轉錄因子RUNX2對IPO8基因啟動子的調節作用
2014-03-21 11:26:06熊建軍龔幀周小鷗王庭劉建云李衛東
中國醫科大學學報 2014年7期
關鍵詞:檢測
熊建軍,龔幀,周小鷗,王庭,劉建云,李衛東
(1.九江學院基礎醫學院藥理學教研室,江西九江332000;2.江西省系統生物醫學重點實驗室,江西九江332000)
轉錄因子RUNX2對IPO8基因啟動子的調節作用
熊建軍1,龔幀1,周小鷗1,王庭1,劉建云2,李衛東2
(1.九江學院基礎醫學院藥理學教研室,江西九江332000;2.江西省系統生物醫學重點實驗室,江西九江332000)
目的研究轉錄因子RUNX2對IPO8基因啟動子的調控作用。方法基于PCR技術,分別突變和剔除IPO8基因啟動子片段P-732/+134上RUNX2結合位點(-497~-502 bp),改造后的啟動子片段定向插入熒光素酶表達載體PGL?3 Basic。重組質粒分別轉染Saos?2與HeLa細胞,檢測報告基因熒光素酶活性。應用染色質免疫共沉淀(ChIP)證實RUNX2與IPO8啟動子在Saos?2細胞內結合。慢病毒介導RUNX2在Saos?2細胞的靶向沉默,定量PCR檢測相應組別中IPO8mRNA的表達。結果成功突變和剔除IPO8啟動子上RUNX2結合位點,破壞RUNX2結合位點導致IPO8啟動子活性在Saos?2細胞中顯著下降(P<0.05)。轉錄因子RUNX2在Saos?2細胞與IPO8啟動子有效結合,下調RUNX2的表達導致IPO8轉錄水平下降(P<0.05)。結論轉錄因子RUNX2正性調控IPO8基因的轉錄。
IPO8;RUNX2;啟動子;熒光素酶
IPO8基因是核質轉運受體Importin β基因家族成員之一[1]。既往研究認為,IPO8是一個在細胞內穩定表達的基因,并在多種腫瘤細胞中作為參照基因用于PCR檢測[2,3]。但是近來研究顯示IPO8蛋白的功能并不局限于核質轉運,有文獻報道,IPO8的表達與眾多microRNA介導的基因沉默有著復雜的聯系,而抑制IPO8表達可導致部分microRNA靶基因上調[4],進而影響細胞內多個基因的表達水平。由此可見,IPO8的表達對于細胞的生理活動可能具有重要意義,但是IPO8基因表達的調控機制尚未見研究報道。前期研究中我們克隆了IPO8基因啟動子,發現其正性調控序列中含有一個典型的RUNX2結合位點。本研究通過定點突變、脂質體瞬時轉染、染色質免疫共沉淀(chromatin immunoprecipita?tion,CHIP)及RNA干擾等技術方法證實轉錄因子RUNX2對IPO8啟動子的轉錄調控作用。
