大白菜雄性不育系MS200712 的研究簡報
2014-02-23 10:50:06
中國蔬菜 2014年8期
(北京市農林科學院蔬菜研究中心,北京 100097)
大白菜雄性不育源主要有細胞核雄性不育和細胞質雄性不育。細胞核雄性不育是張書芳等(1990)在大白菜品種萬泉青幫采種田中發現的;細胞質雄性不育則由原生質體融合、雜交和多代回交等途徑,轉育蘿卜Ogu CMS、甘藍型油菜Pol CMS 和新型甘藍型油菜CMS96 而獲得的大白菜Ogu CMS(Williams,1981;Health et al.,1994)、Pol CMS(柯桂蘭 等,1992)和CMS96(張德雙 等,2002)細胞質雄性不育系。可見,大白菜細胞質雄性不育系的主要來源是轉育已有的十字花科細胞質雄性不育系,目前還未見直接利用蕓薹屬種間雜交的方法創制新型大白菜雄性不育系的報道。
為了拓寬大白菜雄性不育系資源、類型以及研究雄性不育發生的機制,本試驗以蕓薹屬種間雜交的方法創制和獲得新型大白菜雄性不育系,為今后在大白菜育種中更廣范地應用雄性不育系奠定 基礎。
1 材料與方法
1.1 材料
供試蕓薹屬異源六倍體(AABBCC,n=27)擁有蕓薹A、黑芥B、甘藍C 的全部染色體,主要性狀表現為:株型高大,心葉略帶紫色,葉片綠色、有缺刻、覆蠟粉,莖部粗大、略帶紫色、覆蠟粉,腋芽萌發能力強,花瓣黃色,花藥和柱頭正常,結實性好,種莢細長,種子較大、黑褐色(張德雙 等,2009),該異源六倍體來源于北京市農林科學院蔬菜研究中心大白菜課題組;北京新3號的父本832172 也來源于本中心大白菜課題;大白菜品種金春購自遼寧東亞國際種苗有限公司,義和5號購自青島膠州市膠白蔬菜育種中心。
1.2 方法
1.2.1 雜交方法 植株盛花期時,在父本花序上預先套硫酸紙袋,選取母本生長旺盛、花蕾多的主枝做雜交,徹底摘除用于雜交的全部花蕾的6個雄蕊,然后涂抹父本的花粉到母本柱頭上,套上硫酸紙袋,并用曲別針扎口,以防止被昆蟲攜帶的花粉污染;第2 天上午再涂抹父本的花粉到母本柱頭上,然后扎好袋口。雜交完成后應注意澆水,及時修剪多余枝條,撤掉硫酸紙袋。在母本上收獲的種子即為雜交一代。
1.2.2 試驗步驟 試驗在本中心日光溫室內完成。首先,以蕓薹屬異源六倍體為母本,以大白菜品種金春為父本進行雜交,獲得F1(AABBCC×AA);以大白菜品種義和5號為母本,以F1(AABBCC×AA)為父本再次進行雜交。在多親本雜交后代中即可篩選到大白菜雄性不育系MS200712(圖1)。
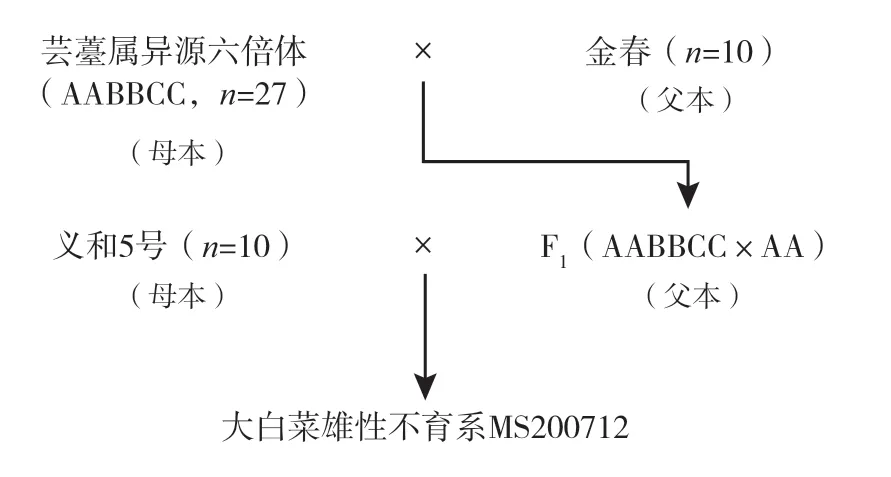
圖1 獲得大白菜雄性不育系MS200712 的技術路線
最后,用義和5號、北京新3號的父本832172 與大白菜雄性不育系MS200712 進行回交轉育,并鑒定轉育后代的花粉育性。
2 結果與分析
2.1 雜交一代F1(AABBCC×AA)的獲得
在金春的花序上預先套上硫酸紙袋,徹底摘除蕓薹屬異源六倍體花序中用于雜交的全部花蕾的6個雄蕊,然后涂抹金春的花粉,套上硫酸紙袋,并扎好袋口;第2 天上午再重復授粉1次。在蕓薹屬異源六倍體上收獲的種子即為F1(AABBCC× AA)。
