青枯菌種內分型研究進展
2013-09-13 11:46:46蔡劉體汪漢成劉艷霞曹毅袁賽飛趙準備石俊雄
生物技術通報 2013年7期
蔡劉體 汪漢成 劉艷霞 曹毅 袁賽飛 趙準備 石俊雄
青枯菌(Ralstonia solanacearum)是一種土傳性的細菌性病原菌,屬于革蘭氏陰性細菌,廣泛分布于熱帶、亞熱帶及一些溫帶地區,它具有廣泛的寄主,可以侵染 50多個科的 200多種植物[1],包括煙草(Nicotiana tabacum)、芝麻(Sesamum indicum)、花生(Arachishypogaea)、大豆(Glycinemax)、蘿卜(Raph-anus sativus)等多種農作物以及一些貴重藥材和花卉植物,嚴重危害一些農作物的產量,其中馬鈴薯、番茄、煙草等茄科作物受害最為嚴重[2]。
青枯菌是一個復雜的菌種[3],寄主范圍寬、分布廣泛,具有高度的變異性和適應性,不同地域或不同寄主的菌株間有明顯的變異和分化現象[3,4],而且青枯菌生理生化致病途徑眾多[5]。為了探索青枯菌引起的病害、青枯病害的流行規律,更好地指導青枯病害的防控及經濟作物的抗病育種,國內外研究者開展了青枯菌內亞分類或菌系分群的研 究 工 作。 前 人 從 生 理 小 種(races)[5]、 生 化型(biovars)[6]、 血 清 型(serotypes)[7]、 溶 源 型(lysotypes)[8]、脂肪酸型[9-11]、基因型(genetypes)[12]、種系型[3]等多方面對種內分型進行研究,目前大多數認同的分型方法主要有生理小種、生化型和種系型(phylotypes)3種分型方法。本文綜述青枯菌種內分型的主要研究進展,并對不同的分型進行比較,期望為煙草青枯菌系的構成和遺傳多樣性分析,以及煙草與青枯菌互作研究提供參考。
1 青枯菌種內分型方法
1.1 生理小種
青枯菌具有廣泛的寄主,按照不同來源的青枯菌株系對不同植物種類的致病性差異,早在20世紀60、70年代,Buddenhagen等[13-17]制定了青枯菌生理小種的劃分方法,把青枯菌劃分為5個生理小種[18]:(1)生理小種 1(race1):可侵染茄科植物和其他科植物的青枯菌;(2)生理小種2(race2):只侵染香蕉(Musa paradisiaca)、大蕉(Musa sapientum L.)和Heliconia的青枯菌;(3)生理小種3(race3):只侵染馬鈴薯,偶爾侵染番茄和茄子的青枯菌;(4)生理小種4(race4):只對姜(Zingiber officinale)有很強致病力的青枯菌;(5)生理小種5(race5):特異性侵染桑屬植物的青枯菌。按照寄主把青枯菌分為不同的生理小種對作物抗病育種非常有用,而且目前青枯菌生理小種的劃分仍然延用該方法,但是由于青枯菌寄主范圍廣泛,按照寄主劃分的方法工作量大,較耗費時間[19],而且有同一青枯菌株系同時被劃歸到幾個生理小種的情況出現。
1.2 生化型
青枯菌的另一個亞分類系統主要是由Hayward等[20,21]提出的,他們根據青枯菌對3種雙糖(麥芽糖、乳糖和纖維二糖)和3種己醇(甘露醇、山梨醇和半乳糖醇)氧化產酸能力的差異,將從各大洲收集的185個青枯菌分為4個生化型:生化型1(不能氧化3種雙糖和3種己醇)、生化型2(只能氧化3種雙糖,不能氧化3種己醇)、生化型3(能氧化3種雙糖和3種己醇)、生化型4(只能氧化3種己醇,不能氧化3種雙糖)。Hayward等[22]在此基礎上,依據青枯菌利用硝酸鹽還原反應和利用硝酸鹽產生氣體情況,將青枯菌劃分為6個生化型。表1顯示按照對糖、醇以及硝酸鹽的利用情況,將青枯菌劃分為5個生化型,但根據其對海藻糖、肌糖、D-核糖以及果膠酶的利用情況,把生化型2分為生化型2A和生化型2T/2N(表2),其中生化型2A包括有RFLP分型的 26組和27組。青枯菌株系生化型的研究報道目前仍沿用[23],鄒陽等[24]采用該方法分析了中國重慶地區煙草青枯菌的生化型。該方法分析青枯菌的生化型相對而言,工作量較小且較短時間獲得結果,但是也有出現個別青枯菌株系不好劃分的情況[25]。
1.3 種系型(或稱為演化型)
