牦牛HSFY基因的克隆及其結(jié)構(gòu)分析
2013-09-13 11:46:52曾賢彬王永劉仲娜馬志杰海汀鐘金城
生物技術(shù)通報(bào) 2013年7期
關(guān)鍵詞:結(jié)構(gòu)功能
曾賢彬 王永 劉仲娜 馬志杰 海汀 鐘金城
熱休克轉(zhuǎn)錄因子HSFY(heat shock transcription factor,Y chromosome)是哺乳動(dòng)物4種熱休克因子(HSF1、HSF2、HSF4和HSFY)之一,控制熱休克蛋白表達(dá)[1]。HSFY 與其它 HSFs(HSF1和HSF2)具有不同的功能[2]。Tessari等[3]將 HSFY作為人精子發(fā)生障礙的候選基因之一。Giovanna[4]、Shinka[2]、Ferlin[5]等報(bào)道 HSFY 基因缺失與無(wú)精子、少精子癥緊密相關(guān)。
Tessari[3]、Christine[6]等對(duì)人、恒河猴、普通牛、家貓、挪威鼠的HSFY蛋白質(zhì)序列進(jìn)行了多序列比對(duì),結(jié)果顯示,普通牛與家貓親緣關(guān)系最近,兩物種間的該基因編碼區(qū)和與DNA結(jié)合的結(jié)構(gòu)域的序列一致性分別為58%和81%。目前認(rèn)為,HSFY基因是Y染色體上的多拷貝基因。人有6個(gè)拷貝,其中兩個(gè)功能拷貝;家貓有8個(gè)功能拷貝,拷貝數(shù)未知;普通牛約有73個(gè)拷貝,功能基因數(shù)未知[3,6,7]。人、普通牛的HSFY主要在睪丸中特異表達(dá)[3,8],即HSFY僅在支持細(xì)胞和精母細(xì)胞中表達(dá)[2],其mRNA表達(dá)水平與精原細(xì)胞和精母細(xì)胞標(biāo)記(UCHL1和 TRPC2)mRNA 表達(dá)水平顯著相關(guān)(P<0.0001)[6]。用抗HSFY抗體證實(shí),正常精子生成時(shí),HSFY在曲細(xì)精管的生殖細(xì)胞和支持細(xì)胞中表達(dá)。精原細(xì)胞期、精母細(xì)胞細(xì)線期均無(wú)HSFY表達(dá)信號(hào);偶線期染色質(zhì)周?chē)袕?qiáng)HSFY表達(dá)信號(hào),但在精子成熟后HSFY蛋白量減少[9],表明其與精子生成有關(guān),但其生物學(xué)功能尚待研究[2-4,10]。
有關(guān)牦牛HSFY基因的研究尚未見(jiàn)報(bào)道,本研究對(duì)牦牛的HSFY基因進(jìn)行克隆測(cè)序、結(jié)構(gòu)和密碼子偏好性分析以及與普通牛等其他物種相應(yīng)基因進(jìn)行比對(duì)研究,以期為進(jìn)一步研究牦牛HSFY基因的結(jié)構(gòu)和功能提供理論依據(jù)。
1 材料與方法
1.1 材料
在四川省龍日種畜場(chǎng)選取健康成年公牦牛兩頭,采取耳組織,置于75%乙醇中帶回實(shí)驗(yàn)室,保存于-20℃冰箱中備用。
1.2 方法
1.2.1 基因組DNA的提取 按試劑盒(天根公司)操作說(shuō)明提取基因組DNA。
1.2.2 牦牛HSFY基因克隆測(cè)序 根據(jù)普通牛HSFY2序列(NC_016145.1)設(shè)計(jì)引物,上下游引物分別為F:5'-TCACAGCCTTTTGACTTTG-3'、R:5'-CTCTTTTCCCTTGCTTCTC-3';引物由上海Invitrogen公司合成。
25μL PCR反應(yīng)體系:基因組 DNA 1μL、上下游引物各1μL、ddH2O 9.5μL 和 2×long Taq酶(博奧維新公司)12.5μL 。PCR反應(yīng)程序:95℃預(yù)變性3min;95℃變性30s,56℃復(fù)性30s,72℃延伸1.5min,共30個(gè)循環(huán);72℃終延伸10min,4℃保存。
經(jīng)膠回收試劑盒回收PCR產(chǎn)物,與pMD19-T(購(gòu)自TaKaRa公司)于16℃過(guò)夜連接,其產(chǎn)物轉(zhuǎn)化大腸桿菌(DH5α)感受態(tài)細(xì)胞,涂布于含50μg/mL氨芐青霉素、200mg/mL IPTG及20mg/mL X-Gal的LB平板,過(guò)夜培養(yǎng)。經(jīng)PCR篩選陽(yáng)性克隆進(jìn)行測(cè)序。
