人肺動脈平滑肌細(xì)胞中微小RNA-210通過MKP-1負(fù)性調(diào)節(jié)低氧下細(xì)胞的增殖*
2014-08-13 12:42:48靳有鵬龐婷婷王玉林
中國病理生理雜志 2014年11期
關(guān)鍵詞:研究
靳有鵬, 龐婷婷, 王 偉, 王玉林
(山東大學(xué)附屬省立醫(yī)院兒科, 山東 濟(jì)南 250021)
慢性低氧性肺動脈高壓是臨床許多心肺疾病發(fā)生發(fā)展過程中伴隨或最終的病理生理環(huán)節(jié),是一種嚴(yán)重的甚至危及生命的肺血管性疾病,臨床上表現(xiàn)為肺動脈壓力增高、肺血管阻力增加,右心室阻力負(fù)荷過重最終致右心衰竭,組織病理學(xué)改變?yōu)槔奂叭珜拥难苎祝鼙诘乃谐煞旨?xì)胞均受累,最終肺血管及右室重塑。
微小RNA(microRNA,miRNA)是廣泛存在于真核生物中的一類長度為20~24個核苷酸所構(gòu)成的內(nèi)源性非編碼調(diào)控單鏈小分子RNA,它是由含有莖環(huán)結(jié)構(gòu)的miRNA前體經(jīng)Dicer剪切而成。越來越多的研究證實,miRNA作為重要的轉(zhuǎn)錄后調(diào)節(jié)基因,通過抑制靶信使mRNA 的翻譯而起到重要的調(diào)控作用,參與細(xì)胞的分化、增殖、凋亡以及多種生物組織的發(fā)育調(diào)節(jié)過程[1]。研究表明哺乳動物中50%~60%的mRNA受miRNA調(diào)控[2]。
近年來的研究表明,miRNA在低氧下細(xì)胞進(jìn)行生理性及病理性調(diào)節(jié)過程中發(fā)揮著舉足輕重的作用。其中一些miRNA的表達(dá)顯著地受低氧調(diào)節(jié),因此被命名為“低氧相關(guān)的微小RNA(hypoxia-related miRNA)”[3]。MicroRNA-210(miR-210)是一種在低氧下表達(dá)顯著上調(diào)的miRNA,它參與調(diào)節(jié)血管生成、細(xì)胞凋亡、增殖、分化、DNA修復(fù)、線粒體代謝以及腫瘤生長[4-7],其在肺動脈高壓中的作用日益受到重視,有研究發(fā)現(xiàn)在慢性低氧所致肺動脈高壓鼠的肺組織中miR-210表達(dá)上調(diào)[8], 但其在低氧性肺動脈高壓中的作用仍不甚清楚。
