TRAIL誘導惡性膠質瘤LN215細胞凋亡抵抗的機制
2013-05-29 09:17:44張艷春于洪泉趙東海劉興吉
中國老年學雜志 2013年6期
關鍵詞:生長
張艷春 于洪泉 王 猛 趙東海 金 宏 齊 玲 劉興吉
(吉化集團公司總醫院麻醉科,吉林 吉林 132013)
惡性膠質瘤目前尚缺乏理想的治療方法〔1〕。腫瘤壞死因子相關凋亡誘導配體(TRAIL)作為一種新型的抗腫瘤藥物,在美國已進入臨床Ⅱ期實驗階段〔2〕。但大部分惡性膠質瘤細胞株對TRAIL誘導凋亡發生抵抗〔3〕,其機制仍然不清。本文對惡性膠質瘤細胞LN215細胞和單克隆細胞進行研究,為TRAIL作為腫瘤治療新藥提供實驗依據。
1 材料與方法
1.1 主要試劑 重組人TRAIL(rhTRAIL)(Peprotech公司);DMEM/F12培養基、小牛血清(Gibco公司);對硝基苯(上海生物工程技術公司);兔抗人第二個線粒體激活因子(SMAC)多克隆抗體(Biomol公司);兔抗人BH3結構域凋亡誘導蛋白(BID)多克隆抗體(Invitrogen公司);兔抗人肌動蛋白(actin)多克隆抗體(Sigma公司);過氧化物酶標記IgG第二抗體(Jackson Immunoresearch公司)。
1.2 細胞培養及分離單克隆細胞 將LN215細胞接種于含10%小牛血清的DMEM/F12培養基中培養,待細胞匯合至80%~90%,用0.25%胰酶消化,臺盼藍計數后稀釋。按1~2個/孔細胞密度接種于96孔板中進行培養。培養條件為5%CO2,37℃孵箱。每天觀察生長情況,標記出含單個細胞的孔。待其匯合,轉入24孔板中進行培養。每周換液2次,90%匯合后轉入65 mm培養皿中進行培養。
1.3 酸性磷酸酶法檢測細胞凋亡的百分率 待細胞融合至85%,0.25%胰酶消化,臺盼藍計數活細胞后,以1×104個/孔細胞密度接種于96孔板中,培養24 h,將細胞分為11組,加入TRAIL(0.01,0.03,0.1,0.3,1,3,10,30,100,300 ng/ml),對照組使用基礎培養基。24 h后小心棄上清,磷酸鹽緩沖液(PBS)洗去培養基,每孔加入酸性磷酸酶底物檢測液(對硝基苯新鮮配制)100μl,繼續培養90 min,加入終止液,室溫下孵育5 min。酶標儀405 nm處測定各孔光吸收值。按照公式計算細胞凋亡率=(1-藥物組光吸收值/對照組光吸收值)×100%。
1.4 Western印跡法檢測蛋白的表達 收集細胞,3 000 r/min離心后棄上清。PBS洗滌細胞離心后棄上清,每個管中加入50μl細胞裂解液,12 000 r/min 4℃ 離心 15 min,棄去沉淀,Braford測定法檢測樣品蛋白濃度,十二烷基硫酸鈉-聚丙烯酰胺凝膠(SDS-PAGE)電泳分離樣品,4℃轉膜封閉,加入一抗4℃過夜。加入二抗,室溫、避光孵育1 h,膠片曝光拍照。
1.5 統計學分析 采用SPSS11.0軟件進行分析,計量資料以±s表示,行t檢驗。
2 結果
2.1 LN215細胞及單克隆細胞的培養 惡性膠質瘤LN215細胞經復蘇后培養,細胞呈現長梭形,有的可見明顯的突起。觀察96孔板內含單細胞孔細胞的生長狀態發現:雖然細胞來自于同一株細胞,但各單克隆細胞生長速度不同、形態也不全相同。待細胞生長至90%時,挑選出生長速度與形態均不相同的32株細胞,用胰酶消化轉入24孔板生長,細胞生長狀態良好。比較后發現有5株單克隆細胞生長形態、生長速度不同,編號分別是1、6、17、13和32。這5株細胞用于后續實驗。
2.2 LN215細胞和單克隆細胞對TRAIL誘導凋亡的敏感性LN215和5株單克隆細胞經不同濃度TRAIL作用后,細胞凋亡率在 -10.2% ~55.7%之間波動,LN215、1、6、17、13 和 32 細胞的最大凋亡率分別為12.1%、3.48%、17.4%、4.85%、55.7%和28.1%,其中6、13和32經TRAIL作用后發生凋亡較LN215更明顯,而1和7則相反,以13對TRAIL誘導凋亡最為敏感。見圖1。
2.3 LN215細胞和單克隆細胞TRAIL相關蛋白的表達TRAIL相關蛋白均有表達。LN215細胞和單克隆細胞表達DR4與細胞凋亡百分率不相關;BID、SMAC表達與細胞凋亡百分率明顯相關,6、13、32較1和17表達量明顯增加,尤其是13最為明顯。見圖2。
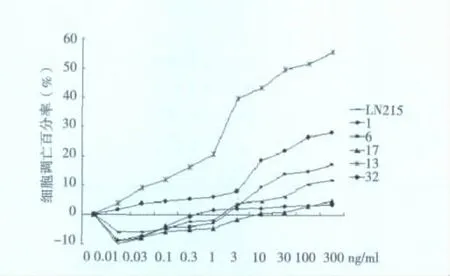
圖1 TRAIL誘導LN215及單克隆細胞發生凋亡情況
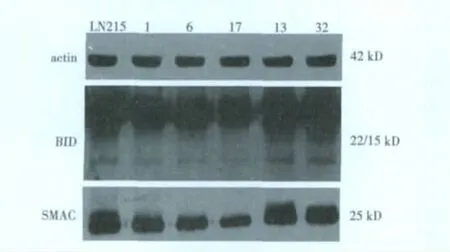