隨著分子生物學和分子進化技術的發展,青枯菌研究者在青枯菌生理小種和生化型劃分的基礎上,期望了解青枯菌的種系結構、遺傳背景、地理起源及其與復雜群體的相關性,Fegan和Prior[3]共同提出了以親緣關系為基礎的劃分體系,將青枯菌分成4個種系型,每一個種系型又繼續分成更小的群體,稱為序列型(sequevars)。在此劃分框架中,種系型定義為菌株的16S-23S rRNA 基因間間隔區(ITS),hrp(超敏反應蛋白基因)和egl(內切葡聚糖酶基因)的序列數據,通過系統發育分析顯示為一個單系群。按照這種劃分,每個種系型反應了菌株的地理起源,其中種系型Ⅰ和Ⅱ分別由亞洲和美國菌株組成,種系型Ⅲ的成員由非洲菌株組成,種系型Ⅳ包括來自印度尼西亞和澳大利亞的 R.syzygii和 BDB[3,18]。青枯菌種系型是根據青枯菌特異性引物,通過PCR擴增的帶型特征來劃分,青枯菌種系型的劃分方法見表3,一個青枯菌株系的種系型可以基于IST區間序列信息設計引物的基礎上,通過復合PCR快速的鑒定出來[3]。徐進等[26]采用復合PCR檢測青枯菌種系型的分析方法,分析了福建主要煙區的45個煙草青枯菌株系都屬于種系型I;潘哲超等[27]基于內切葡萄糖酶基因系統發育學的分析,結果表明來自福建和部分貴州的62個煙草青枯菌株系均歸屬于青枯菌亞洲分支(種系型I)。
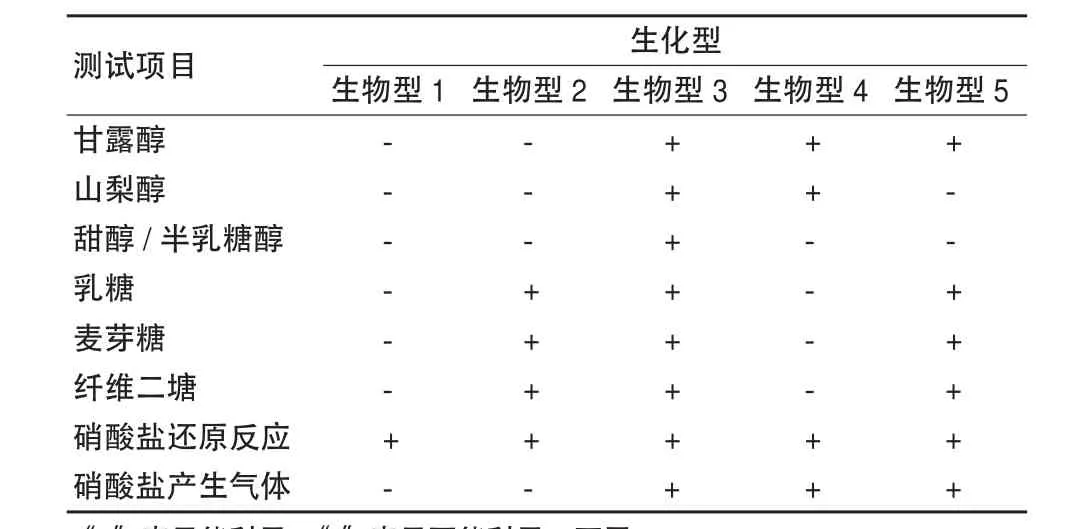
表1 青枯菌不同生化型的劃分[20]

表2 青枯菌生化型II的亞型劃分

表3 基于ITS區設計的用于青枯菌種系型分析的特異性條帶PCR擴增的引物信息[3,18]
2 青枯菌分型的比較
與生理小種和生化型劃分相比較,種系型的劃分框架似乎能更準確的反映出目前已知青枯菌的遺傳多樣性和復雜性。在生理小種劃分框架中,生理小種1(R1)定義為“可侵染茄科植物和其他科植物”的青枯菌株系包含非常寬泛,從種系型看,可以歸為R1的包含有種系型I-種系型IV的青枯菌株系;而生理小種2(R2)和生理小種3(R3)的寄主范圍很窄,在生理小種劃分框架中所反映的遺傳多樣性也比較窄,R2和R3在種系型框架中都是屬于種系型II。在生化型劃分框架中,生化型1和2T青枯菌株系包含有4個種系型中的3個,而且在青枯菌株系劃分中,僅僅知道其是生化型1和2T也不能提供該株系的更多信息。
種系型劃分框架,首先基于對菌株的16S-23S rRNA 基因間間隔區(ITS),hrp和egl基因的序列數據分析,通過系統發育分析定義單系群,然后從基因序列水平上來反應每個種系型與菌株的地理起源和遺傳信息的對應關系,分為4個種系型。劃分框架中每個種系型中都可以細分為序列型,使其成為開放性的劃分框架,有利于對新發現的青枯菌株系進行劃分,也有利于其自身信息和劃分框架的完善。然而,無論是生理小種、生化型還是種系型的劃分框架,都不能直接反映出青枯菌株系的致病性和致病力,許多研究表明采用的種下分型的分析方法與青枯菌的致病性無特定的相關性[28]。
3 小結
青枯菌這個復雜種至少包含有4個遺傳群體或種系型,反映出其遺傳多樣性和地理起源信息。人們通過各種方法把青枯菌分型是為了作物育種工作者、植物病理學者和植物保護人員能更好地了解青枯菌的寄主、分布、遺傳及致病力等信息,減少或控制農作物的經濟損失。隨著收集的青枯菌資源不斷豐富,可能會有越來越多的青枯菌株系劃分到各個種系型中,序列型數目會越來越豐富。收集和分類青枯菌資源非常重要,但更重要的是了解青枯菌株系的生態、進化、遺傳及致病力等特性,這些信息對于控制青枯病危害的研究尤其重要。近年來,已經報道完成了青枯菌多個株系的全基因組的測序[29-34],這些青枯菌全基因組測序信息的解析和積累,青枯菌致病基因家族信息分析及青枯菌致病途徑的闡述,將有助于青枯菌種內分型的推進,及青枯菌與宿主相互作用的研究和青枯病的防控研究。
[1] Hayward AC.Biology and epidemiology of bacterial wilt caused by Ralstonia solanacearum[J].Annu Rev Phytopathol, 1991, 29:65-87.