在人工授粉獲得F1(AABBCC×AA)的過程中,為了防止因雄蕊去除不徹底而混雜自交種子,在剝蕾時只注重質量,不追求數量,AABBCC 異源六倍體的花藥被徹底去除。采用2種保障措施:①用鑷子剝蕾時,舍棄掉那些不容易操作的小花蕾,每一個花序的授粉花蕾數不要過多,以5~10個為宜;②在第2 天重復授粉時,再次確認是否殘留母本的花藥。由圖2可見,F1葉片具有葉翼、長倒卵形,抽薹、現蕾期較晚;而AABBCC 異源六倍體葉片沒有葉翼、近圓形,抽薹、現蕾期較早(張德雙 等,2009)。可見,F1與母本的性狀已經存在較明顯差異,特別是F1葉片具有葉翼,標志著F1基因組中存在父本金春AA基因組。SRAP 標記的電泳檢測結果進一步證明F1為真正的雜交種(待 發表)。

圖2 F1(AABBCC×AA)的植株
2.2 大白菜雄性不育系MS200712 的獲得和鑒定
在F1(AABBCC×AA)花序上預先套上硫酸紙袋,徹底摘除義和5號花序中用于雜交的全部花蕾的6個雄蕊,然后涂抹F1(AABBCC×AA)的花粉,套上硫酸紙袋,并扎好袋口;第2 天上午再重復授粉1次。在義和5號上收獲的種子即包含大白菜雄性不育系MS200712。
翌年春季繼續播種MS200712。在盛花期觀察MS200712 的花藥,結果花藥完全退化、敗育徹底,表現為雄性不育,即獲得了人工創造的大白菜雄性不育系MS200712。該雄性不育系葉片深綠色、長橢圓形,葉緣多鋸齒,花瓣黃色,花藥淺黃色、退化、敗育徹底,花藥長短正常,柱頭正常,蜜腺發達,可正常吸引昆蟲授粉(圖3~5)。

圖3 大白菜雄性不育系MS200712 的植株和花序
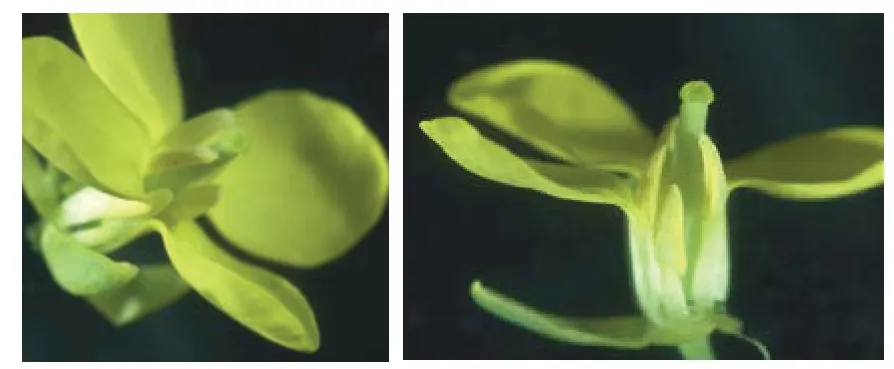
圖4 大白菜雄性不育系MS200712 的花器官

圖5 大白菜雄性不育系MS200712 的花藥和蜜腺
2.3 大白菜雄性不育系MS20071208-840 和MS20071208-916 的獲得
以MS200712 雄性不育系為母本,分別與義和5號、832172 進行雜交,在MS200712 不育系上 收獲的種子即為MS20071208-840和MS20071208-916。分別播種MS20071208-840和MS20071208-916,在盛花期觀察花藥的育性,結果大白菜雄性不育系MS200712 不僅可以與大白菜品種正常雜交、結實,而且MS20071208-840和MS20071208-916的花藥依然表現為穩定的雄性不育性。但MS200712所屬的不育類型有待進一步確認。
3 結論與討論
目前,在大白菜育種中3種主要的細胞質雄性不育系Ogu CMS、Pol CMS 和CMS96 應用較多。但改良的Ogu CMS 不育系存在回交轉育多代后植株生長勢嚴重退化、雜交優勢差等缺點;Pol CMS 不育系由于溫度敏感存在育性受環境條件影響容易出現有粉株等缺點(張德雙,2006);CMS96 不育系雖然克服了上述缺點,但在實際應用中存在個別組合制種產量偏低的問題。因此,在大白菜育種中新型不育系的不斷推陳出新顯得尤為重要,不僅有利于提高雜交種的純度,保護育種者的利益,而且提供了深入開展雄性不育系機理探究的平臺。2004年10月筆者引入蕓薹屬異源六倍體材料,初衷是以蕓薹屬異源六倍體為橋梁,將紫色甘藍的紫色性狀轉到大白菜品種中,以期獲得類似于紫色甘藍葉片的紫色大白菜資源。首先,采用蕓薹屬異源六倍體與紫色甘藍進行正反交,試圖導入甘藍的紫色基因,實踐中發現,雖然每年的授粉工作量很大,但很難獲得真正的雜交種;然后,用蕓薹屬異源六倍體與大白菜雜交,結果發現二者可以正常雜交;于是,選用蕓薹屬異源六倍體與大白菜雜交,以期拓寬大白菜育種資源特別是雄性不育資源。