絲裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)細(xì)胞內(nèi)信號轉(zhuǎn)導(dǎo)途徑廣泛存在。真核細(xì)胞中是由4種激酶組成的鏈?zhǔn)椒磻?yīng),其中,每種激酶都通過復(fù)雜的反應(yīng)網(wǎng)絡(luò)激活下游底物激酶,調(diào)節(jié)細(xì)胞生長、增殖、分化或凋亡,被廣泛用于細(xì)胞信號轉(zhuǎn)導(dǎo)的研究。它的級聯(lián)反應(yīng)實際上是分3步進(jìn)行的:首先是MAPKK激酶(MAPKK kinase,MKKK)磷酸化激活MAPK激酶(MAPK kinase,MKK),后者活化后磷酸化激活MAPK,活化的MAPK再去激活一些轉(zhuǎn)錄因子,調(diào)控基因表達(dá)。活化的MAPK主要被其磷酸酶去磷酸化而失活,這其中絲裂原活化蛋白激酶磷酸酶1(mitogen-activated protein kinase phosphatase 1,MKP-1)的作用最為重要。研究發(fā)現(xiàn), MKP-1可被低氧誘導(dǎo)[9],近年來,越來越多的研究發(fā)現(xiàn)其在肺動脈高壓中起著重要作用。一項研究發(fā)現(xiàn)MKP-1缺失的小鼠易發(fā)展為重度肺動脈高壓[10],也有研究證實西地那非通過誘導(dǎo)MKP-1在肺動脈平滑肌細(xì)胞中發(fā)揮細(xì)胞保護(hù)作用[11]。但關(guān)于miR-210通過調(diào)節(jié)MKP-1而參與肺動脈高壓的相關(guān)研究,至今尚未見報道。
