互花米草入侵對西門島潮灘底棲食物網(wǎng)的影響
2014-05-30 05:13:56溫作柱廖忠鷺
浙江農(nóng)業(yè)科學(xué) 2014年5期
溫作柱,廖忠鷺,李 平,商 栩
(溫州醫(yī)科大學(xué),浙江 溫州 325000)
作為海陸過渡帶,人類活動對濕地的影響較為劇烈,使其生態(tài)環(huán)境脆弱,自然災(zāi)害頻繁,是全球變化的敏感區(qū)域[1]。基于海岸防護(hù)、保灘促淤的目的,我國于1972年從原產(chǎn)地美國引入互花米草(Spartina alterniflora),目前已分布到我國沿海的大部分淤泥質(zhì)灘涂上,北從天津,南至廣西都形成面積大小不等的互花米草潮灘濕地[2]。由于互花米草適應(yīng)范圍廣繁殖能力強,其種群已對我國潮灘濕地生態(tài)系統(tǒng)構(gòu)成嚴(yán)重威脅,成為入侵我國沿海潮灘影響最嚴(yán)重的外來植物[3]。
近幾十年來,生態(tài)學(xué)家試圖闡明互花米草入侵的生態(tài)后果,包括植物的競爭、當(dāng)?shù)厣锒鄻有缘母淖儭⑸车耐嘶蜕锏厍蚧瘜W(xué)過程的改變等[4-7]。其中互花米草對底棲生態(tài)群落的影響受到廣泛關(guān)注。沈永明等[6]認(rèn)為,江蘇省海岸的互花米草使灘面淤高,在侵蝕型岸段,互花米草的入侵會使泥螺、四角蛤蜊、文蛤等灘涂貝類的生存場所消失;但也有人認(rèn)為,互花米草入侵后可能提高原有底棲群落物種豐度和多樣性。然而,關(guān)注這種外來植物入侵對鹽沼濕地營養(yǎng)功能和基礎(chǔ)影響的研究相對較少[8-9],加之對本土潮灘濕地底棲食物網(wǎng)結(jié)構(gòu)認(rèn)識的不足,為深入探討入侵生物可能導(dǎo)致的危害造成了瓶頸。此外,不管是土著植物還是入侵植物,都會因其高生物量而習(xí)慣性地被認(rèn)為是此處食物網(wǎng)的營養(yǎng)基礎(chǔ)。大量研究表明,微型藻類才是潮灘食物網(wǎng)的能量來源。鑒于微型藻類和植物碎屑之間能量基礎(chǔ)的爭論以及目前不同初級生產(chǎn)者對底棲食物網(wǎng)貢獻(xiàn)大小認(rèn)識不足,本研究通過估算不同初級生產(chǎn)者對底棲食物網(wǎng)中各消費者的優(yōu)勢種類的相對營養(yǎng)貢獻(xiàn)大小,評價互花米草在潮灘濕地底棲食物網(wǎng)的地位和作用,從底棲動物營養(yǎng)來源的角度闡述互花米草入侵后對潮灘濕地生態(tài)系統(tǒng)及灘涂養(yǎng)殖業(yè)的影響。
1 材料與方法
1.1 樣品采集
西門島潮灘濕地原本是溫州典型的紅樹林濕地,互花米草入侵后,迅速擴(kuò)張,紅樹林濕地很快被互花米草覆蓋,成為入侵嚴(yán)重的典型區(qū)域。