3.1 種間雜交拓寬了大白菜雄性不育系類型
在以往的研究中,有很多經由屬間或種間雜交途徑獲得雄性不育尤其是新型細胞質雄性不育的成功范本可供借鑒。Shiga 和Baba(1971,1973)在甘藍型油菜千莢油菜(Chisayanatane)×北陸23(Hokuriku 23)的F1中發現S-CMS 細胞質雄性不育(屬于Nap CMS)。Thompson(1972)在冬春甘藍型油菜RD58×Bronowski 的F2中發現T-CMS細胞質雄性不育(屬于Nap CMS),反交則花粉正常。李殿榮(1984)在甘藍型油菜S74-3(陜西)×〔豐收4號(四川)×7207(西藏)〕多親本雜交后代中發現雄性不育株,并育成1763A和6223A 雄性不育系。本試驗中先配制遠緣雜種(AABBCC×AA),再用普通大白菜品種義和5號作母本、(AABBCC×AA)作父本進行雜交,結果在義和5號×(蕓薹屬異源六倍體×金春)多親本雜交后代中首次發現雄性不育材料MS200712,這是本試驗的最大收獲。之所以選用春種大白菜金春和秋種大白菜義和5號作親本,是因為在進行雜交授粉時金春、義和5號與蕓薹屬異源六倍體的花期正好相遇。
3.2 大白菜雄性不育系MS200712 的敗育原因
以雄性不育系MS200712 為母本,分別與義和5號、北京新3號的父本832172 進行雜交,結果表明大白菜雄性不育系MS200712可以與大白菜品種正常雜交,并獲得回交二代MS20071208-840和回交一代MS20071208-916,但結實較少。經花期鑒定,播種的3株MS20071208-840和5株MS20071208-916全部表現為花藥敗育。
雄性不育系MS200712 為多親本雜交后代,細胞質來源于義和5號,導致雄性不育產生的可能原因:①蕓薹屬異源六倍體AABBCC 的細胞核基因與義和5號的細胞質基因互作結果;②金春的細胞核基因與義和5號的細胞質基因互作結果。但由于本試驗僅用義和5號獲得回交二代,鑒定的植株又少,目前很難判斷雄性不育產生的原因。今后的工作重點:①繼續進行回交轉育,增加回交代數,并跟蹤回交后代的育性;②著手配制(AABBCC×金春)×義和5號、金春×(AABBCC×義和5號)、(AABBCC× 義 和5號)× 金春、義和5號× 金春的多親本雜交后代,通過調查各后代單株育性,進一步解釋MS200712 雄性不育系產生的可能原因;③通過分子標記手段,開發鑒別雄性不育系的線粒體mtDNA(張揚勇 等,2011)、葉綠體cpDNA(張揚勇 等,2011;Wang et al.,2012)及核DNA 的SSR 等標記,進一步鑒定MS200712 雄性不育系類型。
本試驗中,大白菜雄性不育系MS200712 的獲取方法具有操作簡單、重復性好等優點,細胞質雄性不育系的不育性穩定、敗育徹底,且與其他大白菜品種雜交性良好、轉育容易。因此,可以繼續用于大白菜育種中的回交轉育,以獲得更多骨干雄性不育系材料。
