皮膚衰老過程中TGF-β1,2,3的表達(dá)差異
2014-07-25 06:54:38王露萍陳煒余丕軍石瀟
組織工程與重建外科雜志 2014年3期
王露萍 陳煒 余丕軍 石瀟
皮膚老化過程復(fù)雜,受許多細(xì)胞因子影響,轉(zhuǎn)化生長因子β(TGF-β)可能在其中起著重要的作用。TGF-β是一類多功能細(xì)胞因子,其3種異構(gòu)體的生物學(xué)功能不盡相同。與TGF-β2、3相比,TGF-β1與創(chuàng)傷修復(fù)的關(guān)系更為密切,對(duì)成纖維細(xì)胞及其他間葉組織來源細(xì)胞的形態(tài)發(fā)生、增殖和分化過程起著重要作用[1];TGF-β2能夠誘導(dǎo)胚胎發(fā)育過程中中胚層的形成,促進(jìn)毛囊等皮膚輔助結(jié)構(gòu)的形成[2];TGF-β3對(duì)角質(zhì)細(xì)胞和內(nèi)皮細(xì)胞的增殖具有強(qiáng)烈的抑制作用[3]。本實(shí)驗(yàn)旨在探討TGF-β1,2,3在不同年齡SD大鼠皮膚組織中表達(dá)的變化,及其可能的生物學(xué)意義,深入揭示TGF-β與皮膚衰老過程的關(guān)系和內(nèi)在規(guī)律,為進(jìn)一步研究人類皮膚衰老機(jī)制提供相關(guān)動(dòng)物實(shí)驗(yàn)依據(jù)。
1 材料和方法
1.1 實(shí)驗(yàn)動(dòng)物及材料
采用清潔級(jí)成年雌性SD大鼠(上海斯萊克實(shí)驗(yàn)動(dòng)物中心)12只。其中,2月齡6只(青年組),18月齡6只(老年組),取頸背部4 cm×4 cm大小皮膚組織,備用。
TGF-β1抗體SC-146、TGF-β2抗體SC-90、TGF-β3抗體SC-82(Santa Cruz公司,美國);hrp標(biāo)記的山羊抗兔IgG(Jackson Immunology公司,美國)
1.2 實(shí)驗(yàn)方法
采用Western blot檢測兩組大鼠皮膚樣本中TGF-β1,2,3的蛋白表達(dá)情況。
組織塊勻漿后PBS沖洗,重懸洗滌離心,收獲細(xì)胞。細(xì)胞沉淀加入RIPA裂解液和PMSF蛋白酶抑制劑混合液(100∶1)裂解1 h,12 000 r/min,4℃離心10 min,取上清液,加入上樣緩沖液,95℃水浴10 min,-80℃保存。比色法測定蛋白濃度,定量上樣,進(jìn)行SDS-PAGE電泳1.5~2.0 h。轉(zhuǎn)膜后加一抗稀釋液(TGF-β1抗體SC-146,TGF-β2抗體SC-90,TGF-β3抗體SC-82),稀釋濃度1∶5 000,4℃過夜。第2天PBS-T洗膜后加入稀釋的二抗,hrp標(biāo)記的山羊抗兔IgG(1∶10 000),37℃孵育1 h,PBS-T洗膜,ECL化學(xué)發(fā)光法檢測顯影。底片通過凝膠灰度分析軟件Image J進(jìn)行灰度值定量。
1.3 統(tǒng)計(jì)學(xué)處理
實(shí)驗(yàn)所得數(shù)據(jù)均采用SPSS17.0軟件進(jìn)行方差齊性和正態(tài)分布檢驗(yàn),并通過單因素ANOVA分析比較均值。結(jié)果以均值±標(biāo)準(zhǔn)差表示,P0.01為差異具有統(tǒng)計(jì)學(xué)意義。
2 結(jié)果
TGF-β1在青年組大鼠皮膚中呈強(qiáng)陽性表達(dá)(Western blot灰度值相對(duì)定量1.91±0.67),而在衰老組大鼠的皮膚組織呈陰性表達(dá)(Western blot灰度值相對(duì)定量0.14±0.09),兩者比較差異有統(tǒng)計(jì)學(xué)意義(P=0.0001),說明TGF-β1在皮膚衰老過程中的表達(dá)存在顯著差異。
TGF-β2在青年組Western blot灰度值相對(duì)定量為0.16±0.07,衰老組該值為0.1±0.04,均呈陰性表達(dá)(P=0.17)。
TGF-β3在青年組Western blot灰度值相對(duì)定量為0.95±0.20,衰老組該值為0.80±0.26,均呈陽性表達(dá)(P=0.26)(圖1-4)。