本文旨在研究低氧時人肺動脈平滑肌細(xì)胞(human pulmonary artery smooth muscle cells, hPASMCs)miR-210和MKP-1的表達(dá)情況、它們之間的相互關(guān)系以及對肺動脈平滑肌細(xì)胞增殖的影響,為治療低氧性肺動脈高壓提供新的思路。
材 料 和 方 法
1 主要試劑
人肺動脈平滑肌細(xì)胞購于Lonza;RNA提取試劑Trizol和轉(zhuǎn)染試劑購自Invitrogen;實時熒光定量PCR試劑盒、引物、miRNA抑制劑及增強(qiáng)劑均購自生命技術(shù)公司。
2 細(xì)胞培養(yǎng)和分組
將hPASMCs置于含4.9% FBS及多種生長因子的培養(yǎng)基中,當(dāng)細(xì)胞貼壁生長至80%~90%融合時進(jìn)行傳代或用于實驗分組接種,取4~8代細(xì)胞用于實驗。每次實驗前取狀態(tài)良好的對數(shù)生長期細(xì)胞,經(jīng)含0.02% EDTA/0.25%胰蛋白酶消化液消化,形成細(xì)胞懸液后分組接種,細(xì)胞于貼壁24 h后,換無血清培養(yǎng)基24 h使細(xì)胞生長同步化,再分別處理各實驗組。實驗分組:共分成12組,常氧處理(21% O2、5% CO2)6組和低氧處理(1% O2、5%CO2)6組,見表1。
3 主要方法
3.1實時熒光定量PCR法檢測各組平滑肌細(xì)胞中miR-210和MKP-1 mRNA的表達(dá) RNA提取試劑和轉(zhuǎn)染試劑均購自Invitrogen。用Trizol試劑提取細(xì)胞總RNA并測定其濃度,逆轉(zhuǎn)錄成cDNA,再按說明書進(jìn)行實時熒光定量PCR。以RNU48作為小RNA含量的內(nèi)參照。PCR反應(yīng)條件:95 ℃ 15 min熱啟動,然后94 ℃ 15 s,60 ℃ 30 s、72 ℃ 30 s,共40個循環(huán)。最后通過解鏈曲線分析擴(kuò)增產(chǎn)物的特異性。 miRNA的相對表達(dá)量用2-ΔΔCt方法來表示。