1 材料與方法
1.1 細胞與主要試劑
人骨肉瘤細胞Saos?2、人宮頸癌細胞HeLa,均為本室保存;基因突變試劑盒TaKaRa Mutan BEST Kit,購自大連寶生物公司;轉染試劑Fugene6.0及雙熒光素酶檢測試劑盒,購自美國Promega公司;靶向沉默RUNX2基因的慢病毒顆粒(sc?37145?V)、RUNX2的羊多克隆抗體(sc?8566),購自美國Santa Cruz公司;McCOY′s 5A、DMEM培養基和胎牛血清,購于美國Gibco公司;ChIP試劑盒,購自美國Milli?pore公司;反轉錄試劑盒、實時PCR試劑盒,購自日本TOYOBO生物公司。
1.2 突變和剔除修飾RUNX2結合位點
以IPO8啟動子片段-732/+134連接的PMD?18載體為模板,分別設計合成隨機突變導入引物,剔除導入引物和對應的反向引物。突變引物序列:5′?AGCCTGACCTAAAAATCCTAGTACTC?3′;剔除引物序列:5′?ACCTAAAAATCCTAGTACTC?3′;反向引物序列:5′?TTATAACAAGATGAATTCCA?3′。PCR擴增,凝膠電泳后切膠回收目的DNA片段。產物DNA片段經Blunting Kination反應后進行自身連接反應,轉化感受態菌,挑選變異體,測序。將測序正確的修飾片段利用MluⅠ和BglⅡ酶切,定向插入PGL3?Basic,分別命名為P-732/RUNX2 MUT及P-732/RUNX2 DEL。
1.3 細胞的瞬時轉染與熒光素酶的檢測[5]
轉染前1 d將細胞按1×105/mL密度接種于24孔板,貼壁后12 h后采用Fugene 6.0介導質粒轉染。P-732/WT、P-732/RUNX2 MUT及P-732/RUNX2 DEL 3種質粒各0.5 μg,分別與內參質粒PRL?TK 0.1 μg混合轉染各孔。24 h后吸棄培養基,PBS洗滌細胞2次,隨后每孔中加入100 μL細胞裂解液,室溫水平搖動,以充分溶解轉染細胞。20 min后吸取細胞裂解液,12 000 g離心5 min,取50 μL上清液上機檢測,操作嚴格遵循試劑盒說明書,以螢火蟲熒光素酶/海腎熒光素酶的熒光活性值為相對熒光素酶表達量。