[2] Hayward A.Thehosts of Pseudomonas solanacearum[M]//Hayward AC, Hartman GL.Bacterial wilt:the disease and its causative agent Pseudomonas solanacearum.UK:CAB International, Oxford, 1994:9-24.
[3] Fegan M, Prior P.How complex is the Ralstonia solanacearum species complex[M]//Allen C, Prior P, et al.Bacterial wilt disease and the Ralstonia solanacearum species complex.St.Paul, MN:American Phytopathological Society Press, 2005:449-461.
[4] Hayward AC.Biology and epidemiology of bacterial wilt caused by Pseudomonas solanacearum[J].Annual Review of Phytopathology,1991, 29:65-87.
[5] Poueymiro M, Genin S.Secreted proteins from Ralstonia solanacearum:ahundred tricks to kill a plant[J].Current Opinion in Microbiology, 2009, 12(1):44-52.
[6] Buddenhagen I, Kelman A.Biological and physiological aspects of bacterial wilt caused by Pseudomonas solanacearum[J].Annual Review of Phytopathology, 1964, 2:203-230.
[7] 徐麗慧, 徐福壽, 李芳, 等.桑細菌性青枯病病原及其生化型鑒定[J].植物保護學報, 2007, 34(2):141-146.
[8] Malandrin L, Samson R.Serological andmolecular size characterization of flagellins of Pseudomonas syringae pathovars and related bacteria[J].Systematic and Applied Microbiology, 1999, 22(4):534-545.
[9] 許東, 賴文姜, 范懷忠.桑青枯菌血清型與其它分型的比較研究[J].植物病理學報, 1986, 16(1):29-36.
[10] Khan AA, Furuya N, Matsumoto M, Matsuyama N.Identification of Ralstonia solanacearum isolated from wilted tobacco plant by fatty acid profiles and PCR-RFLP analysis[J].Journal of the Faculty of Agriculture, Kyushu University, 1999, 44(1-2):59-65.
[11] Krej?í E, Kroppenstedt RM.Differentiation of species combined into the Bukholderia cepacia complex and related taxa on the basis of their fatty acid patterns[J].Journal of Clinical Microbiology,2006, 44(3):1159-1164.