1.2.3 序列分析 測(cè)序結(jié)果用DNAMAN4.0、DNAssist1.0等生物信息學(xué)軟件進(jìn)行拼接、比對(duì)分析,用 GENSCAN(http://genes.mit.edu/GENSCAN.html)預(yù)測(cè)蛋白的氨基酸序列,并與普通牛(NP_001070-474)、瘤牛(AFV09891)、人(hHSFY1:NP_1490-99.2;hHSFY2:NP_714927.1)、挪威鼠(NP_00101-2132.1)、家短尾負(fù)鼠(ACT35160)、恒河猴(ACL-51668.1)、家貓(NP_001035212.1)等的 HSFY 蛋白的序列用 Clustal W[11]進(jìn)行多序列比對(duì)[6],用MEGA5軟件構(gòu)建系統(tǒng)進(jìn)化樹(shù)。在UCSC基因組瀏覽器(http://genome.ucsc.edu/)的普通牛基因組中,以HSFY引物對(duì)搜索,選取位于Y染色體上的HSFY相似序列。GENSCAN預(yù)測(cè)外顯子區(qū)域,condon W(http://mobyle.pasteur.fr/cgi-bin/portal.py?#forms::codonw)分析其密碼子偏好性和有效密碼子數(shù)等。用 SWISS-MODEL(http://swissmodel.expasy.org/)同源建模構(gòu)建HSFY蛋白三維結(jié)構(gòu)。
2 結(jié)果
2.1 PCR擴(kuò)增結(jié)果
經(jīng)PCR擴(kuò)增,電泳檢測(cè)為一清晰條帶(圖1)。結(jié)合普通牛HSFY2序列(NC_016145.1),通過(guò)DNA分子量標(biāo)準(zhǔn)和測(cè)序結(jié)果分析得知,牦牛HSFY基因PCR產(chǎn)物大小為2000bp左右,與預(yù)期結(jié)果一致。

圖1 牦牛HSFY基因的PCR擴(kuò)增電泳圖
2.2 牦牛HSFY基因的核苷酸序列及其比對(duì)
本研究得到的牦牛HSFY基因由2個(gè)外顯子和1個(gè)內(nèi)含子組成,與普通牛基因組中的相應(yīng)序列進(jìn)行比對(duì),其一致性高達(dá)99.54%。第一外顯子未出現(xiàn)插入/缺失,但在1012-1020bp(Ⅰ)處有AAAGAAGA/TG等9個(gè)核苷酸缺失,位于內(nèi)含子內(nèi);在1071-1073bp(Ⅱ)、1249-1251bp(Ⅲ)處分別有AAG、CCA/C等3個(gè)核苷酸缺失,位于第二外顯子內(nèi),分別導(dǎo)致賴氨酸和組氨酸的缺失;在1 708-1 710bp(Ⅳ)處有CAA 3個(gè)核苷酸缺失,位于3'-URT。在普通牛HSFY基因的84條序列中Ⅰ、Ⅱ、Ⅲ、Ⅳ插入/缺失分別有29、3、2、1次,未發(fā)現(xiàn)Ⅱ、Ⅲ同時(shí)缺失的拷貝,但Ⅱ、Ⅲ缺失均伴有Ⅰ缺失。Ⅰ、Ⅱ、Ⅲ、Ⅳ插入/缺失的序列與其相鄰序列極為相似(圖2)。

圖2 兩條牦牛HSFY基因序列與普通牛部分相應(yīng)序列的比對(duì)
HSFY基因在許多物種中為多拷貝基因,但在物種內(nèi)序列的相似性極高,人的兩個(gè)功能拷貝的氨基酸序列相似性達(dá)100%。本研究得到的牦牛HSFY序列1僅有Ⅲ缺失,且預(yù)測(cè)的基因剪切方式也與普通牛中具有功能的HSFY基因剪切方式一致。相對(duì)序列2含有Ⅰ、Ⅲ、Ⅳ3處缺失,序列1預(yù)測(cè)的氨基酸序列更能反映牦牛HSFY蛋白質(zhì)信息。其與人、恒河猴、普通牛、瘤牛、家貓、挪威鼠、家短尾負(fù)鼠的HSFY蛋白的氨基酸序列進(jìn)行多序列比對(duì),構(gòu)建系統(tǒng)進(jìn)化樹(shù)(圖3)。牦牛與普通牛、瘤牛首先聚為一類(lèi),再相繼與家貓、家短尾負(fù)鼠、人等聚類(lèi),聚類(lèi)結(jié)果與動(dòng)物學(xué)分類(lèi)結(jié)果基本一致。
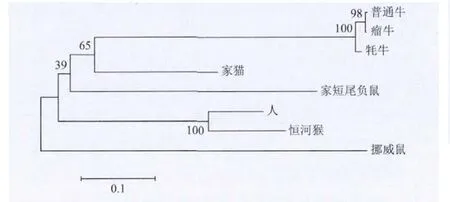
圖3 牦牛與普通牛等物種HSFY的系統(tǒng)進(jìn)化關(guān)系
2.3 牦牛HSFY蛋白質(zhì)的三維結(jié)構(gòu)