圖1 TGF-β1蛋白電泳(Western blot)Fig.1Expression of TGF-β1 by western blot

圖2 TGF-β2蛋白電泳(Western blot)Fig.2Expression of TGF-β2 by western blot
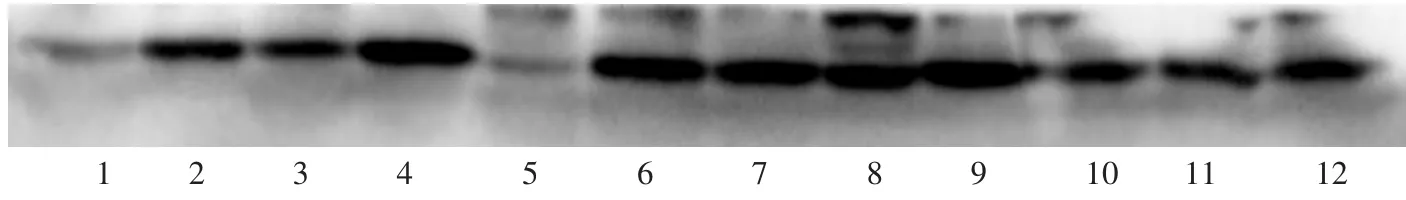
圖3 TGF-β3蛋白電泳(Western blot)Fig.3Expression of TGF-β3 by western blot

圖4 TGF-β1,2,3蛋白相對(duì)定量Fig.4Relative quantification of TGF-β1,2,3
3 討論
皮膚衰老是一個(gè)復(fù)雜的、多因素作用引起的一系列功能與美學(xué)改變的過程,與遺傳、免疫、自由基、成纖維細(xì)胞、細(xì)胞外基質(zhì)、細(xì)胞因子等諸多因素有關(guān)。老化皮膚的組織學(xué)改變主要表現(xiàn)為膠原蛋白減少和彈性纖維片段增加;分子學(xué)改變主要表現(xiàn)為表皮增生和分化性標(biāo)志表達(dá)異常[4]。成纖維細(xì)胞作為真皮中的主體細(xì)胞成分,在衰老的過程中發(fā)揮了核心作用[5-6]。其主要生物學(xué)功能是分泌膠原纖維、彈性纖維及各種基質(zhì)成分,如硫酸軟骨素及透明質(zhì)酸等,和各種蛋白質(zhì)的復(fù)合物,如蛋白多糖。這些共同構(gòu)成了皮膚充盈、承受拉力、保持光滑有彈性的物質(zhì)基礎(chǔ)[7]。衰老皮膚的成纖維細(xì)胞數(shù)量逐漸減少、合成膠原蛋白和彈性蛋白的能力下降,且Ⅰ型膠原和Ⅲ型膠原的比例變小,膠原纖維變粗,出現(xiàn)異常交鏈。同時(shí)分泌基質(zhì)降解蛋白酶(MMPs)增加,MMPs能特異性降解幾乎所有的細(xì)胞外基質(zhì)成分,使膠原纖維減少、彈性纖維斷裂紊亂,皮膚出現(xiàn)皺紋[8]。
研究表明,TGF-β三種異構(gòu)體在皮膚表皮層、真皮層以及毛囊和汗腺細(xì)胞內(nèi)都有表達(dá)[10]。與TGF-β2,3相比,TGF-β1作為成纖維細(xì)胞的主要趨化因子,與創(chuàng)傷修復(fù)、皮膚老化的關(guān)系顯得尤為密切;TGF-β2隨著毛囊的發(fā)育成熟,其在毛囊細(xì)胞內(nèi)表達(dá)顯著增強(qiáng),而TGF-β3則在角質(zhì)細(xì)胞和血管內(nèi)皮細(xì)胞內(nèi)表達(dá)明顯。
TGF-β1對(duì)成纖維細(xì)胞及其他間葉組織來源的細(xì)胞有明顯的刺激作用,主要分布于表層基底細(xì)胞及胞外基質(zhì)中,可以通過增加基質(zhì)蛋白的合成,如膠原蛋白、糖蛋白的表達(dá)和分泌,加強(qiáng)細(xì)胞黏附蛋白受體的轉(zhuǎn)錄、翻譯和處理過程;并可減少M(fèi)MPs的合成,對(duì)細(xì)胞的形態(tài)發(fā)生、增殖和分化過程起著重要作用,有利于胚胎發(fā)育和細(xì)胞修復(fù)、組織修復(fù)[9]。本實(shí)驗(yàn)證實(shí),TGF-β1在不同年齡的SD大鼠皮膚組織中表達(dá)有顯著差異,在青年組大鼠皮膚中TGF-β1蛋白表達(dá)強(qiáng),而在衰老組大鼠的皮膚組織中呈陰性表達(dá),提示皮膚的生長發(fā)育、表型和功能的維持可能存在TGF-β1介導(dǎo)的信號(hào)通路參與,并且內(nèi)源性TGF-β1可能是皮膚修復(fù)與再生的重要內(nèi)源性調(diào)控因子。
TGF-β2主要分布于汗腺細(xì)胞及毛囊細(xì)胞中,能夠誘導(dǎo)胚胎發(fā)育過程中中胚層的形成,促進(jìn)毛囊等皮膚輔助結(jié)構(gòu)的形成[10],在本實(shí)驗(yàn)中,各年齡組大鼠的皮膚組織中TGF-β2含量極少,年齡差異不明顯,提示TGF-β2在皮膚組織中的含量極微,對(duì)皮膚的老化產(chǎn)生的影響可能較TGF-β1、3小。