3.2用Western blotting法比較各組MKP-1蛋白的表達(dá) 提取細(xì)胞總蛋白,取20 μg細(xì)胞總蛋白進(jìn)行聚丙烯酰胺凝膠電泳,按濕轉(zhuǎn)法將電泳產(chǎn)物轉(zhuǎn)移到PVDF膜,5%脫脂奶粉4 ℃封閉2 h,滴加Ⅰ抗(1∶1 000)4 ℃過夜,TBST洗膜,滴加Ⅱ抗(1∶1 000)室溫孵育2 h,TBST洗膜后顯色,并用β-actin作為內(nèi)參照。
3.3MTT法檢測肺動脈平滑肌細(xì)胞的增殖 培養(yǎng)細(xì)胞中加入20 μL MTT溶液,繼續(xù)37 ℃孵育4 h后,吸棄培養(yǎng)上清液,加入DMSO 150 μL,振蕩10 min。在ELISA檢測儀上測定各孔的A490值。
4 統(tǒng)計學(xué)處理
數(shù)據(jù)以均數(shù)±標(biāo)準(zhǔn)誤 (mean±SEM) 來表示,兩組間比較采用t檢驗,多組間比較采用方差分析,用SPSS 11.0統(tǒng)計軟件處理,以P<0.05為差異有統(tǒng)計學(xué)意義。
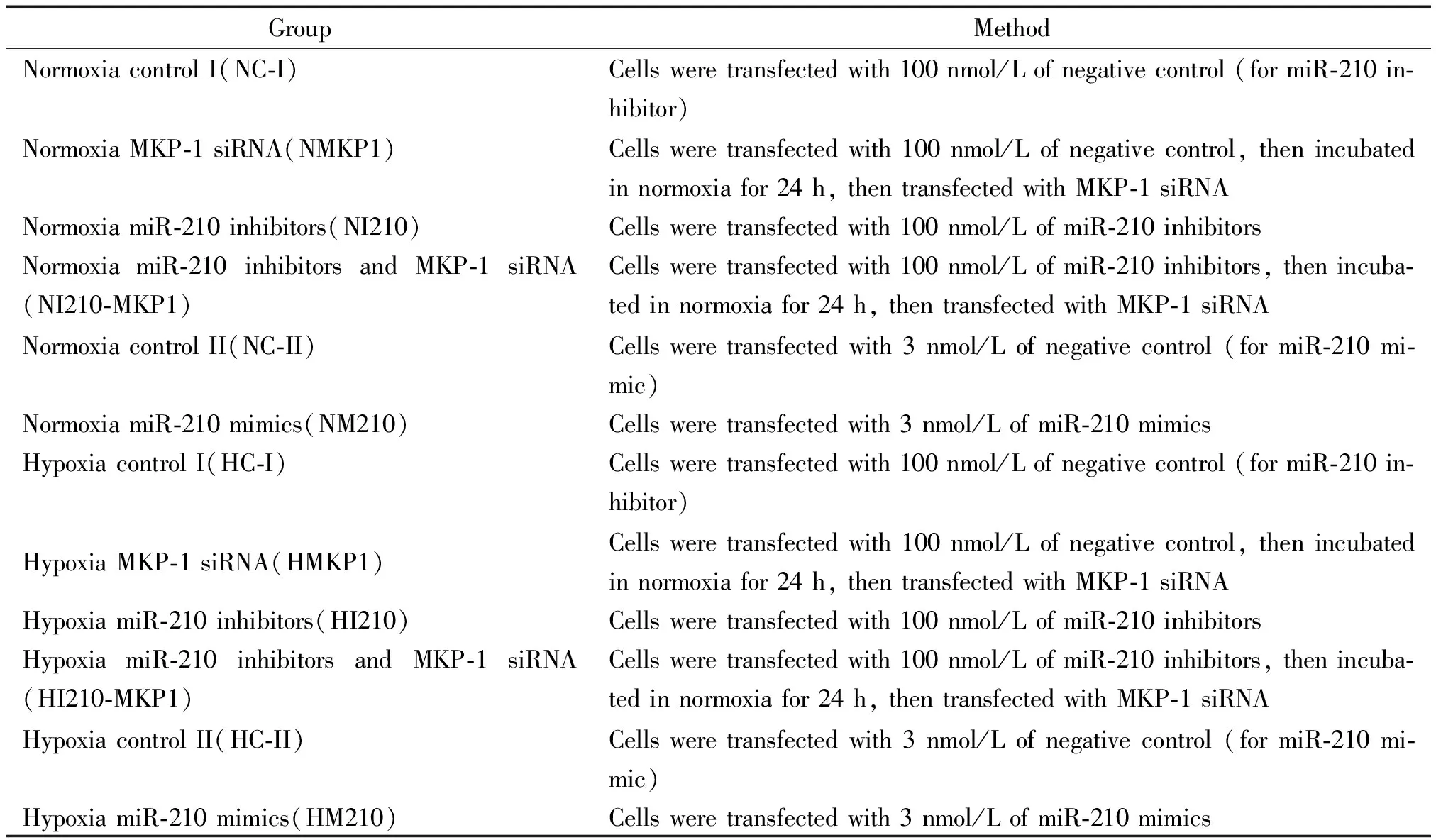
表1 細(xì)胞培養(yǎng)和實驗分組
結(jié) 果
1 低氧時hPASMCs miR-210的表達(dá)
hPASMCs分別放置于21% O2或1% O2的培養(yǎng)箱中48 h后,發(fā)現(xiàn)miR-210的表達(dá)水平明顯增加,見圖1。
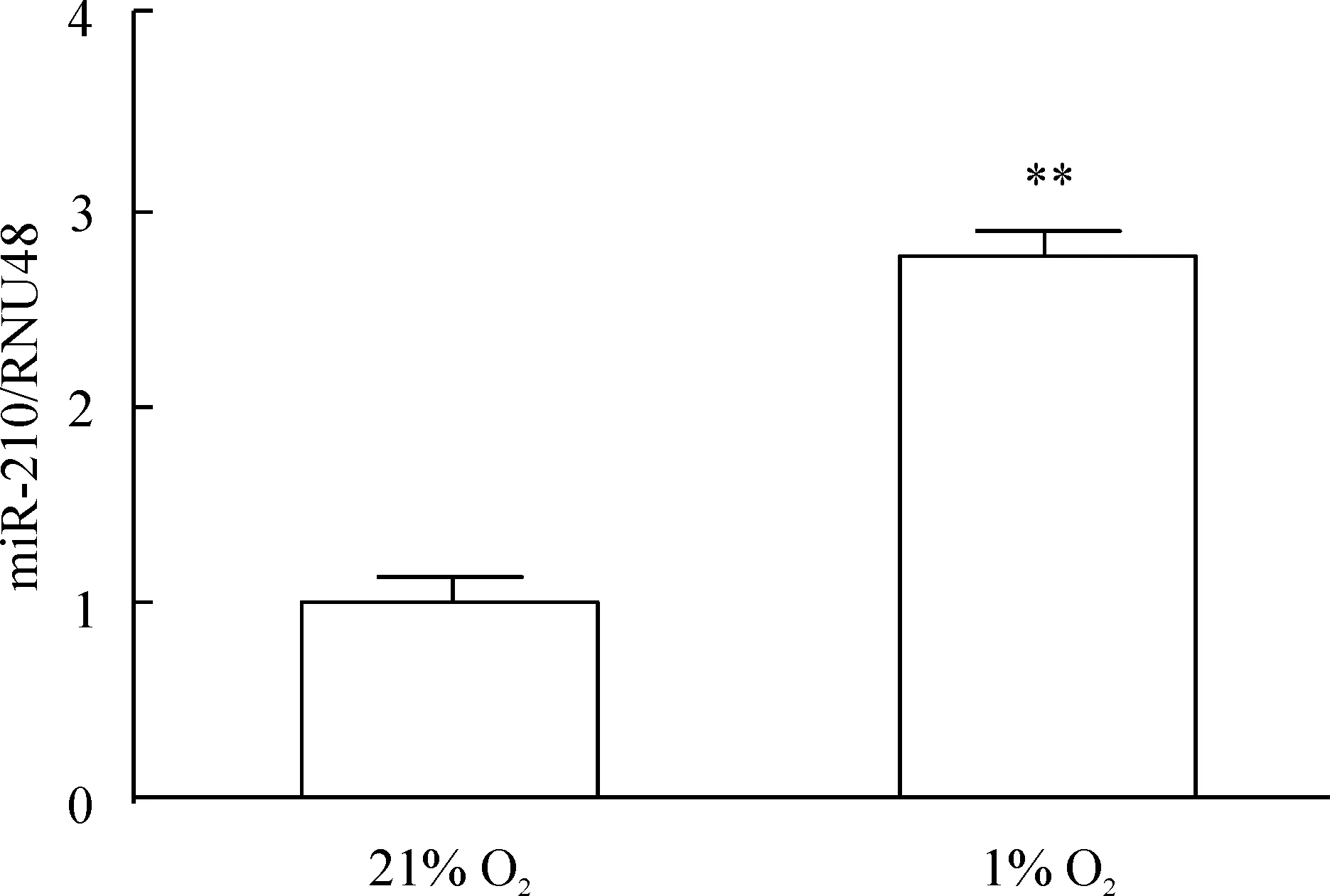
Figure 1. The expression of miR-210 in the hPASMCs determined by quantitative real-time PCR analysis.The hPASMCs were exposed to 21% O2 (normoxia) or 1% O2 (hypoxia) for 48 h. miR-210 mRNA levels were normalized to RNU48 expression.Mean±SEM.n=3. **P<0.01 vs 21% O2.