以牦牛序列1的氨基酸序列為基礎(chǔ)序列,研究HSFYⅡ、Ⅲ插入/缺失與蛋白的三維結(jié)構(gòu)變化情況。結(jié)果(圖4)表明,Ⅱ與Ⅲ的單獨(dú)缺失與Ⅱ和Ⅲ均缺失的三維結(jié)構(gòu)基本一致,但與Ⅱ、Ⅲ均未缺失的結(jié)構(gòu)差異較大。Ⅱ、Ⅲ插入/缺失對(duì)應(yīng)的帶正電荷的賴氨酸、組氨酸比較傾向于位于蛋白表面,通過(guò)靜電作用等維持HSFY蛋白的立體結(jié)構(gòu),Ⅱ、Ⅲ任一缺失均使HSFY蛋白的立體結(jié)構(gòu)塌陷。Ⅱ、Ⅲ插入/缺失可能會(huì)使HSFY蛋白的功能發(fā)生變化進(jìn)而影響精子發(fā)生。
2.4 牦牛HSFY基因的密碼子偏好性
以牦牛HSFY序列1作為密碼子偏好性分析,GENSCAN預(yù)測(cè)得到長(zhǎng)1131bp的蛋白編碼區(qū)。A、T、C和G分別占33.24%、29.27%、20.34%和17.15%,A3s、T3s、G3s和C3s分別為30.77%、37.93%、15.91%和15.38%,其中G+C、G3s+C3s含量分別為37.19%和31.30%。有效密碼子數(shù)為46.82。可見(jiàn),HSFY基因偏好使用A、T和以A、T結(jié)尾的密碼子,UUU、UUG、CUU、AUU、GUG、UAU、CAU、CAA、GAA、GAU、UCU、UCA、CCU、CCA、ACU、ACA、GCU、GCA、UGU、AGA、AGG、GGC和 GGA等 23個(gè)密碼子RSCU(相對(duì)密碼子使用度)均大于1,為牦牛HSFY偏好性密碼子;Asn、Lys、Arg、Ser 4個(gè)氨基酸沒(méi)有密碼子偏好現(xiàn)象;除編碼Leu、Val、Arg、Gly的UUG、GUG、AGG、GGC外,避免密碼子后兩位以G或C結(jié)尾,尤其以CG結(jié)尾如CCG、ACG、GCG的 RSCU為 0(表1)。

圖4 牦牛HSFY蛋白質(zhì)的Ⅱ、Ⅲ插入/缺失的三維結(jié)構(gòu)
3 討論
3.1 關(guān)于牦牛HSFY基因的拷貝數(shù)
在人體中,研究發(fā)現(xiàn)Yq11區(qū)存在與精子發(fā)生相關(guān)的基因,這些基因或基因家族又稱為無(wú)精子癥因子(azoospermia facter,AZF),目前已經(jīng)確定AZF由相互疊加的4個(gè)區(qū)域組成,即AZFa、AZFb、AZFc、AZFd[12,13]。HSFY 是位于 AZFb 區(qū)無(wú)精子癥因子候選因子之一。在許多物種中發(fā)現(xiàn)是由多拷貝組成的基因,人有6個(gè)拷貝,家貓有8個(gè)功能拷貝,普通牛約有73個(gè)拷貝[3,6,7]。以往的研究表明,牦牛與普通牛具有較近的親緣關(guān)系,在許多基因組成和基因結(jié)構(gòu)上都有相似性,本研究得到的2條牦牛HSFY基因序列雖在核苷酸組成上有一些差異,但相似性很高。因此認(rèn)為牦牛的HSFY基因是一個(gè)多拷貝基因。

表1 牦牛HSFY序列1的RSCU
3.2 關(guān)于牦牛HSFY基因的結(jié)構(gòu)與遺傳變異
Hamilton等[6]研究普通牛HSFY基因的拷貝數(shù)在67-84范圍波動(dòng),基因表達(dá)和56天NRR系數(shù)(反映受精比率)均無(wú)顯著性差異。與Shaw-Smith[14]、Inoue[15]、Lupski[16]等研究表明的基因微量刪除和重復(fù)嚴(yán)重影響生物學(xué)功能現(xiàn)象不一致,推測(cè)是由多拷貝和基因調(diào)控序列區(qū)域變化兩種調(diào)節(jié)機(jī)制確保進(jìn)行精子發(fā)生。該機(jī)制可能受到嚴(yán)格的自然選擇。本研究得到2條牦牛的HSFY序列,并搜索得到普通牛的HSFY序列84條,與Hamilton[6]研究結(jié)果基本一致,表明引物結(jié)合處比較保守,可以用于檢測(cè)HSFY基因缺失狀態(tài)。比對(duì)發(fā)現(xiàn)牦牛、普通牛有4處插入/缺失,其序列均分別與其相鄰序列極為相似,可能由染色體內(nèi)同源重組導(dǎo)致缺失的發(fā)生[17]。Ⅰ、Ⅳ缺失雖然不引起蛋白結(jié)構(gòu)的變化,但估計(jì)起轉(zhuǎn)錄調(diào)控作用,以便精細(xì)調(diào)節(jié)細(xì)胞中HSFY蛋白濃度變化彌補(bǔ)個(gè)別基因因突變或缺失造成的功能缺失。Ⅱ、Ⅲ插入/缺失對(duì)應(yīng)的賴氨酸和組氨酸的零深度相對(duì)傾向因子分別為30.850、15.312[18],使均帶正電荷的賴氨酸、組氨酸傾向于位于蛋白表面,HSFY蛋白的立體結(jié)構(gòu)塌陷。Ⅱ、Ⅲ插入/缺失可能會(huì)使HSFY蛋白的功能發(fā)生變化進(jìn)而影響精子發(fā)生。普通牛中Ⅱ和Ⅲ缺失均伴有Ⅰ缺失,很可能Ⅰ插入/缺失參與該基因的精確表達(dá)調(diào)控,但是使得該基因更易發(fā)生缺失突變。當(dāng)然作者的這些推測(cè)還有待于進(jìn)一步的生物試驗(yàn)證實(shí)。