分別于2011年3-4月在樂清灣西門島潮灘濕地采集底棲微藻 (MPB)、土著紅樹植物秋茄(Kandelia candel)、入侵植物互花米草等初級生產(chǎn)者樣品,以及小型和大型底棲動物、游泳動物等消費者類群的代表種類樣品。其中底棲微藻采用Blanchard等[10]的方法收集后,過濾到 Whatman 0.45 μm GF/F濾膜上,于-20℃下避光冷凍保存。懸浮顆粒物 (POM)則直接過濾到GF/F模上,于-20℃下避光冷凍保存。秋茄和互花米草葉片從采樣點不同簇個體中手工摘取,于現(xiàn)場潮溝水中洗凈后冷凍保存。
浮游動物樣品于潮溝中用孔徑160 μm的浮游生物采集網(wǎng)在水體表層進(jìn)行水平拖網(wǎng)采集獲得,活體帶回實驗室后于XTL-3400體視顯微鏡下挑出優(yōu)勢種類個體。包含小型底棲動物的沉積物樣品,經(jīng)上層1mm,下層50 μm的套篩淘洗后,將留于50 μm篩網(wǎng)上的線蟲 (Nematoda)和猛水蚤 (Harpacticoid spp.)于體視顯微鏡下分別挑出。以上小型消費者均活體于過濾后的現(xiàn)場海水中過夜,以排空腸道,在-20℃下避光冷凍保存。優(yōu)勢腹足類短擬沼螺 (Assiminea brevicula)、織紋螺(Nassariidae),優(yōu)勢雙殼類縊蟶(Sinonovacula constricta)、菲律賓簾蛤 (Ruditapes philippinarum),優(yōu)勢底棲魚類大彈涂魚(Periophthalmini)的樣品均為手工從潮灘、潮溝表面采集。優(yōu)勢短尾類甲殼動物譚氏泥蟹(Ilyoplax deschampsi)、招潮蟹 (Uca)以及沙蠶(Nereis)和紐蟲 (Nemertina)用小刀從中潮灘和潮溝邊灘沉積物中挖出。優(yōu)勢雙殼類、寡毛類則用孔徑為1mm的篩網(wǎng)淘洗潮溝底部沉積物獲得,并于-20℃下避光冷凍保存。
1.2 樣品處理
所有植物樣品經(jīng)自來水沖洗干凈后,再用Milli-Q水清洗,以去除其附著物,在60℃烘箱內(nèi)干燥48 h,然后用不銹鋼粉碎機進(jìn)行粉碎。濾有底棲微藻的GF/F濾膜在60℃下烘干24 h。因浮游動物和小型底棲動物的單個個體不足以提供足夠的生物量用于穩(wěn)定同位素測定,所以根據(jù)其個體大小選取10~100只合并為1個樣品,用Milli-Q水清洗后于60℃下烘干24 h。對個體較大的大型底棲動物和游泳動物則分別分離其腹足部分 (腹足類和雙殼類)、大螯 (蟹)和背部 (魚)的肌肉,用Milli-Q水漂洗后于60℃下烘干24 h,并研為粉末。所有樣品烘干前均在體視顯微鏡下將混雜的貝殼、骨骼及碎屑等挑出。由于酸洗過程被認(rèn)為對δ13C值的影響顯著,并可能明顯改變δ15N值,且酸洗可能造成平行樣品間所測得δ13C和δ15N的值差異增大,由此所有動物和植物樣品均未作酸化處理[11-12]。
1.3 穩(wěn)定同位素測定及數(shù)據(jù)分析
穩(wěn)定同位素的測定通過同位素質(zhì)譜儀 (Delta plus XP Finnigan)完成,其中用于δ13C測定所需要的干重樣品量0.2~0.3mg,δ15N測定所需要的干重樣品量0.5~1.0mg。同位素豐度計算方法:
δX/% = [(Rstandard/Rstandard)-1]× 103。