1.4 ChIP
在Saos?2細胞中進行CHIP分析的方法參照文獻[6]。RUNX2結合位點所在區域的PCR擴增引物為:5′?TTCACTTCGCCCATCCCTAT?3′,5′?CATTCATTCATTCGCTTTCA?3′。預期產物大小為240 bp。
1.5 慢病毒感染
靶細胞Saos?2培養于含10%胎牛血清的Mc?COY′s 5A培養基中,病毒感染前24 h消化,接種細胞密度為1×103/mL。待細胞生長鋪滿至50%,吸棄培養基,加入稀釋后的病毒上清液1 mL,并按8 μg/ mL濃度加入聚凝胺。細胞置32℃、5%CO2條件下培養12 h,吸棄培養液,McCOY′s 5A清洗2次,加入正常培養液于37℃、5%CO2條件下繼續培養72 h。
1.6 實時PCR
RNA提取、逆轉錄及實時PCR方法參照文獻。引物序列如下:IPO8:5′?TGTTCAGCTCCTTCCTGAT TC?3′,5′?CTTCTTACACTTCCACCATAC?3′;RUNX2:5′?GCCTTCAAGGTGGTAGCCC?3′,5′?AAGGTGAAACTCTTGCCTCGTC?3′;GAPDH:5′?AGAAGGCTGGGGCTCATTTG?3′,5′?AGGGGCCAT CCACAGTCTTC?3′。
1.7 統計學處理
應用統計學軟件SPSS 12.0進行相應的統計學分析。2組間比較采用t檢驗。P<0.05為差異有統計學意義。
2 結果
2.1 突變和剔除修飾IPO8啟動子上RUNX2結合位點
近期我們研究IPO8啟動子區域時,發現一個經典的RUNX2結合位點,位于轉錄起始點上游(-497~-502 bp)。為了證實該位點是否參與了IPO8基因的轉錄調控,我們對該位點進行突變和剔除修飾。測序結果顯示,成功突變和剔除IPO8啟動子RUNX2結合位點(圖1)。
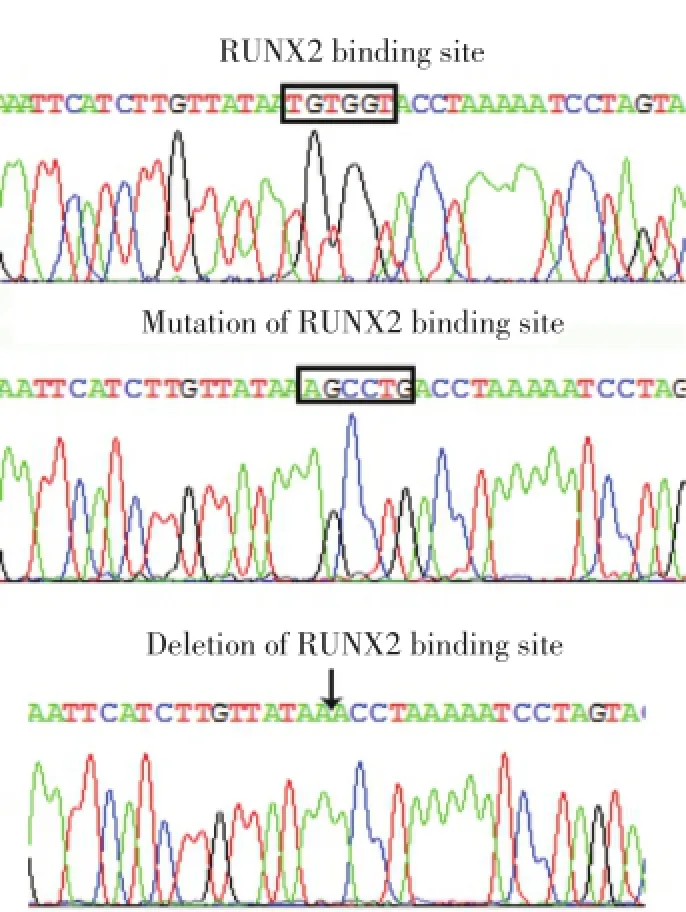
圖1 IPO8啟動子上RUNX2結合位點的測序圖Fig.1 DNA sequencing for RUNX2 binding site in IPO8 promoter
2.2 破壞RUNX2結合位點導致IPO8啟動子活性在Saos?2細胞中下降