3.3 關(guān)于牦牛HSFY蛋白的結(jié)構(gòu)與功能
熱休克蛋白(heat shock protein,HSP)是指所有生物細(xì)胞在應(yīng)激原刺激后,發(fā)生熱休克反應(yīng)時(shí)產(chǎn)生的一類(lèi)細(xì)胞伴侶蛋白的總稱。該類(lèi)蛋白在細(xì)胞生命過(guò)程中可參與蛋白的折疊、裝配及運(yùn)輸?shù)然顒?dòng),對(duì)調(diào)控細(xì)胞的生長(zhǎng)、存活及分化起著重要作用。在哺乳動(dòng)物中,HSP家族參與精子發(fā)生、精子獲能及受精等一系列活動(dòng),與雄性生殖過(guò)程密切相關(guān)[19]。研究表明,人和貓Y染色體上多拷貝基因都在睪丸組織中表達(dá)并行使功能[8,20-22],則進(jìn)一步推測(cè)牦牛HSFY在睪丸中表達(dá)和行使生物學(xué)功能。HSF成員HSF1、HSF2在精子發(fā)生中也起到了重要作用[23,24]。Ahn 等[25]報(bào)道,HSFY 和其它 HSFs(HSF1、HSF2、HSF4)的結(jié)構(gòu)不同,后者的中心螺旋型是由α-螺旋2和3組成,α-螺旋3是識(shí)別的DNA的螺旋結(jié)構(gòu),然而HSFY有類(lèi)似HSFs的DNA結(jié)合區(qū),但沒(méi)有α-螺旋2和3。推測(cè)在精子發(fā)生活動(dòng)中,HSFY可能具有與其它HSFs不同的調(diào)控HSP表達(dá)途徑。
蛋白質(zhì)序列變化[26]與其生物學(xué)功能變化密切相關(guān)。本研究中,牦牛、普通牛HSFY基因未發(fā)現(xiàn)Ⅱ、Ⅲ同時(shí)缺失,且Ⅱ、Ⅲ任意一處缺失與均缺失的三維結(jié)構(gòu)基本一致且有5個(gè)序列,占5.95%,可能與精子發(fā)生受阻相關(guān)。因缺失的比例較小和調(diào)控其它正常基因,確保了生物體精子發(fā)生正常。可通過(guò)Ⅱ、Ⅲ兩端的保守序列設(shè)計(jì)引物進(jìn)一步研究其數(shù)量變化與生物活動(dòng)變化之間的規(guī)律。值得一提的是基因密碼子的使用與基因表達(dá)的生理功能有密切聯(lián)系[27,28]。其不僅調(diào)控基因表達(dá)[29],而且與翻譯的準(zhǔn)確性和效率有關(guān)[30]。本研究以牦牛序列1預(yù)測(cè)的編碼區(qū)偏好使用A、T和以A、T結(jié)尾的密碼子。考慮到該基因在小鼠精子發(fā)生過(guò)程中有瞬時(shí)高表達(dá)現(xiàn)象,估計(jì)牦牛HSFY翻譯比較低的準(zhǔn)確度,更有利于瞬時(shí)高表達(dá)行使功能。但其生物學(xué)機(jī)制需要進(jìn)一步研究。
4 結(jié)論
牦牛HSFY基因由2個(gè)外顯子和1個(gè)內(nèi)含子組成,是在睪丸中表達(dá)的多拷貝基因,與普通牛基因組中的相應(yīng)序列的一致性較高,屬于同源性的基因。
牦牛、普通牛的該基因具有4處插入/缺失,且序列與其相鄰序列極為相似。牦牛HSFY基因編碼的蛋白質(zhì)偏好使用以A或T結(jié)尾的密碼子。
[1] Pirkkala L, Nykanen P, Sistonen L.Roles of theheat shock transcription factors in regulation of theheat shock response and beyond [J].FASEB J, 2001, 15(7):1118-1131.