式中,X=13C或15N,R=13C/12C或15N/14N,標(biāo)準(zhǔn)采用的是PDB國際通用標(biāo)準(zhǔn) (Pee Dee Belelmnite標(biāo)準(zhǔn))。δ13C的分析精度為 ±0.01%,δ15N的分析精度為±0.02%。
測得的碳、氮穩(wěn)定同位素用 IsoSource(http://www. epa. gov/wed/pages/models/stableIsotopes/isotopes.htm)對各初級生產(chǎn)者的營養(yǎng)貢獻(xiàn)進(jìn)行分析。IsoSource方法是將各種源的組成比例之和定為100%,首先以預(yù)先定制的增量(Source increment,通常設(shè)為1%)把所有可能的組合計算出來,然后用基于質(zhì)量守恒方程基礎(chǔ)上的線性混合模型計算各種組合中得到的混合物的預(yù)期穩(wěn)定同位素值。將計算得到的混合物穩(wěn)定同位素值與實際測定值進(jìn)行比較,則在一定容差范圍 (如0.01%)內(nèi),所有符合實際穩(wěn)定同位素值的計算所得組合就反映了各種源在混合物中所占比例。當(dāng)實際應(yīng)用于食物網(wǎng)研究時,由于δ13C和δ15N在攝食過程中存在逐營養(yǎng)級分餾現(xiàn)象,因此在進(jìn)行IsoSource分析時需根據(jù)不同消費者及其送測組織的穩(wěn)定同位素分餾特征扣除富集值。
本文使用的δ13C逐營養(yǎng)級富集為0.1%,δ15N則為0.34%。分別把互花米草、秋茄、懸浮顆粒有機物和底棲硅藻等4個源作為主要有機質(zhì)食物源。參考Benstead等[13]研究方法,計算中將增量設(shè)為1%,容差初始設(shè)為0.01%,但如果混合物的同位素值超出了計算閾值,即超出了由食物網(wǎng)中的若干個源所組成的多邊形[14],則將容差最多增加到0.05%。
2 結(jié)果與分析
2.1 各主要初級生產(chǎn)者的δ13C和δ15N值
碳、氮穩(wěn)定同位素測定結(jié)果顯示 (表1),各初級生產(chǎn)者的 δ13C值范圍較廣 (-2.6% ~-1.3%),且新鮮植株和枯萎植株的δ13C值差異不大。外來C4植物互花米草比土著C3植物秋茄有顯著富集的δ13C值 (-1.34%)。
底棲微藻 (MPB)和懸浮有機顆粒 (POM)的δ13C值均介于C3和C4植物間且差異不大。6種初級生產(chǎn)者δ15N值均在0.27% ~0.58%,相比δ13C,各初級生產(chǎn)者間 δ15N差異不明顯(表2)。
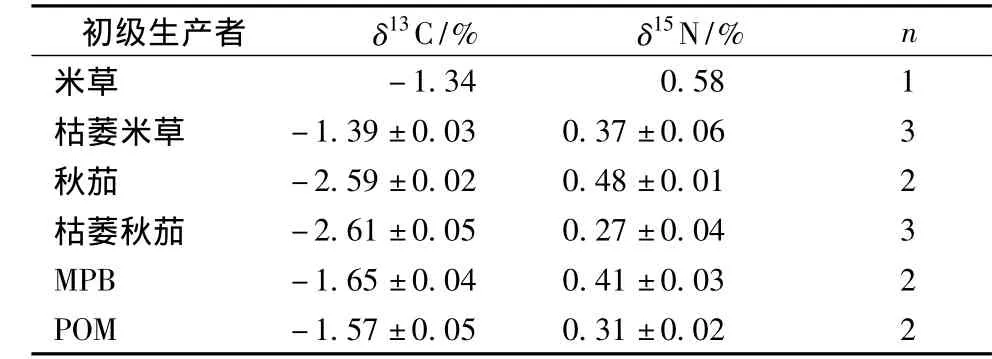
表1 各初級生產(chǎn)者碳、氮穩(wěn)定同位素值
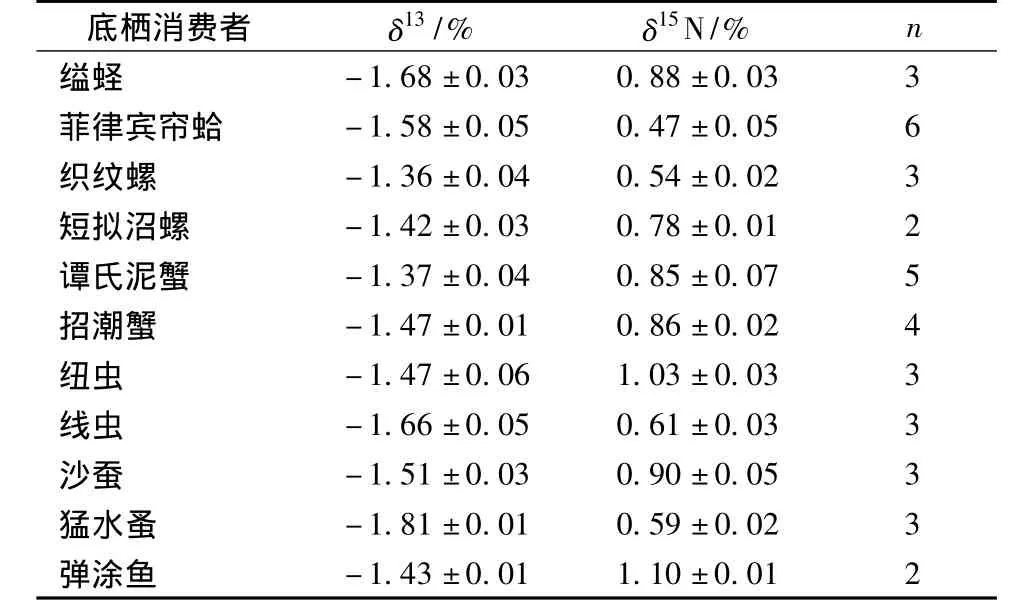
表2 各底棲消費者碳、氮穩(wěn)定同位素值
2.2 各主要底棲消費者的δ13C和δ15N值
