水分脅迫對不同品種小麥幼芽抗氧化酶活性的影響
2013-09-15 08:04:52張述義劉玲玲
山西農業科學 2013年2期
關鍵詞:植物
張述義,劉玲玲
(1.山西省農業科學院旱地農業研究中心,山西太原030006;2.山西省農業科學院小麥研究所,山西臨汾041000)
植物體在水分脅迫逆境條件下,質膜膜脂的物理狀態改變可能是植物感受滲透脅迫的原初響應[1]。植物在逆境下通常伴隨大量活性氧自由基的產生,引起細胞中產生活性氧自由基與活性氧自由基的清除系統失衡,如果不及時清除將會造成細胞膜和一些大分子物質的破壞[2],膜結構中含有的不飽和脂肪酸的類脂易受活性氧自由基攻擊,導致膜脂過氧化作用發生,脂質過氧化產物MDA含量增加,使質膜的選擇透性喪失,電解質與一些小分子有機物質外滲,最終導致植物傷害發生[3]。植物體內的抗氧化系統是植物細胞對抗氧化脅迫的關鍵因素,其中,超氧化物歧化酶( SOD)、過氧化氫酶( CAT)、過氧化物酶( POD)是抗氧化酶系統中3種重要的保護酶,是生物體內重要的活性氧自由基清除劑,可以清除機體內多余的活性氧自由基,解除活性氧自由基對生物體的毒性[4],維持植物體內活性氧自由基產生和淬滅的動態平衡,防止膜脂過氧化。
干旱是影響小麥播種出苗的主要因素,尤其在小麥幼芽期,干旱逆境下成苗的關鍵不在于萌發,而主要在于萌發后的幼芽伸長[5-6]。為了解不同肥水類型小麥品種幼芽抗氧化酶系統對水分脅迫逆境的反應,本研究選用晉麥47號(冬性半冬性、旱地品種)和舜麥1718(冬性半冬性、水地品種)2個不同肥水類型的小麥品種,在發芽期進行水分脅迫處理,對幼芽中的抗氧化酶SOD,POD,CAT活性以及MDA含量和蛋白質含量進行了分析,以期為旱地栽培中選用高產小麥品種提供理論依據。
1 材料和方法
1.1 試驗材料
供試材料晉麥47號、舜麥1718小麥品種,分別由山西省農業科學院小麥研究所和棉花研究所提供。
1.2 材料培養
溫湯浸泡小麥種子30 min,之后瀝出種子,(25±1)℃清水浸泡24 h后,再將種子瀝出,挑選萌動勢一致的種子,擺入培養皿(Ф10 cm)中,加入混合溶液( 0.1 mmol/LKCl,0.1 mmol/LCaCl2,0.1 mmol/L MgCl2,0.5 mmol/L NaCl,0.2 mmol/L Na20.3mmol/LMES),( 25±1)℃繼續培養 5d。
1.3 試驗設計
將培養5 d的小麥培養皿內水分吸凈,加入0(對照),19.2%(水分脅迫處理)聚乙二醇(PEG6000)溶液進行處理,24 h后取樣,測定SOD,POD,CAT酶活性和MDA及蛋白質含量,重復3次。
1.4 測定項目及方法
SOD活性測定,以反應抑制氮藍四唑(NBT)光化還原50%的酶量為一個酶活單位,U/(g·h)[7];POD活性采用愈創木酚法測定,以每分鐘內吸光度(變化0.1為一個酶活單位,U/(g·min)[8];CAT活性采用紫外分光光度法測定,以1 min內A240減少0.1的酶量為一個酶活單位,U/(g·min)[9];MDA含量采用硫代巴比妥酸法測定[7];蛋白質含量采用考馬斯亮藍染色法測定[9]。
1.5 數據分析
采用Excel對試驗數據進行統計分析。
2 結果與分析
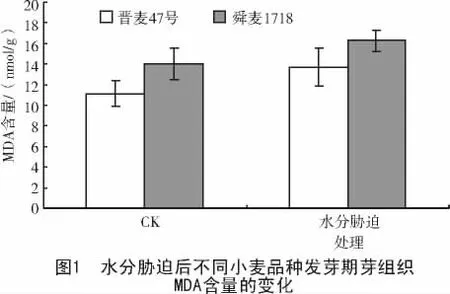
2.1 水分脅迫后不同小麥品種幼芽MDA含量的變化
由于水分脅迫處理可以引起植物體細胞內有害自由基增多,引發膜中不飽和脂肪酸過氧化反應發生。MDA是膜脂過氧化作用的主要產物,能與酶蛋白發生鏈式反應聚合,使膜系統變性,造成膜脂流動性降低,影響膜結合酶的功能,膜脂過氧化產物MDA含量的增加,會造成膜傷害,則MDA含量反映膜脂質過氧化程度[10-11]。