有研究采用MTT比色法檢測TGF-β3對(duì)瘢痕組織成纖維細(xì)胞增殖活性的影響,并以3H-脯氨酸摻入法檢測細(xì)胞內(nèi)膠原合成及TGF-β1蛋白表達(dá)情況。結(jié)果證實(shí),TGF-β3可抑制成纖維細(xì)胞的增殖,下調(diào)TGF-β1蛋白及Ⅰ型膠原蛋白表達(dá)[11]。本實(shí)驗(yàn)中,TGF-β3在不同年齡的SD大鼠皮膚組織中有著相對(duì)穩(wěn)定的表達(dá),年輕組表達(dá)少于TGF-β1卻明顯多于TGF-β2,而老年組表達(dá)均高于另外兩種蛋白,提示TGF-β3可能在抑制成纖維細(xì)胞增殖,下調(diào)TGF-β1蛋白及Ⅰ型膠原蛋白表達(dá)方面對(duì)皮膚的衰老有一定的促進(jìn)作用。
因此,我們可以認(rèn)為在三種TGF-β蛋白中,TGF-β1表達(dá)的改變可能在皮膚衰老過程中起到主要作用,而TGF-β3通過何種機(jī)制對(duì)皮膚衰老產(chǎn)生影響還有待進(jìn)一步的研究證實(shí)。
[1]Gressner AM,Weiskirchen R,Breitkopf K,et al.Roles of TGF-β in hepatic fibrosis[J].Front Biosci,2002,7:d793-d807.
[2]Foitzik K,Paus R,Doetschman T,et al.The TGF-beta2 isoform is both a required and sufficient inducer of murine hair follicle morphogenesis[J].Dev Biol,1999,212(2):278-289.
[3]Merwin JR,Newman W,Beall LD,et al.Vascular cells respond differentially to transforming growth factors beta1 and beta2[J].Am J Pathol,1991,138(1):37-51.
[4]Ten Dijke P,Goumans MJ,Itoh F,et al.Regulation of cell proliferation by Smad proteins[J].Cell Physio,2002,191(1):1-16.
[5]Verreechia F,Mauviel A.Transforming growth factor-beta signaling through the Smad pathway:role in extracelular matrix gene expression and regulation[J].J Invest Dermatol,2002,118(2):211-215.
[6]Chung JH,Seo JY,Choi HR,et a1.Modulation of skin collagen metabolism in aged and photoaged human skin in vivo[J].J Invest Dermatol,2001,117(5):1218-1224.
[7]吳國平,周燕,譚文松,等.人皮膚成纖維細(xì)胞在二維和三維培養(yǎng)系統(tǒng)中的生長代謝特性[J].中國組織工程研究與臨床康復(fù),2007,11(1):74-77.
[8]關(guān)英杰,金錫鵬.皮膚衰老研究進(jìn)展[J].中國老年學(xué)雜志,2003,23:396-397.
[9]Acharya PS,Majumdar S,Jacob M,et al.Fibroblast migration is mediated by CD44-dependent TGF beta activation[J].J Cell Sci,2008,121(9):1393-1402.
[10]陳偉,付小兵,孫同柱,等.胎兒和成人皮膚組織中TGF-β1,2,3因子蛋白含量的變化及其與創(chuàng)面無瘢愈合的關(guān)系[J].感染.炎癥.修復(fù),2001,2(4):204-206.
[11]黃輝,唐勝建,鮑衛(wèi)漢.α-平滑肌肌動(dòng)蛋白在瘢痕成纖維細(xì)胞中的表達(dá)[J].中國修復(fù)重建外科雜志,2001,15(6):347-350.