2 低氧時hPASMCs MKP-1的表達(dá)
hPASMCs分別放置于21% O2或1% O2的培養(yǎng)箱中48 h后,發(fā)現(xiàn)MKP-1 mRNA和蛋白的表達(dá)明顯增加,見圖2。
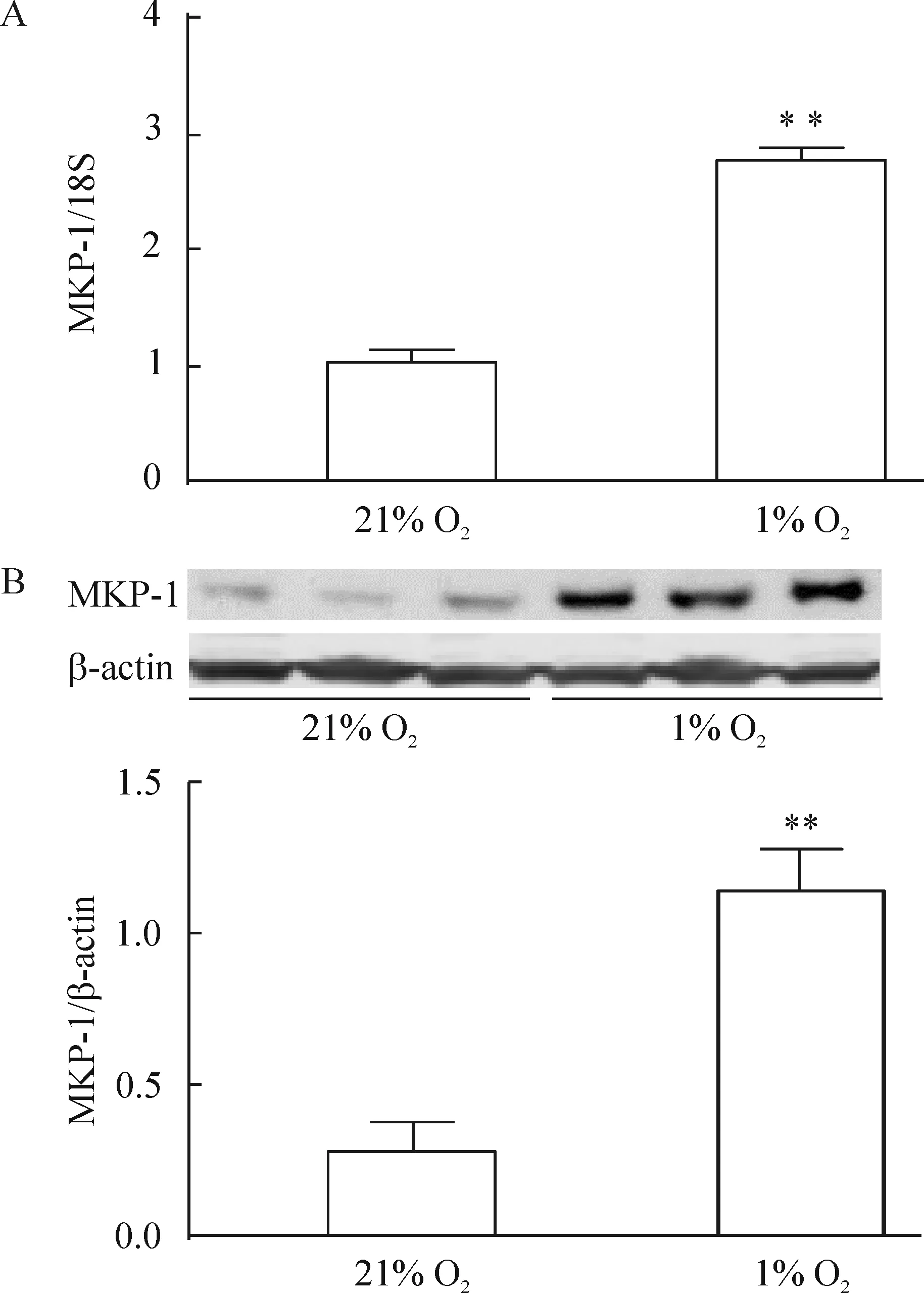
Figure 2. Expression of MKP-1 mRNA (A) and protein (B) in the hPASMCs. The hPASMCs were exposed to 21% O2 (normoxia) or 1% O2 (hypoxia) for 48 h.Mean±SEM.n=3.**P<0.01 vs 21% O2.