今后,將開展有關大白菜雄性不育系MS200712 敗育機理的研究,包括細胞學、分子生物學等方面研究,力爭闡明大白菜雄性不育系MS200712 的所屬類型及其敗育機理,為該雄性不育系更好地在大白菜育種中應用及通過種間或屬間雜交途徑創造更多種類的雄性不育系奠定基礎。
柯桂蘭,趙稚雅,宋胭脂,張魯剛,趙利民.1992.大白菜異源胞質雄性不育系CMS3411-7的選育與應用.園藝學報,19(4):333-340.
李殿榮.1984.甘藍型油菜雄性不育系1763A 和6223A 的研究應用.陜西農業科學,(1):7-10.
張德雙,徐家炳,曹鳴慶,張鳳蘭.2002.大白菜轉育新型甘藍型油菜細胞質雄性不育系的研究.華北農學報,17(1):60-63.
張德雙.2006.大白菜CMS96 細胞質雄性不育的分子特性及育種應用研究〔博士論文〕.北京:中國農業科學院.
張德雙,張鳳蘭,余陽俊,趙岫云,于拴倉,徐家炳.2009.蕓薹屬異源六倍體的葉片突變體的獲得.中國蔬菜,(4):12-16.
張書芳,宋兆仲,趙雪云.1990.大白菜細胞核基因互作雄性不育系選育及應用模式.園藝學報,17(2):117-125.
張揚勇,方智遠,王慶彪,劉玉梅,楊麗梅,莊木,孫培田.2011.兩種甘藍Ogura 細胞質雄性不育源的分子鑒別.中國農業科學,44(14):2959-2965.
Health D W,Earle E D,Dickson M H.1994.Introgressing coldtolerant Ogura cytoplasm from rapeseed into Pak Choi and Chinese cabbage.HortScience,29:202-203.
Shiga T,Baba S.1971.Cytoplasmic male sterility in rape plants (Brassica napusL.).Japan J Breed,21:15-17.
Shiga T,Baba S.1973.Cytoplasmic male sterility in oilseed rape (Brassica napusL.)and its utilization to breeding.Japan J Breed,23:187-197.
Thompson K F.1972.Cytoplasmic male sterility in oil-seed rape plants.Heredity,29:253-257.
Wang Q B,Zhang Y Y,Fang Z Y,Liu Y M,Yang L M,Zhuang M.2012.Chloroplast and mitochondrial SSR help to distinguish allocytoplasmic male sterile types in cabbage(BrassicaoleraceaL.var.capitata).Mol Breeding,30:709-716.
Williams P H.1981.The origin and development of cytoplasmic male sterile Chinese cabbage.Taiwan:Asian Vegetable Research and Development Center:293-300.