為研究RUNX2結合位點對IPO8基因啟動子活性的影響,我們將重組質粒P-732/WT、P-732/ RUNX2 MUT及P-732/RUNX2 DEL分別轉染Saos?2細胞,經雙熒光素酶報告系統檢測螢光素酶的活性值,結果顯示,重組質粒P-732/RUNX2 MUT及 P-732/RUNX2 DEL在Saos?2細胞中熒光素酶活性與P-732/WT相比下降約40%。
為進一步證實RUNX2對IPO8啟動子的調控作用,我們將以上3種質粒分別轉染入RUNX2缺失的HeLa細胞,通過雙熒光檢測儀檢測熒光素酶活性。結果顯示,3組質粒的熒光素酶活性無統計學差異(P>0.05),見圖2。
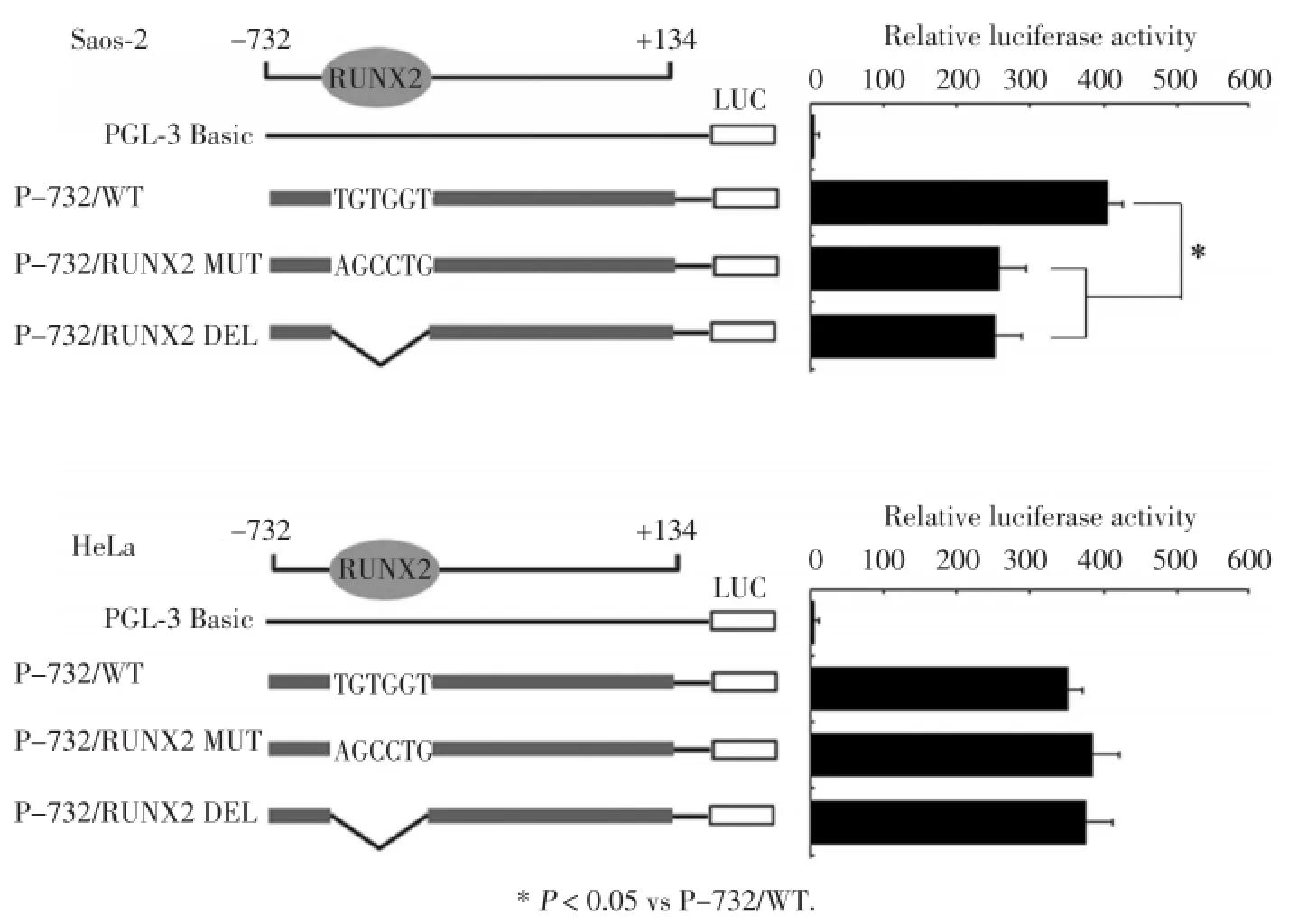
圖2 突變型和剔除型IPO8啟動子報告重組體熒光素酶活性Fig.2 Luciferase activity of the mutant or deleted IPO8promoter reporter constructs
2.3 RUNX2在Saos?2細胞內與IPO8啟動子結合
為了探索轉錄因子RUNX2是否與IPO8啟動子在Saos?2細胞內有效結合,本研究對從Saos?2細胞提取的染色質進行ChIP分析。染色質經IgG或RUNX2抗體免疫共沉淀后,經PCR擴增顯示,內源性RUNX2結合在IPO8啟動子區(圖3)。
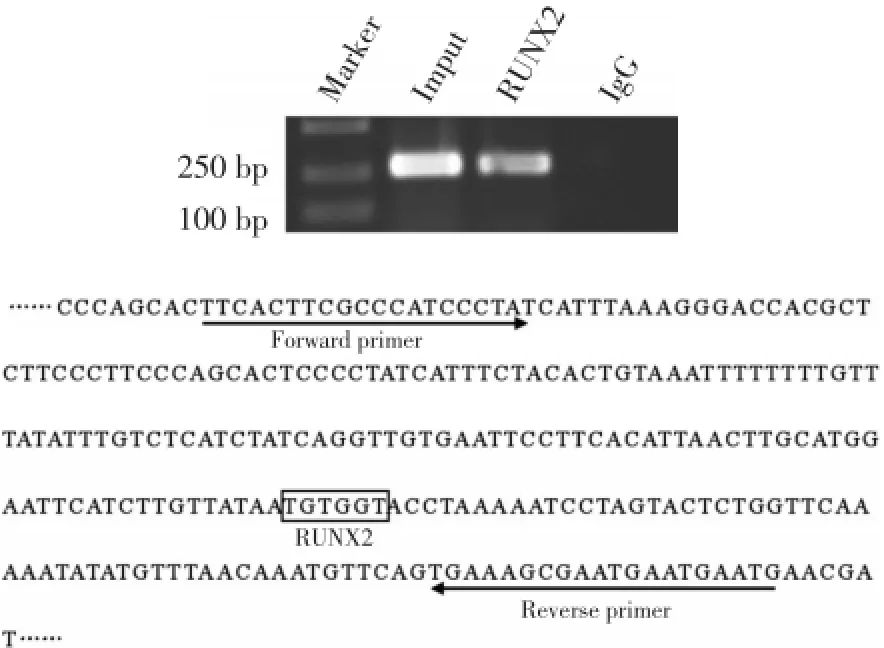
圖3 RUNX2結合在IPO8啟動子Fig.3 Identification of RUNX2 binding to the IPO8promoter
2.4 下調RUNX2表達抑制IPO8轉錄
為了檢測下調RUNX2的表達對IPO8轉錄活性的影響,慢病毒介導的RNA干擾下調Saos?2細胞中80%左右RUNX2mRNA的表達。進一步結果顯示,RUNX2下調的同時導致IPO8轉錄活性顯著下降(圖4),表明RUNX2表達與IPO8的轉錄水平存在正性調控關系。
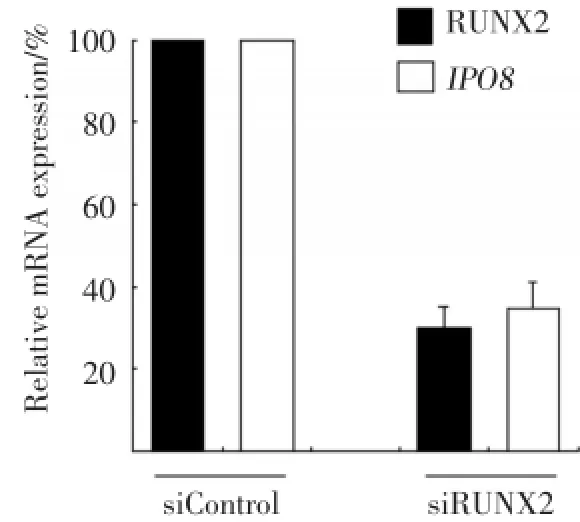
圖4 沉默RUNX2抑制Saos?2細胞中IPO8mRNA表達Fig.4 Silencing of RUNX2 reduces IPO8 mRNA expression in Saos?2 cells
3 討論
核轉運受體包括Importin α和Importin β兩個家族,分別識別不同的蛋白質核定位信號,在蛋白質入核的主動運輸過程中發揮重要作用[7]。本研究中的IPO8即是Importin β家族成員之一,定位于人類常染色體的12p11.21。