3 抑制miR-210表達(dá)使MKP-1的表達(dá)增加并可抑制低氧誘導(dǎo)的細(xì)胞增殖
將miR-210抑制劑轉(zhuǎn)染入hPASMCs內(nèi),再分別放置于21% O2或1% O2的培養(yǎng)箱中48 h后,發(fā)現(xiàn)常氧及低氧下,miR-210抑制劑明顯抑制miR-210的表達(dá),增加MKP-1的表達(dá),但只有低氧下抑制miR-210的表達(dá)后hPASMCs的增殖受到抑制,常氧下卻沒有這種效應(yīng),見圖3。
4 miR-210過表達(dá)可抑制低氧誘導(dǎo)的MKP-1表達(dá)上調(diào)但不影響細(xì)胞增殖
將miR-210增強(qiáng)劑轉(zhuǎn)染入hPASMCs內(nèi),再分別放置于21% O2或1% O2的培養(yǎng)箱中48 h后,實驗結(jié)果表明,miR-210過表達(dá)明顯抑制低氧誘導(dǎo)的MKP-1表達(dá)上調(diào),但常氧及低氧均未影響hPASMCs的增殖,見表2、圖4。
5 MKP-1基因沉默后,低氧下miR-210抑制劑對細(xì)胞增殖的抑制作用消失
再將miR-210抑制劑轉(zhuǎn)染入hPASMCs內(nèi),再分別放置于1% O2培養(yǎng)箱中48 h后,實驗結(jié)果表明MKP-1基因沉默后,低氧下miR-210抑制劑對細(xì)胞增殖的抑制作用消失,見圖5。
討 論
2007年首次報道了通過微距陣的方法發(fā)現(xiàn)了低氧下miR-210的表達(dá)會上調(diào),該作用可被低氧誘導(dǎo)因子 1α(hypoxia-inducible factor 1α,HIF-1α)或低氧誘導(dǎo)因子 2α(hypoxia-inducible factor 2α,HIF-2α)所誘導(dǎo)。本研究發(fā)現(xiàn)將hPASMCs置于低氧培養(yǎng)箱中48 h,miR-210的表達(dá)升高2.5倍左右,說明hPASMCs中miR-210的表達(dá)可被低氧所誘導(dǎo)。有研究發(fā)現(xiàn),在急性缺氧的情況下,低氧誘導(dǎo)的miR-210通過下調(diào)鐵硫蛋白1/2[12]或線粒體電子傳遞蛋白[13-14]的表達(dá)來保障細(xì)胞存活,然而,另一方面這又是引起慢性缺氧性疾病的病理生理基礎(chǔ)。