[2] Shinka T, Sato Y, Chen G, et al.Molecular characterization ofheat shock-like factor encoded on thehuman Y chromosome, and implications formale infertility [J].Biol Reprod, 2004, 71(1):297-306.
[3] Tessari A, Salata E, Ferlin A, et al.Characterization of HSFY, a novel AZFb gene on the Y chromosome with a possible role inhuman spermatogenesis [J].Mol Hum Reprod, 2004, 10(4):253-258.
[4] Vinci G, Raicu F, Popa L, et al.A deletion of a novelheat shock gene on the Y chromosome associated with azoospermia [J].Mol Hum Reprod, 2005, 11(4):295-298.
[5] Ferlin A, Moro E, Rossi A, et al.Thehuman Y chromosomes azoospermia factor b(AZFb)region:squence, structure and deletion analysis infertilemen [J].J Med Genet, 2003, 40:13-24.
[6] Hamilton CK, Revay T, Domander R, et al.A large expansion of the HSFY gene family in cattle shows dispersion across Yq and Testisspecific expression [J].PLoS One, 2011, 6(3):e17790.
[7] Pearks Wilkerson AJ, Raudsepp T, Graves T, et al.Gene discovery and comparative analysis of X-degenerate genes from the domestic cat Y chromosome [J].Genomics, 2008, 92(5):329-338.
[8] Skaletsky H, Kuroda-Kawaguchi T, Minx PJ, et al.Themale-specific region of thehuman Y chromosome is amosaic of discrete sequence classes [J].Nature, 2003, 423:825-837.
[9] Sato Y, Yoshida K, Shinka T, et al.Altered expression pattern ofheat shock transcription factor, Y chromosome(HSFY)may be related to altered differentiation of spermatogenic cells in testes with deteriorated spermatogenesis[J].Fertility and Sterility, 2006, 86(3):612-618.
[10] Kinoshita K, Shinka T, Sato Y, et al.Expression analysis of amouse orthologue of HSFY, a candidate for the azoospermic factor on thehuman Y chromosome [J].J Med Invest, 2006, 53:117-122.
[11] Thompson JD, Higgins DG, Gibson TJ.CLUSTAL W:Improving the sensitivity of progressivemultiple sequence alignment through sequence weighting, position-specific gap penalties and weightmatrix choice [J].Nucleic Acids Res, 1994, 22:4673-4680.
