約17 kb的5′區域。PCR擴增產物經10%(W/V)瓊脂糖凝膠電泳分離后,都分別得到一條比4 254 bp條帶稍長的片段(圖2B、C),與預期結果一致。
23 重組質粒pMD18-DgERECTA雙酶切鑒定
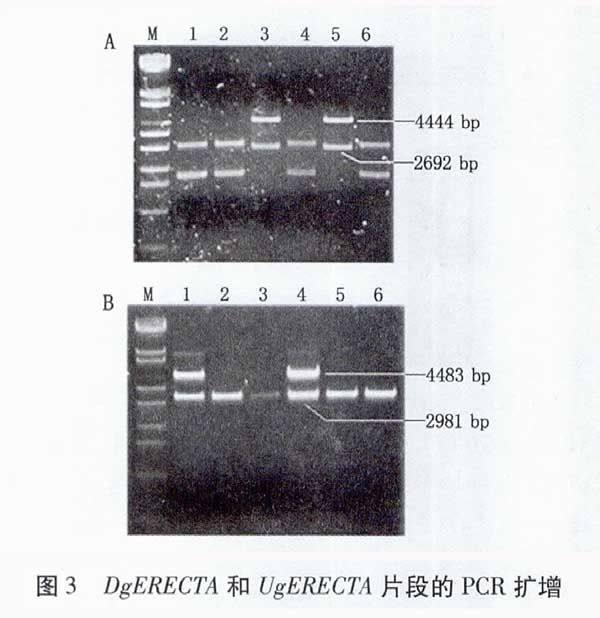
將DgERECTA與pMD18-T載體連接,挑取經過Amp抗性篩選的6個單菌落,搖菌過夜后提取質粒,用Hind Ⅲ和SpeⅠ雙酶切,10%(W/V)的瓊脂糖凝膠電泳檢測。預測陽性克隆酶切后應出現兩條帶,一條為2 692 bp的pMD18質粒片段;一條為4 444 bp的DgERECTA目的片段(引物P4到Hind Ⅲ之間的長度)。結果(圖3A)表明,3、5號單菌落質粒出現了與預期結果相符的兩條帶,為陽性克隆。將酶切正確的重組克隆質粒進行測序,測序結果與已報道的序列完全匹配,并帶有完整的酶切位點。表明酶切檢測正確的重組克隆質粒pMD18-DgERECTA中含有正確的目的基因片段。
24 重組質粒pTA2-UgERECTA單酶切鑒定
將UgERECTA與pTA2-T載體連接,挑取經過Amp抗性篩選的6個單菌落,搖菌過夜后提取質粒,用Hind Ⅲ單酶切,10%(W/V)的瓊脂糖凝膠電泳檢測。預測陽性克隆酶切后應出現兩條帶,一條為2 981 bp的pTA2質粒片段;一條為4 483 bp的UgERECTA目的片段。結果(圖3B)表明,1、4號單菌落質粒出現了與預期結果相符的兩條帶,為陽性克隆。將酶切正確的重組克隆質粒進行測序,比對結果表明插入的UgERECTA序列與已報道的序列完全匹配,并帶有完整的酶切位點。表明酶切檢測正確的重組克隆質粒pTA2-UgERECTA中含有正確的目的基因片段。
25 重組表達質粒pCAMBIA1390-gERECTA的構建
由于gERECTA基因片段較長,直接同pCAMBIA1390載體連接較為困難。因此,我們利用gERECTA基因序列內部的Hind Ⅲ酶切位點和人為添加的SpeⅠ 酶切位點,先將DgERECTA連接到pCAMBIA1390,再利用Hind Ⅲ酶切位點將UgERECTA連接到重組質粒pCAMBIA1390-DgERECTA上。由于插入的UgERECTA序列帶有自身的啟動子序列,所以gERECTA的表達受自身啟動子的調控。
251 重組質粒pCAMBIA1390-DgERECTA的酶切鑒定 經Kan抗性篩選后的單菌落搖菌過夜后提取質粒,用Hind Ⅲ和Spe Ⅰ雙酶切,10%(W/V)的瓊脂糖凝膠電泳檢測。預測陽性質粒酶切后應出現兩條帶,一條為8 864 bp的pCAMBIA1390質粒片段;一條為4 444 bp的DgERECTA目的片段。結果(圖4A)表明,1~3號單菌落質粒出現了與預期結果相符的兩條帶,為陽性重組質粒。
252 重組質粒pCAMBIA1390-DgERECTA-UgERECTA的酶切鑒定 由于UgERECTA目的片段同pCAMBIA1390-DgERECTA連接反應是利用HindⅢ單一位點進行連接,所以pCAMBIA1390-DgERECTA重組載體去磷酸化反應要徹底,盡量減少載體的自連。連接反應前利用瓊脂糖凝膠電泳檢測回收UgERECTA目的片段和去磷酸化后的pCAMBIA1390-DgERECTA重組載體(圖4B)。
對經Kan抗性篩選后的單菌落搖菌過夜后提取質粒,用Hind Ⅲ單酶切,10%(W/V)的瓊脂糖凝膠電泳檢測。預測質粒酶切后應出現兩條帶,一條為13 308 bp的pCAMBIA1390-DgERECTA質粒片段;一條為4 483 bp的UgERECTA目的片段。結果(圖5A)表明,1、2、5、6、7號質粒為陽性重組質粒。由于這步連接反應只利用了Hind Ⅲ單一酶切位點,為檢測UgERECTA片段插入到pCAMBIA1390-DgERECTA重組質粒上的方向性,我們利用SpeⅠ酶切檢測1、2、5、6、7號質粒,預測連接方向正確的陽性質粒應出現一條7 960 bp的條帶,連接方向顛倒的質粒出現一條5 483 bp的條帶。結果(圖5B)表明,2、5、7號質粒為連接正確的陽性重組質粒。
3 討論
近年來,全球水資源短缺境況越來越嚴重,干旱造成農作物減產導致的經濟損失逐年加重。植物可以通過誘導可溶性滲透調節物質的積累、膨壓的維持或者耐脫水等抵抗干旱[13],但是僅靠現有植物資源本身的調節能力抵御干旱已不能滿足農業高產和穩產的需要。基因工程技術已逐漸滲透到植物的遺傳改良研究中,與傳統抗旱育種方法相比,基因工程以其周期短、效率高等特點得到越來越廣泛的應用。
已有研究表明擬南芥ERECTA基因可以通過調控蒸騰效率來增強植株耐干旱脅迫的能力[11]。水稻是我國重要的糧食作物,改良水稻耐旱性是科學家育種的重要目標之一。構建能在水稻中表達OsERECTA基因的植物表達載體為培育抗旱水稻新品種提供了可能。為了便于研究OsERECTA基因在水稻生長發育中的作用,構建OsERECTA基因植物表達載體時要排除外源啟動子對OsERECTA基因表達的影響。Dievart和Song等證明擬南芥ERECTA基因的啟動子可表達擬南芥CLAVATA1和LhG4基因[14,15],是相對較強的啟動子。這同Karve等認為擬南芥ERECTA基因的上游序列作為強啟動子起作用的結論一致[12]。因此,我們構建的pCAMBIA1390-gERECTA載體攜帶的OsERECTA自身啟動子可以保證gERECTA基因過量表達,并具有同內源OsERECTA基因相同的表達模式,例如組織和器官特異性等,這比使用組成型啟動子調控gERECTA基因的表達更便于研究OsERECTA基因的功能。
本實驗通過基因工程技術手段,以日本晴全基因組DNA為模板,通過分段PCR擴增,克隆亞片段再連接的策略,成功分離并構建了包含水稻OsERECTA基因全長基因組片段的植物表達載體pCAMBIA1390-gERECTA,后續的基因轉化工作正在進行,OsERECTA基因將在其自身啟動子的控制下在轉基因水稻中表達。本實驗OsERECTA基因植物表達載體的構建為研究OsERECTA過表達對水稻生長發育及環境適應性方面的影響奠定了基礎,也為研究水稻OsERECTA的功能奠定了基礎。參 考 文 獻:
[1] Torii K U, Mitsukawa N, Oosumi T, et al The Arabidopsis ERECTA gene encodes a putative receptor protein kinase with extracellular leucine-rich repeats [J] Plant Cell, 1996, 8(4): 735-746
[2] Yokoyama R, Takahashi T, Kato A, et al The Arabidopsis ERECTA gene is expressed in the shoot apical meristem and organ primordia [J] Plant J, 15(3): 301-310
[3] Shpak E D, Berthiaume C T, Hill E J, et al Synergistic interaction of three ERECTA-family receptor-like kinases controls Arabidopsis organ growth and flower development by promoting cell proliferation [J] Development, 2004, 131(7): 1491-1501
[4] Woodward C, Bemis S M, Hill E J, et al Interaction of auxin and ERECTA in elaborating Arabidopsis inflorescence architecture revealed by the activation tagging of a new member of the YUCCA family putative flavin monooxygenases [J] Plant Physiol, 2005, 139(1): 192-203
[5] Pillitteri L J, Bemis S M, Shpak E D, et al Haploinsufficiency after successive loss of signaling reveals a role for ERECTA-family genes in Arabidopsis ovule development [J] Development, 2007, 134(17): 3099-3109
[6] Masle J, Gilmore S R, Farquhar G D The ERECTA gene regulates plant transpiration efficiency in Arabidopsis [J] Nature, 2005, 436: 866-870
[7] Qi Y, Sun Y, Xu L, et al ERECTA is required for protection against heat-stress in the AS1/AS2 pathway to regulate adaxial-abaxial leaf polarity in Arabidopsis [J] Planta, 2004, 219(2): 270-276
[8] Godiard L, Sauviac L, Torii K U, et al ERECTA, an LRR receptor-like kinase protein controlling development pleiotropically affects resistance to bacterial wilt [J] Plant J, 2003, 36(3): 353-365
[9] Xing H T, Guo P, Xia X L, et al PdERECTA, a leucine-rich repeat receptor-like kinase of poplar, confers enhanced water use efficiency in Arabidopsis [J] Planta, 2011, 234(2): 229-241
[10] 朱錦程,沈海濤,祝建波擬南芥ERECTA基因的克隆及其對番茄轉化 [J] 生物技術通報, 2010, 8: 102-105
[11] Callis J, Fromm M, Walbot V Introns increase gene expression in cultured maize cells [J] Genes Dev,1987,1:1183-1200
[12] Karve R, Liu W, Willet S G, et al The presence of multiple introns is essential for ERECTA expression in Arabidopsis [J] RNA, 2011, 17(10): 1907-1921
[13] Bhatnagar-Mathur P, Vadez V, Sharma K K Transgenic approaches for abiotic stress tolerance in plants: retrospect and prospects [J] Plant Cell Rep, 2008, 27(3): 411-424
[14] Diévart A, Dalal M, Tax F E, et al CLAVATA1 dominant-negative alleles reveal functional overlap between multiple receptor kinases that regulate meristem and organ development [J] Plant Cell, 2003, 15(5): 1198-1211
[15] Song S K, Lee M M, Clark S E POL and PLL1 phosphatases are CLAVATA1 signaling intermediates required for Arabidopsis shoot and floral stem cells [J] Development, 2006, 133(23): 4691-4698
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