有研究認為,IPO8是一個在細胞內穩定表達的基因,在多種腫瘤細胞中可作為參照基因用于PCR檢測[2,3]。但還有研究顯示,IPO8蛋白的表達對細胞生理活動有直接影響,例如抑制IPO8表達可導致部分miRNA靶基因的上調,進而影響細胞的生理功能,其原因在于IPO8蛋白與Argonaute蛋白的相互作用是介導靶mRNA降解的必經環節[4]。此外,IPO8也是Smad蛋白、糖皮質激素受體入核轉運的主要核受體[8,9]。由此可見,IPO8的表達對于細胞正常功能具有重要意義,但是IPO8表達的調控機制尚不清楚。
利用生物信息學分析,我們發現一個典型的RUNX2結合位點,位于IPO8啟動子的正性調控區域,而熒光素酶實驗顯示包含該區域的5′端非編碼區具有最高的轉錄活性(另文發表),這促使我們對RUNX2調控IPO8轉錄的可能性進行驗證。實驗結果顯示,突變或剔除RUNX2結合位點可導致IPO8啟動子片段活性在Saos?2細胞明顯下降,而在無RUNX2表達的HeLa細胞內,以上差異則不明顯。隨后ChIP實驗也進一步證實,在Saos?2細胞內RUNX2與IPO8啟動子中RUNX2結合位點所在區域(-424~-663 bp)有著直接結合。最后,我們利用慢病毒介導RUNX2在Saos?2細胞的下調,發現IPO8mRNA水平隨之明顯下降。基于以上研究結果,我們認為轉錄因子RUNX2與IPO8啟動子相互作用是調控IPO8轉錄的重要因素。
RUNX2蛋白結構含有1個與DNA結合的runt結構域、3個轉錄激活區、1個轉錄抑制區和1個短的核定位信號,通過識別靶基因啟動子中TGTGGT序列,進而調控相關基因的轉錄[10]。RUNX2的3種異構體分別在成骨細胞的早期增殖和后期成熟中的多種信號途徑中起中心作用[11]。此外,RUNX2也被認為與多種腫瘤的發生、進展有內在聯系[12]。基于RUNX2對IPO8基因在轉錄水平上的表達調控,我們推測IPO8也許存在眾多未知的功能,而這些功能可能與成骨分化或腫瘤發生、進展有關。
[1]Gorlich D,Dabrowski M,Bischoff FR,et al.A novel class of RanGTP binding proteins[J].J Cell Biol,1997,138(1):65-80.