試驗結果(圖1)表明,在正常供水的對照中,晉麥47號幼芽中MDA含量低于舜麥1718;水分脅迫處理24 h后,晉麥47號和舜麥1718的MDA含量均升高,與對照相比,晉麥47號的MDA含量增加了23.26%,舜麥1718增加了15.9%。說明舜麥1718幼芽中的過氧化代謝產物積累相對少,對膜損傷程度較小。
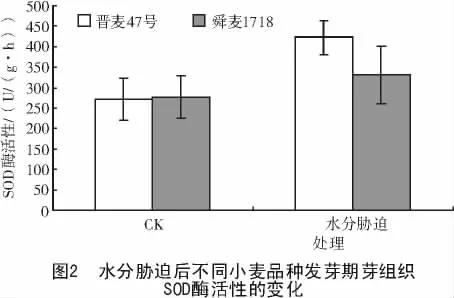
2.2 水分脅迫后不同小麥品種幼芽SOD酶活性的變化
SOD是植物體內一個關鍵的抗氧化酶,可通過Haber-weiss反應清除植物體內多余的超氧根陰離子,將2個超氧自由基發生歧化反應形成水和氧。從圖2可以看出,對照的幼芽中晉麥47號與舜麥1718的SOD酶活性相近,分別為271.25,277.16 U/(g·h)。水分脅迫處理24 h后,2個小麥品種的SOD酶活性均上升,晉麥47號上升幅度大于舜麥1718,二者的SOD酶活性分別增加了55.83%,19.58%,可見,在水分脅迫下,晉麥47號幼芽以提高SOD酶活性淬滅超氧陰離子的能力大于舜麥1718。
2.3 水分脅迫后不同小麥品種幼芽CAT酶活性的變化
過氧化氫(H2O2)是植物生長過程中的代謝產物,逆境會導致過量的過氧化氫產生,對機體造成損害。CAT主要功能是分解植物組織中氧代謝中產生的H2O2以及有機過氧化物 ROOH,快速地轉化為其他無害較小的物質,可以改善質膜氧化損害程度,CAT酶活力越高,H2O2以及ROOH增加的比例越小。從圖3可以看出,對照的舜麥1718幼芽中CAT酶活性高于晉麥47號。經過24 h水分脅迫處理后,2個小麥品種的CAT酶活性均提高。晉麥47號的CAT酶活性比對照增加了163.84%,舜麥1718的CAT酶活性比對照增加了11.69%,可見,晉麥47號幼芽中的過氧化氫酶活性對水分脅迫反應非常敏感。

2.4 水分脅迫后不同小麥品種幼芽POD酶活性的變化
POD在植物體內的主要作用也是清除H2O2,并將其分解為O2和H2O,其活性升高可以減輕活性氧對細胞膜的傷害。由圖4可知,晉麥47號幼芽POD酶活性高于舜麥1718。水分脅迫處理24 h后,晉麥47號的POD酶活性比對照增加92.52%,舜麥1718比對照增加59.25%。

2.5 水分脅迫后不同小麥品種幼芽蛋白質含量的變化
可溶性蛋白質是植物滲透調節物質之一,在維持生物膜結構功能穩定性、調節離子運輸、抑制蛋白酶活性等方面起重要作用。很多與植物耐旱性相關的逆境誘導蛋白可能在不同小麥品種之間存在較一致的調節機制。由圖5可知,舜麥1718幼芽中蛋白質含量高于晉麥47號,水分脅迫處理24 h后,晉麥47號蛋白質含量與對照相比增加118.84%,舜麥1718增加了23.39%,與酶活性增加趨勢一致。

2.6 水分脅迫后不同小麥品種幼芽抗氧化酶活性總體的變化
從圖6可以看出,小麥發芽期,水分脅迫處理24 h后,晉麥47號幼芽中3個抗氧化酶活性增加百分率的總和大于舜麥1718,晉麥47號的抗氧化酶活性對水分脅迫敏感。晉麥47號以蛋白質為主要滲透調節物質,以增強抗氧化酶活性,來抵御水分脅迫的危害。

3 結論與討論
晉麥47號是一個抗旱、高產的優良旱地冬小麥品種,舜麥1718是一個高產優良的水地冬小麥品種[12-15],晉麥47號幼芽在水分脅迫處理24 h后,3個抗氧化酶活性增強,表現為清除體內活性氧自由基能力加強。SOD在清除超氧化物陰離子自由基的過程中,會產生H2O2,對細胞造成傷害。過氧化氫酶(CAT)具有清除過量H2O2的作用,與SOD和POD一起保護膜系統免受自由基的傷害。舜麥1718幼芽中的3個抗氧化酶對水分脅迫反應沒有晉麥47號那么敏感,其抗氧化酶活性增強水平較低,活性氧代謝活動相對平穩,可能是由于自身具有其他抑制或清除過氧化物積累的能力,但這還有待于進一步研究。