表2顯示,各底棲動物間δ13C值差異不大(-1.8%~-1.3%)。其中小型底棲動物猛水蚤具有最貧δ13C值 (-1.81±0.01)%,此外小型底棲動物線蟲δ13C值為 (-1.66±0.05)%。大型底棲動物中織紋螺具有最富 δ13C值 (-1.36±0.04)%,幾乎與米草δ13C值重疊 (圖1)。其他大型底棲動物的δ13C值分布各異,基本上介于MPB和米草之間 (圖1)。西門島潮灘濕地主要底棲消費者的δ15N值介于0.47% ~1.10%,顯示該食物網(wǎng)中可能存在多個營養(yǎng)級,彈涂魚是本研究涉及生物中營養(yǎng)級最高的。
2.3 西門島灘涂濕地主要生產(chǎn)者和底棲消費者δ13 C和δ15N的關(guān)系
從西門島潮灘濕地食物網(wǎng)各主要生產(chǎn)者和底棲消費者的碳、氮穩(wěn)定同位素特征圖 (圖1)可以看出,大部分底棲動物的δ13C值均處于MPB和C4植物米草之間,表明δ13C值稍貧的MPB必然存在于這些底棲動物的食譜中。而POM的δ13C值與MPB非常接近,也可能是一些底棲動物的食物來源。而小型底棲動物猛水蚤的δ13C值略高于MPB,落在秋茄和MPB之間,表明δ13C值最貧的C3植物秋茄也可能對其有一定的營養(yǎng)貢獻(xiàn)。
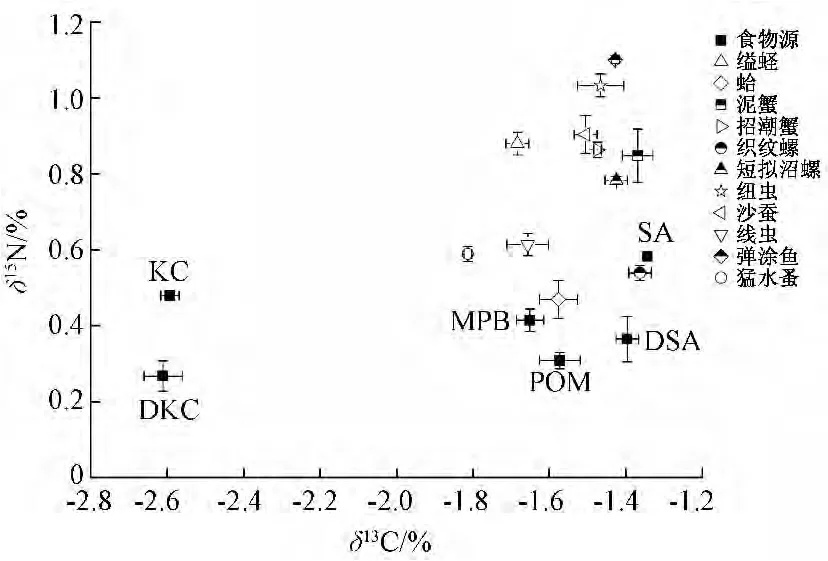
圖1 西門潮灘沼濕地各主要初級生產(chǎn)和優(yōu)勢底棲消費者δ13C和δ15N的關(guān)系
為了更加清晰地看出各生產(chǎn)者和底棲消費者之間的營養(yǎng)特征,繪制出扣除營養(yǎng)富集值后的碳、氮穩(wěn)定同位素特征校正圖 (圖2)。
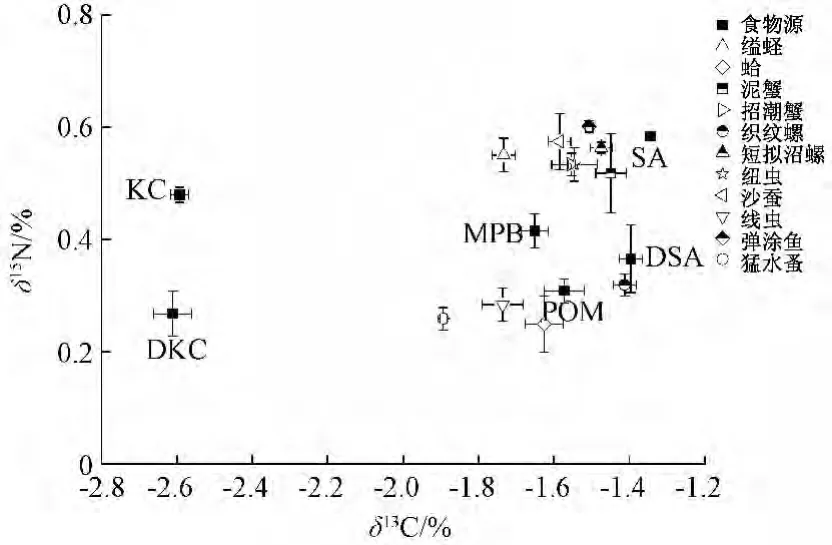
圖2 西門島潮灘濕地各主要初級生產(chǎn)和優(yōu)勢底棲消費者δ13C和δ15N的關(guān)系 (校正后)
圖2顯示,大部分大型底棲消費者的碳、氮穩(wěn)定同位素特征分布非常接近米草,表明C4植物互花米草很可能是這些大型底棲消費者重要的營養(yǎng)來源;與其他大型底棲動物不同,菲律賓簾蛤和小型底棲動物猛水蚤、線蟲的穩(wěn)定同位素特征更靠近POM,表明這些底棲消費者更多從POM(或者浮游植物)獲取營養(yǎng)支持。相反,C3植物秋茄的δ13C值遠(yuǎn)低于這些底棲消費者 (差值≥0.8%),不大可能成為其營養(yǎng)來源。MPB在整個生產(chǎn)者和底棲消費者的穩(wěn)定同位素特征分布中處于居中的位置,不能排除其對整個底棲食物網(wǎng)的營養(yǎng)貢獻(xiàn)。
2.4 Isosource計算所得各初級生產(chǎn)者對消費者的可能營養(yǎng)貢獻(xiàn)
圖3顯示了6種初級生產(chǎn)者對11中底棲消費者營養(yǎng)貢獻(xiàn)的可能范圍。從圖可以看出,C4植物互花米草新鮮植株對7種大型底棲動物 (彈涂魚、短泥沼螺、譚氏泥蟹、縊蟶、紐蟲、沙蠶、招潮蟹)各自的平均營養(yǎng)貢獻(xiàn)在66.2%以上,其中對譚氏泥蟹的最小貢獻(xiàn)最低,但也高達(dá)58%,反映出入侵C4植物互花米草對這些大型底棲動物較為重要的營養(yǎng)地位。