[2]Hurtado del Pozo C,Calvo RM,Vesperinas?Garcia G,et al.IPO8 and FBXL10:new reference genes for gene expression studies in hu?man adipose tissue[J].Obesity(Silver Spring),2010,18(5):897-903.
[3]Kreth S,Heyn J,Grau S,et al.Identification of valid endogenous control genes for determining gene expression in human glioma[J]. Neuro Oncol,2010,12(6):570-579.
[4]Weinmann L,Hock J,Ivacevic T,et al.Importin 8 is a gene silenc?ing factor that targets argonaute proteins to distinct mRNAs[J]. Cell,2009,136(3):496-507.
[5]熊建軍,周英妹,干麗君,等.原癌基因USP22啟動子的克隆及轉錄活性分析[J].中國醫科大學學報,2011,40(7):580-583.
[6]Xiong J,Che X,Li X,et al.Cloning and characterization of the hu?man USP22 gene promoter[J].PLoS One,2012,7(12):e52716.
[7]Cook A,Bono F,Jinek M,et al.Structural biology of nucleocytoplas?mic transport[J].Annu Rev Biochem,2007,76:647-671.
[8]Yao X,Chen X,Cottonham C,et al.Preferential utilization of Imp7/ 8 in nuclear import of Smads[J].J Biol Chem,2008,283(33):22867-22874.
[9]Freedman ND,Yamamoto KR.Importin 7 and importin alpha/impor?tin beta are nuclear import receptors for the glucocorticoid receptor[J].Mol Biol Cell,2004,15(5):2276-2286.
[10]van der Deen M,Akech J,Lapointe D,et al.Genomic promoter oc?cupancy of runt-related transcription factor RUNX2 in osteosarco?ma cells identifies genes involved in cell adhesion and motility[J]. J Biol Chem,2012,287(7):4503-4517.
[11]Stock M,Otto F.Control of RUNX2 isoform expression:the role of promoters and enhancers[J].J Cell Biochem,2005,95(3):506-517.
[12]Barnes GL,Javed A,Waller SM,et al.Osteoblast-related transcrip?tion factors Runx2(Cbfa1/AML3)and MSX2 mediate the expres?sion of bone sialoprotein in human metastatic breast cancer cells[J].Cancer Res,2003,63(10):2631-2637.
(編輯 陳姜)
Regulation of Human IPO8 Promoter by Transcription Factor RUNX2
XIONG Jian?jun1,GONG Zhen1,ZHOU Xiao?ou1,WANG Ting1,LIU Jian?yun2,LI Wei?dong2
(1.Department of Pharmacology,College of Basic Medical Science,Jiujiang University,Jiujiang 332000,China;2.Key Laboratory of Jiangxi Province for the Systems Biomedicine,Jiujiang 332000,China)
ObjectiveTo investigate the regulation effects of transcription factor RUNX2 on human importin 8(IPO8)promoter.MethodsMu?tation or deletion of the RUNX2?binding site(-497 to-502 bp)within theIPO8promoter region was performed by PCR technology.The modifiedIPO8promoters were subsequently inserted into luciferase report vector PGL?3 Basic.The recombinant plasmids were transfected into human Saos?2 and HeLa cells respectively and the reporter gene luciferase activities were measured by dual luciferase reporter system.Furthermore,ChIP assay was performed to confirm the binding of RUNX2 toIPO8promoter in Saos?2 cells.In addition,lentivirus?mediated siRNA down?regulated RUNX2 expression in Saos?2 cells and expression ofIPO8mRNA was determined by real?time PCR.ResultsThe RUNX2 binding site was mutated and deleted withinIPO8promoter.Disruption of RUNX2 binding site led to significant decrease ofIPO8promoter activity in Saos?2 cells(P<0.05). ChIP assay demonstrated that RUNX2 binds toIPO8promoter.Down?regulation of RUNX2 attenuated the transcription level ofIPO8gene(P<0.05).ConclusionTranscription factor RUNX2 positively regulates the activity ofIPO8promoter.
IPO8;RUNX2;promoter;luciferase
R34
A
0258-4646(2014)07-0585-04
國家自然科學基金(81060075,81260140);江西省教育廳科學研究項目(GJJ12683)
熊建軍(1975-),男,副教授,博士.
李衛東,E-mail:lwd626518@163.com
2014-04-22
網絡出版時間:
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