在水分脅迫下,不同小麥品種的蛋白質含量都有所增加,這可能是由于幼芽蛋白質組分發生了變化,有新蛋白亞基產生,這些蛋白可能是一些結構蛋白,也可能是一些滲透調節物質或酶,會在植物受到傷害時起保護作用[16]。在水分脅迫下,晉麥47號蛋白質含量增長最大,在逆境誘導蛋白的表達方面優于舜麥1718。
不同小麥品種幼芽,在水分脅迫下通過不同的生理代謝過程和代謝水平,維持體內活性氧產生和淬滅的動態平衡。旱地品種晉麥47號主要是依靠提高SOD,POD,CAT酶活性和蛋白質含量來抵御干旱的危害;水地品種舜麥1718雖然抗氧化酶活性變化幅度小,但從膜脂過氧化狀態分析,有研究顯示,抗旱性較強的品種MDA含量增加幅度低于抗旱性弱的品種[17],水分脅迫下舜麥1718的MDA含量增加小于晉麥47號,由于MDA含量變化率小,質膜的損傷程度較輕,表明水地小麥品種舜麥1718可能具有適合旱地栽培的潛力,植物受到干旱脅迫后,可通過啟動體內的一系列反應來抵御干旱[18],2個品種抵御水分脅迫危害可以通過不同途徑來實現,其生理代謝特點還需要從多方面進一步深入探討。
[1]邱全勝.滲透脅迫對小麥根質膜膜脂物理狀態的影響[J].植物學報,1999,41( 2):161-165.
[2]高銀.植物抗逆機制與基因工程研究進展[J].內蒙古農業科技,2007( 5):75-78.
[3]陳少裕.膜脂過氧化與植物逆境脅迫 [J].植物通報,1989,6( 4):211-217.
[4]賀杰,王偉,胡海燕.小麥種子活力與其保護酶活性關系的研究[J].天津農業科學,2012,18( 2):17-19.
[5]山侖,郭禮坤.逆境成苗生態生理研究Ⅰ.春播谷類作物成苗期間的抗旱性及其需水條件 [J].作物學報,1984,10(4):257-263.
[6]楊美紅,賈俊仙,李國鋒.水分脅迫對不同耐旱性品種小麥幼芽、幼苗的保護酶和淀粉酶活性的影響 [J].山西農業科學,2001,29( 1):45-48.
[7]中國科學院上海植物生理研究所.現代植物生理學實驗指南[M].北京:科學出版社,1999.
[8]龔富生,張嘉寶.植物生理學實驗[M].北京:氣象出版社,1995.
[9]鄒琦.植物生理學實驗指導[M].北京:中國農業出版社,2000.
[10]山侖,陳培元.旱地農業生理生態基礎[M].北京:科學出版社,1998:1-18.
[11]王寶山.生物自由基與植物膜傷害 [J].植物生理學通訊,1988( 2) :2-16.
[12]孫來虎,李秀絨,柴永峰,等.晉麥47號產量結構特點與高產栽培技術[J].耕作與栽培,2003( 5):48-49.
[13]董孟雄,李秀絨,柴永峰,等.旱地小麥新品種:晉麥47號[J].麥類作物學報,2001,21( 1):98.
[14]張運校,樊立強,劉彥軍,等.國審優質小麥新品種舜麥1718高產栽培技術[J].種子科技,2012( 3):28-29.
[15]潘幸來,史引紅,王永杰,等.舜麥1718小麥新品種選育報告[J].小麥研究,2007,28( 4):26-36.
[16]戴明,鄧西平,楊淑慎,等.水分脅迫對不同基因型小麥幼芽蛋白質表達和某些生理特性的影響 [J].應用生態學報,2009,20( 9):2149-2156.
[17]張文英,智慧,柳斌輝.谷子孕穗期一些生理性狀與品種抗旱性的關系[J].華北農學報,2011,26( 3):128-133.
[18]榮少英,郭蜀光,張彤.干旱脅迫對甜高粱幼苗滲透調節物質的影響[J].河南農業科學,2011,40( 4):56-59.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