新鮮互花米草對其他4種底棲動物各自最大貢獻(xiàn)可能都很低 (<12%),且其最小貢獻(xiàn)可能均為0,表現(xiàn)出較為不重要的營養(yǎng)地位。但是計算所得枯萎米草對織紋螺的貢獻(xiàn)可能為47%~100%,營養(yǎng)地位明確;雖然枯萎米草對蛤有著相對高的最大貢獻(xiàn)可能 (52%),但其為0的最小貢獻(xiàn)可能,使得營養(yǎng)地位無法確定。與C4植物互花米草相比,不管是新鮮或者枯萎的秋茄,雖然其對一些底棲動物 (例如沙蠶、縊蟶和猛水蚤)有著較為明確的營養(yǎng)貢獻(xiàn) (分別為14% ~20%,26% ~32%,22% ~40%),但總體上C3植物秋茄對整個底棲食物網(wǎng)的營養(yǎng)貢獻(xiàn)偏低,反映其較低的營養(yǎng)重要性。底棲微藻 (MPB)對所有底棲動物的最大貢獻(xiàn)均不高 (最大為37%),且最小貢獻(xiàn)可能均為零,反映其較為有限的營養(yǎng)貢獻(xiàn)。POM對2種小型底棲動物 (猛水蚤和線蟲)的營養(yǎng)地位較為明確 (營養(yǎng)貢獻(xiàn)可能分別為31% ~73%,53%~87%);而對蛤的貢獻(xiàn)可能為27% ~100%,可能有一定的營養(yǎng)地位,但其較大的貢獻(xiàn)分布跨度增大了POM對這種底棲動物營養(yǎng)地位的不確定性。
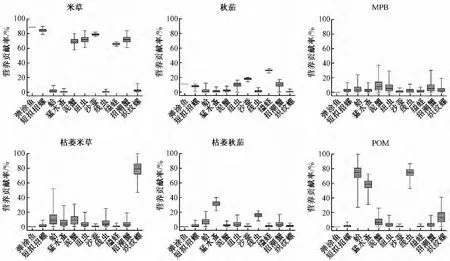
圖3 西門島鹽沼濕地各初級生產(chǎn)者對底棲消費者的營養(yǎng)貢獻(xiàn)
3 小結(jié)和討論
3.1 不同初級生產(chǎn)者的相對營養(yǎng)貢獻(xiàn)
維管植物和微型藻類對潮灘濕地食物網(wǎng)的相對重要性目前仍有爭論[15-18]。相比維管植物纖維質(zhì)的細(xì)胞壁,主要由硅藻組成的底棲微藻的外殼是較易被消化的硅質(zhì)薄殼,單細(xì)胞的底棲微藻更容易被底棲消費者攝取。由此長久以來濕地研究者就推測底棲微藻對鹽沼消費者有著更高的營養(yǎng)價值[15]。然而本研究結(jié)果顯示,在西門島潮灘濕地底棲食物網(wǎng)中,互花米草可能是大型底棲動物更為重要的營養(yǎng)來源,而底棲微藻的營養(yǎng)貢獻(xiàn)則非常有限。這可能是由于在該處潮間帶區(qū)域高密度的互花米草覆蓋了灘涂表面,使得互花米草叢中的底棲微藻難以獲得足夠的光照而無法提供較高的生產(chǎn)力和生物量。此外也有研究指出,互花米草入侵可能改變原有食物網(wǎng)的能量基礎(chǔ),由入侵前以藻類為主轉(zhuǎn)變?yōu)橐杂袡C碎屑為主要食源[19]。這種食物網(wǎng)能量基礎(chǔ)的改變至少部分被上行效應(yīng)所調(diào)控,即底棲微藻生物量的降低和植物碎屑數(shù)量的增加改變了鹽沼濕地消費者的食源構(gòu)成[19],驅(qū)動大型底棲動物同化更多的互花米草有機碎屑,甚至其營養(yǎng)比重超過底棲微藻。