MKP-1屬于絲裂原活化蛋白激酶家族,是細(xì)胞內(nèi)的一種絲氨酸/蘇氨酸蛋白激酶。MKP-1的表達(dá)在多種因素刺激下均會增加,包括低氧、過氧化、糖皮質(zhì)激素升高、熱休克等。本研究同樣發(fā)現(xiàn)低氧下的hPASMCs中MKP-1的表達(dá)增加。在多種細(xì)胞生理反應(yīng)(如生長因子信號調(diào)節(jié)通路、細(xì)胞的炎癥、分化及凋亡等)中,MKP-1都通過使絲裂原活化蛋白激酶去磷酸化而提供重要的負(fù)反饋調(diào)節(jié)機(jī)制。類似miR-210,在低氧狀態(tài)下MKP-1也發(fā)揮著很多作用。研究表明MKP-1基因敲除的小鼠易表現(xiàn)為過度的炎癥反應(yīng),易患自身免疫性疾病及代謝缺陷病[15-16]。
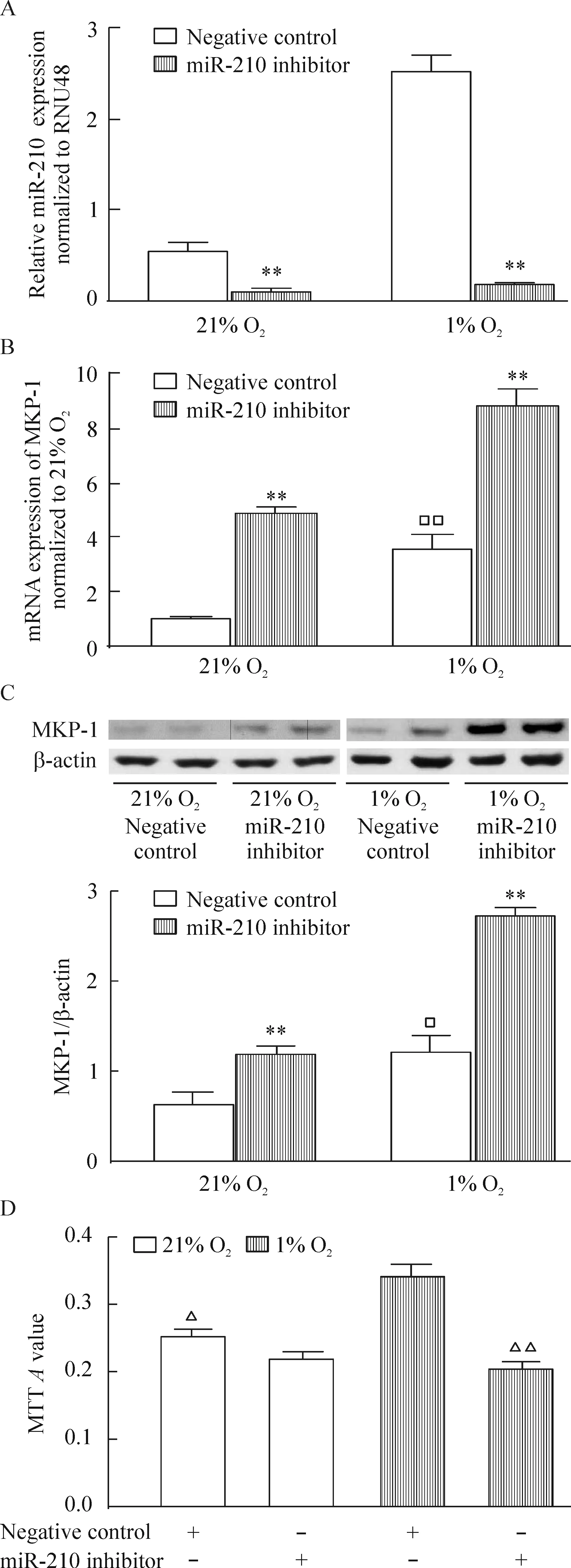
Figure 3. The effects of miR-210 inhibitor on hypoxia-induced MKP-1 expression and proliferation in the hPASMCs. The hPASMCs were exposed to 21% O2 (normoxia) or 1% O2 (hypoxia) for 48 h. A: miR-210 expression; B,C: MKP-1 mRNA and protein expression; D: proliferation of hPASMCs. Mean±SEM.n=3.**P<0.01 vs negative control;□P<0.05,□□P<0.01 vs 21% O2;△P<0.05,△△P<0.01 vs 1% O2+negative control.

表2 miR-210增強(qiáng)劑對miR-210表達(dá)的影響
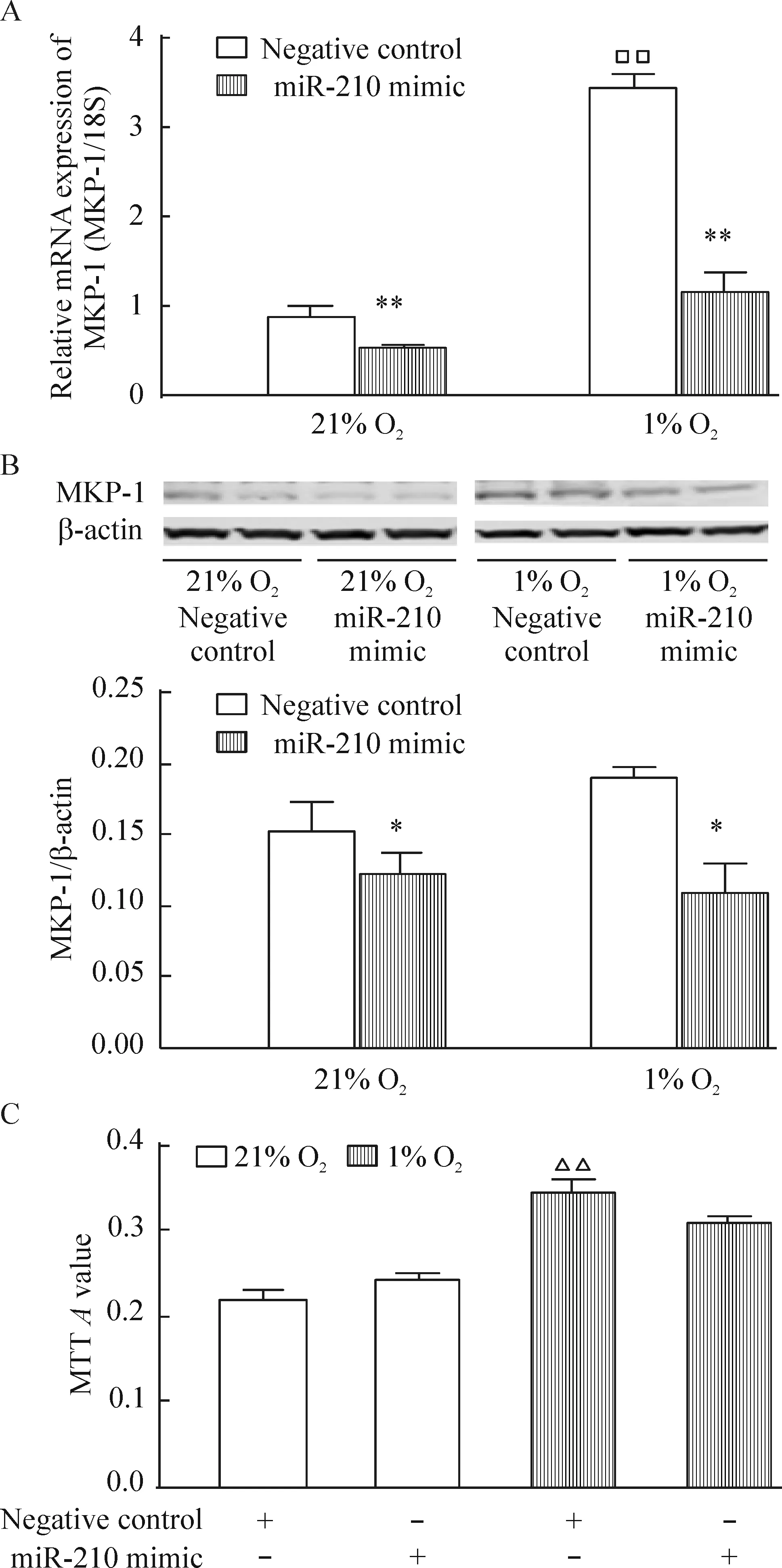