[12] Vogt PH.Human chromosome deletions in Yq11, AZF candidate genes andmale infertility:History and update [J].Mol Hum Reprod, 1998, 4(8):739-744.
[13] Kent-First M, Muallem A, Shultz J, et al.Defining regions of the Y chromosome responsible formale infertility and identification of a fourth AZF region(AZFd)by Y-chromosomemicrodeletion detection [J].Mol Reprod Dev, 1999, 53(1):27-41.
[14] Shaw-Smith C, Redon R, Rickman L, et al.Microarray based comparative genomichybridisation(array-CGH)detects submicroscopic chromosomal deletions and duplications in patients with learning disability/mental retardation and dysmorphic features[J].J Med Genet, 2004, 41(4):241-248.
[15] Inoue K, Lupski JR.Molecularmechanisms for genomic disorders[J].Annu Rev Genomics Hum Genet, 2002, 3:199-242.
[16] Lupski JR, Stankiewicz P.Genomic disorders:molecularmechanisms for rearrangements and conveyed phenotypes [J].PLoS Genet, 2005, 1(6):e49.
[17] Vogt PH.Molecular genetic ofhumanmale infertility:from genes to new therapeutic perspectives [J].Curr Pharm Des, 2004, 10(5):471-500.
[18] 沈世錳, 胡剛, 張華.氨基酸在蛋白質(zhì)結(jié)構(gòu)中的深度傾向性因子[J].生物數(shù)學(xué)學(xué)報(bào), 2007, 22(2):305-310.
[19] 朱曄敏, 丁之德.雄性生殖中熱休克蛋白作用的研究進(jìn)展[J].國(guó)際生殖健康/計(jì)劃生育雜志, 2010, 29(1):18-21.
[20] Murphy WJ, Pearks Wilkerson AJ, Raudsepp T, et al.Novel gene acquisition on carnivore Y chromosomes [J].PLoS Genet, 2006, 2:e43.
[21] Lahn BT, Pearson NM, Jegalian K.Thehuman Y chromosome, in the light of evolution [J].Nat Rev Genet, 2001, 2:207-216.
[22] Lahn BT, Page DC.Functional coherence of thehuman Y chromosome [J].Science, 1997, 278(5338):675-680.
[23] Nakai A, Suzuki M, Tanabe M.Arrest of spermatogenesis inmice expressing an activeheat shock transcription factor [J].EMBO J,2000, 19:1545-1554.
[24] Kallio M, Chang M, Manuel M, et al.Brain abnormalities defectivemeiotic chromosome synapsis and female subfertility in HSF2 nullmice [J].EMBO J, 2002, 21:2591-2601.
[25] Ahn SG, Liu PCC, Klyachko K, et al.The loop domain ofh eat shock transcription factor 1dictates DNA-binding specificity and responses toheat stress [J].Genes Dev, 2001, 15:2134-2145.
[26] Redon R, Ishikawa S, Fitch KR, et al.Global variation in copy number in thehuman genome [J].Nature, 2006, 444(7118):444-454.
[27] Karlin S, Campbell AM, Mrázek J.Comparative DNA analysis across diverse genomes [J].Annu Rev Genet, 1998, 32:185-225.
[28] Kennedy SP, Ng WV, Salzberg SL.Understanding the adaptation of Halobacterium species NRC-1 to its extreme environment through computational analysis of its genome sequence [J].Genome Res,2001, 11(10):164-165.
[29] Carlini DB, Stephan W.In vivo introduction of unpreferred synonymous codons into the Drosophila Adh gene results in reduced levels of ADH protein [J].Genetics, 2003, 163(1):239-243.
[30] Sharp PM, Matassi G.Codon usage and genome evolution [J].Curr Opin Genet Dev, 1994, 4(6):851-860.
猜你喜歡
中華詩(shī)詞(2022年6期)2022-12-31 06:41:24
哲學(xué)評(píng)論(2021年2期)2021-08-22 01:53:34
中華詩(shī)詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中國(guó)科技論壇(2017年7期)2017-07-25 08:49:53
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國(guó)際漢語(yǔ)學(xué)報(bào)(2016年1期)2017-01-20 08:21:20
影視與戲劇評(píng)論(2016年0期)2016-11-23 05:26:01
現(xiàn)代企業(yè)(2015年9期)2015-02-28 18:56:50
中國(guó)中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年22期)2014-03-01 04:32:55