浮游植物細(xì)胞 (直徑2~15 μm)通常較小,除懸浮物取食者 (雙殼類)外,潮灘濕地上的沉積物取食者 (如螺類和蟹類)并不能有效濾食水體中的浮游藻類[20]。另外,由于較短的潮淹時間,生活于高、中潮區(qū)的大型無脊椎動物很少能夠有效地利用水體中的浮游植物初級生產(chǎn)。本研究結(jié)果顯示,POM與MPB一樣,對絕大多數(shù)大型底棲動物(除蛤外)的營養(yǎng)貢獻(xiàn)非常有限;然而,對小型底棲動物 (猛水蚤和線蟲)的營養(yǎng)比重卻截然不同:浮游植物可能是這2種小型底棲動物最主要的食物來源。盡管沉積食性動物攝取浮游植物的效率比濾食性動物低,但沉積到潮灘上的浮游植物仍可能在底棲猛水蚤的食譜中占有一定比重[21],而自由生活的線蟲同樣可能利用這些沉積下來的易分解的有機碳[22]。
盡管有人認(rèn)為C3植物和米草共同出現(xiàn)在同一潮灘濕地時,C3植物的營養(yǎng)貢獻(xiàn)可能超過米草[23],相比米草難以分解的木質(zhì)素組成,其有機產(chǎn)出更容易通過細(xì)菌途徑被利用[24]。但是,IsoSource計算結(jié)果顯示,土著C3植物對整個底棲食物網(wǎng)的營養(yǎng)貢獻(xiàn)并非如想象的重要,僅對一些底棲動物 (縊蟶、沙蠶、猛水蚤、線蟲)有著相對明確的營養(yǎng)貢獻(xiàn) (圖3),但與米草和POM相比,C3植物更可能是作為補充性的食物來源。
3.2 互花米草入侵對鹽沼底棲食物網(wǎng)的影響
米草入侵后改變當(dāng)?shù)卦械牡讞珓游锝M成和數(shù)量的事例眾多,也有學(xué)者預(yù)測其可能改變食物網(wǎng)結(jié)構(gòu)[25-26]。但近期才有少數(shù)研究關(guān)注到米草入侵后對原有營養(yǎng)結(jié)構(gòu)造成的影響,并證明鹽沼生態(tài)系統(tǒng)中存在著由營養(yǎng)基礎(chǔ)自下而上對底棲動物群落結(jié)構(gòu)的控制[19]。雖本研究并未調(diào)查原有土著C3植物群落中底棲動物的生物組成和營養(yǎng)結(jié)構(gòu),但調(diào)查中發(fā)現(xiàn),土著紅樹植物秋茄的分布非常稀疏,互花米草已占據(jù)絕大部分灘面。互花米草強大的競爭優(yōu)勢及其快速擴(kuò)張顯著壓縮了原有C3植物和底棲微藻的生長空間,進(jìn)而可能改變生活在該濕地內(nèi)許多土著動物的食物基礎(chǔ)。
對西門島潮灘濕地中主要生產(chǎn)者和消費者的穩(wěn)定同位素分析顯示,入侵種互花米草是這一區(qū)域底棲食物網(wǎng)最重要的食物來源之一。優(yōu)勢種中僅有2種小型底棲動物 (猛水蚤和線蟲)和1種雙殼類 (蛤)未表現(xiàn)出對互花米草的明顯攝取,互花米草對西門島鹽沼濕地中的營養(yǎng)地位與其原產(chǎn)地相似。這一結(jié)果表明,對于絕大多數(shù)的大型底棲動物來說,外來種互花米草已成為其重要的食物源,因此對于這些大型底棲動物的生存可能暫時未受到威脅。但與Levin等[19]在舊金山海灣鹽沼濕地的發(fā)現(xiàn)類似,西門島潮灘濕地中一些重要的次級生產(chǎn)者 (蛤、猛水蚤、線蟲)并沒有或者很少攝入入侵植物 (互花米草)的有機碳,這可能會導(dǎo)致一些關(guān)鍵物種的消失,并進(jìn)而影響其他物種乃至整個生態(tài)系統(tǒng)結(jié)構(gòu)和功能,引起濱海養(yǎng)殖業(yè)的經(jīng)濟(jì)創(chuàng)傷。