[12] 劉波, 藍江林, 車建美, 等.青枯雷爾氏菌脂肪酸型與致病性的關系[J].中國農業科學, 2009, 42(2):511-522.
[13] Hartung F, Wemer R, Muhlbach HP, Buttner C.Highly specific PCR-diagnosis to determine Pseudomonas solanacearum strains of different geographic origins[J].Theoretical and Applied Genetics, 1998, 96(6-7):797-802.
[14] Buddenhagen IW, Sequeira L, Kelman A.Designation of physiological races in Pseudomonas solanacearum[J].Phytopathology,1962, 52:726.
[15] French ER, Sequeira L.Strains of Pseudomonas solanacearu from central and south America:a comparative study[J].Phytopathology, 1970, 60:506-512.
[16] Sequeira L, Averre CW.Distribution and pathogenicity of strains of Pseudomonas solanacearum from virgin soils in Costa Rica[J].Plant Dis Rep, 1961, 45:435-440.
[17] Lozano JC, Sequeira L.Differention of physiological races of Pseudomonas solanacearum by a leaf infiltration technique[J].Phytopathology, 1970, 60:833-838.
[18] Denny TP.Plant pathogenic Ralstonia species[M]//Gnanamanickam SS.Plant-Associated Bacteria.Dordrecht, The Netherla-nds:Springer Publishing, 2006:573-644.
[19] Prior P, Fegan M.Recent developments in the phylogeny and classification of Ralstonia solanacearum[J].Acta Hortic, 2005,695:127-136.
[20] Hayward AC.Characteristics of Pseudomonas solanacearum[J].J Appl Bact, 1964, 27:265-277.
[21] He LY, Sequeira L, Kelman A.Characteristics os strains of Pseudomonas solanacearum[J].Plant Dis, 1983, 67:1357-1361.
[22] Hayward AC.Ralstonia solanacearum[M]Lederberg J.Encyclopedia ofmicrobiology.San Diego, CA:Academic Press, 2000:32-42.
[23] Stevens P, van Elsas JD.Genetic and phenotypic diversity of Ralstonia solanacearum biovar 2 strains obtained from Dutch waterways[J].Antonie Van Leeuwenhoek, 2010, 97(2):171-188.
[24] 鄒陽, 肖崇剛, 易龍.重慶地區煙草青枯病菌的生物型測定[J].煙草科技, 2007, 6:61-64.
[25] Guidot A, Elbaz M, Carrere S, et al.Specific genes from the potato brown rot strains of Ralstonia solanacearum and their potential use for strain detection[J].Phytopathology, 2009, 99:1105-1112.
[26] 徐進, 顧剛, 潘哲超, 等.福建煙草青枯菌演化型及生化變種鑒定研究[J].中國煙草學報, 2010, 16(6):66-71.
[27] 潘哲超, 徐進, 顧鋼, 等.福建及貴州等地煙草青枯菌系統發育分析[J].植物保護, 2012(1):18-23.
[28] Stefani E, Giosue S, Mazzucchi U.Detection of latent infections of Ralstonia solanacearum biovar 2, race 3 in tomato crops[J].Journal of Plant Pathology, 2005, 87(3):167-171.
[29] Salanoubat M, Genin S, et al.Genome sequence of the plant pathogen Ralstonia solanacearum[J].Nature, 2002, 415:497-502.
[30] Remenant B, et al.Genomes of three tomato pathogens within the Ralstonia solanacearum species complex reveal significant evolutionary divergence[J].BMC Genomics, 2010, 11:379.
[31] Genin S.Molecular traits controllinghost range and adaptation to plants in Ralstonia solanacearum[J].New Phytologist, 2010,187(4):920-928.
[32] Milling A, Babujee L, Allen C.Ralstonia solanacearum extracellular polysaccharide is a specific elicitor of defense responses in wiltresistant tomato plants[J].PLoS ONE, 2011, 6(1):e15853.
[33] Poueymiro M, Genin S.Secreted proteins from Ralstonia solanacearum :ahundred tricks to kill a plant[J].Current Opinion in Microbiology, 2009, 12(1):44-52.
[34] Li ZF, Wu SL, Bai XF, et al.Genome sequence of the tobacco bacterial wilt pathogen Ralstonia solanacearum[J].Journal of Bacteriology, 2011, 193(21):6088-6089.