圖2 惡性膠質瘤LN214細胞及單克隆細胞TRAIL相關蛋白的表達
3 討論
膠質瘤是起源于中樞神經系統的最常見腫瘤,由于其發病隱匿,一經發現病理診斷就是Ⅲ~Ⅳ期膠質瘤,給臨床治療帶來了很大困難。并且由于腦膠質瘤在腦內呈彌散分布〔4〕,手術難以清除。這可能是由于激活細胞生長途徑和/或抑制細胞凋亡途徑而對治療產生抵抗〔5〕。TRAIL可以誘導膠質瘤細胞凋亡〔6,7〕,TRAIL誘導凋亡的非線粒體途徑需要線粒體途徑的補充,由線粒體途徑通過(caspase 8)間接激活BID,再依次與Bax和Bak作用,改變了線粒體膜的生物學特性,促使細胞色素C和SMAC等線粒體蛋白釋放。在胞質內,SMAC與相關蛋白作用解除抑制因子的抑制作用,使TRAIL誘導的非線粒體途徑得以進行〔8,9〕,細胞發生凋亡。
根據腫瘤發生可能來源于多個細胞的突變,本實驗利用細胞梯度稀釋方法由LN215分離出5株單克隆細胞。LN215細胞和單克隆細胞BID、SMAC表達與細胞凋亡百分率明顯相關。因此,筆者認為LN215和單克隆細胞經TRAIL誘導凋亡百分率與BID和SMAC表達量密切相關,說明TRAIL誘導LN215和單克隆細胞可能是通過BID和SMAC引起,表達量的減少可能會引起TRAIL誘導凋亡抵抗,但更進一步的機制仍待研究。
1 Stupp R,Mason WP,van den Bent MJ,et al.Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma〔J〕.N Engl J Med,2005;352(10):987-96.
2 Bellail AC,Qi L,Mulligan P,et al.TRAIL agonists on clinical trials for cancer therapy:the promises and the challenges〔J〕.Revi Rec Clin Trials,2009;(4):34-41.
3 Li YC,Tzeng CC,Song JH,et al.Genomic alterations in human malignant glioma cells associate with the cell resistance to the combination treatment with tumor necrosis factor-related apoptosis-inducing ligand and chemotherapy〔J〕.Clin Cancer Res,2006;12(9):2716-29.
4 Holland EC.Glioblastoma multiforme:the terminator〔J〕.Proc Natl Acad Sci USA,2000;97(12):6242-44.
5 Lefranc F,Brotchi J,Kiss R.Possible future issues in the treatment of glioblastomas:special emphasis on cell migration and the resistance of migrating glioblastoma cells to apoptosis〔J〕.J Clin Oncol,2005;23(10):2411-22.
6 南栗巖,齊 玲,肖振晶,等.原代培養的惡性膠質瘤細胞表達死亡受體與TRAIL抵抗的關系〔J〕.中國老年學雜志,2011;18(1):3591-2.
7 齊 玲,于洪泉,丁麗娟,等.Caspase-8在膠質母細胞瘤抵抗TRAIL誘導凋亡中的作用〔J〕.吉林大學學報(醫學版),2011;37(4):612-6.
8 Li H,Zhu H,Xu CJ,et al.Cleavage of BID by caspase8 mediates the mitochondrial damage in the Fas pathway of apoptosis〔J〕.Cell,1998;94(8):491-501.
9 Du C,Fang M,Li Y,et al.Smac,a mitochondrial protein that promotes cytochrome c-dependent caspase activation by eliminating IAP inhibition〔J〕.Cell,2000;102(1):33-42.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