[1]Hassen M B.Spatial and temporal variability in nutrients and suspended material processing in the Fier d'Ars Bay(France)[J].Estuarine,Coastal and Shelf Science,2001,52(4):457-469.
[2]徐國萬,卓榮宗.我國引種互花米草的初步研究.南京大學(xué)學(xué)報 (米草研究的進(jìn)展:22年來的研究成果論文集)[M].南京:南京大學(xué)出版社,1985:212-225.
[3]郭云文,陳莉麗,盧百靈,等.我國對互花米草的研究進(jìn)展[J].草業(yè)與畜牧,2007(9):1-6.
[4]Fell P E.Does invasion of oligohaline tidal marshes by reed grass,Phragmites australis(Cav.)Trin.ex Steud.,affect the availability of prey resources for the mummichog,F(xiàn)undulus heteroclitus L? [J].Journal of Experimental Marine Biology and Ecology,1998,222(2):59-77.
[5]Wei J S,Weis P.Is the invasion of the common reed,Phragmites australis,into tidal marshes of the eastern US an ecological disaster?[J].Marine Pollution Bulletin,2003,46(7):816-820.
[6]沈永明,劉詠梅,陳全站.互花米草鹽沼土壤有機質(zhì)分布特征 [J].海洋通報,2003,22(6):43-48.
[7]Netto S A,Lana P C.The role of above-and below-ground components of Spartina alterniflora (Loisel)and detritus biomass in structuring macrobenthic associations of Paranaguá Bay (SE, Brazil)[J]. Hydrobiologia, 1999, 400:167-177.
[8]商栩,管衛(wèi)兵,張國森,等.互花米草入侵對河口鹽沼濕地食物網(wǎng)的影響 [J].海洋學(xué)報,2009,31(1):132-141.
[9]全為民.長江口鹽沼濕地食物網(wǎng)的初步研究:穩(wěn)定同位素分析[D].上海:復(fù)旦大學(xué),2007.
[10]Blanchard G F,Guarini J M,Dang C,et al.Characterizing and quantifying photoinhibition in intertidal microphytobenthos[J].Journal of Phycology,2004,40(4):692-696.
[11]Bunn S E,Loneragan N R,Kempster M A.Effects of acid washing on stable isotope ratios of C and N in penaeid shrimp and seagrass:Implications for food-web studies using multiple stable isotopes[J].Limnology and Oceanography,1995,40(3):622-625.
[12]Bosley K L,Wainright S C.Effects of preservatives and acidification on the stable isotope ratios(15N:14N13C:12C)of two species of marine animals [J].Canadian Journal of Fishery Aquatic Science,1999,56(11):2181-2185.
[13]Benstead J P,March J G,F(xiàn)ry B,et al.Testing IsoSource:stable isotope analysis of a trophical fishery with diverse organic matter sources[J].Ecology,2006,87(2):326-333.
[14]Phillips D L,Gregg J W.Source partitioning using stable isotopes:coping with too many sources [J].Oecologia,2003,136(2):261-269.
[15]Teal J M.Energy flow in the salt marsh ecosystem of Georgia[J].Ecology,1962,43(4):614-624.
[16]Haines E B.The origins of detritus in Georgia salt marsh estuaries[J].Oikos,1977,29(2):254-260.
[17]Paterson A W,Whitfield A K.A stable carbon isotope study of the food-web in a freshwater-deprived South African estuary,with particular emphasis on the ichthyofauna [J].Estuarine Coastal and Shelf Science,1997,45(6):705-715.
[18]Bouillon S,Koedam N,Raman A V,et al.Primary producers sustaining macro-invertebrate communities in intertidal mangrove forests[J].Oecologia,2002,130(3):441-448.
[19]Levin L A,Neira C,Grosholz E D.Invasive cordgrass modifies wetland trophic function [J].Ecology,2006,87(2):419-432.
[20]Kreeger D A,Newell R I E.Trophic complexity between producers and invertebrate consumers in salt marshes[C] //Weinstein M P,Kreeger D A.Concepts and Controversies in Tidal Marsh Ecology.Dordrecht:Kluwer Academic Publishers,2000:187-220.
[21]Buffan-Dubau E,Carman K R.Diel feeding behavior of meiofauna and their relationships with microalgal resources[J].Limnol Oceanogr,2000,45(2):381-395.
[22]Moens T,Luyten C,Middelburg J J,et al.Tracing organic matter sources of estuarine tidal flat nematodes with stable carbon isotopes [J].MarEcolProgSer,2002,234:127-137.
[23]Wainright SC, WeinsteinM P, AbleK W.Relative importance ofbenthic microalgae,phytoplankton and the detritus of smooth cordgrass Spartina alterniflora and the common reed Phragmites australis to brackish-mash food webs[J].Marine Ecology Progress Series,2000,200:77-91.
[24]Boschker H T S,de Brouwer J F C,Cappenberg T E.The contribution of macrophyto-derived organic matter to microbial biomass in salt-marsh sediments:Stable carbon isotope analysis of microbial biomarkers [J].Limnol Oceanogr,1999,44:309-319.
[25]Hedge P,Kriwoken L K.Evidence for the effects of Spartina anglica invasion on benthic macrofauna in Little Swanport estuary,Tasmania [J].AustEcol, 2000, 25(2):150-159.
[26]Simenstad C A,Thorn R M.Spartina alterniflora(smooth cordgrass)as an invasive halophyte in Pacific Northwest Estuaries [J].Hortus Northwest:A Pacific Northwest Native Plant Directory & Journal,1995(6):9-12.
猜你喜歡
今日農(nóng)業(yè)(2021年11期)2021-08-13 08:53:34
中國生殖健康(2019年8期)2019-01-07 01:18:24
幸福(2018年33期)2018-12-05 05:22:46
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
作文周刊·小學(xué)一年級版(2017年17期)2017-06-27 19:32:55
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
廣東第二課堂·小學(xué)(2017年2期)2017-02-20 15:10:08
爆笑show(2016年7期)2017-02-09 09:36:13
海峽姐妹(2016年5期)2016-02-27 15:20:20