Figure 4. The effects of miR-210 mimic on MKP-1 expression (A,B) and proliferation (C) in the hPASMCs.The hPASMCs were exposed to 21%O2 (normoxia) or 1%O2 (hypoxia)for 48 h. Mean±SEM.n=3.*P<0.05,**P<0.01 vs negative control;□□P<0.01 vs 21% O2; △△P<0.01 vs 21% O2 negative control.
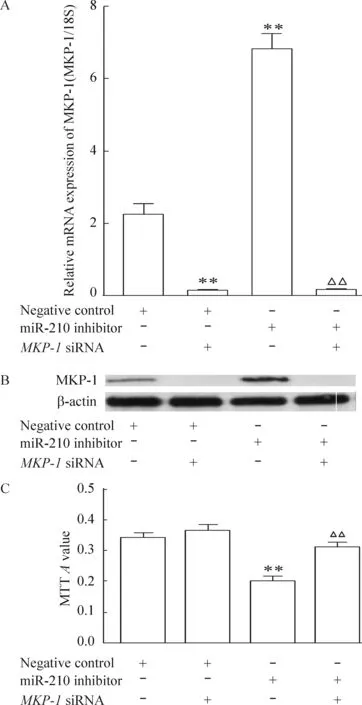
Figure 5. The effects of MKP-1 knockdown (A,B) on miR-210 inhibitor-decreased hPASMC proliferation (C) under hypoxia.Mean±SEM.n=3.**P<0.01 vs negative control; △△P<0.01 vs miR-210 inhibitor.
另有一項研究發(fā)現(xiàn)MKP-1缺失的小鼠在低氧下易患更嚴(yán)重的肺動脈高壓,提示MKP-1在低氧性肺高壓中發(fā)揮著重要作用[10]。
雖然miR-210和MKP-1在低氧性肺動脈高壓的發(fā)生發(fā)展中均起著舉足輕重的作用,但它們之間的相互關(guān)系仍不甚清楚。本研究結(jié)果顯示不論是低氧下或常氧下的hPASMCs中,抑制miR-210的表達(dá)后,MKP-1轉(zhuǎn)錄后水平的表達(dá)均增加,而增加miR-210的表達(dá)后,MKP-1轉(zhuǎn)錄后水平的表達(dá)均下降。由此說明,hPASMCs的MKP-1是miR-210的靶基因。通常MKP-1通過MAPK起作用,且MKP-1的作用取決于其與MKK的相對活性,關(guān)于miR-210與MKK的關(guān)系尚未見報道,miR-210是否進(jìn)一步通過調(diào)節(jié)MKK的活性來調(diào)節(jié)肺動脈平滑肌細(xì)胞的增殖尚不清楚,仍需進(jìn)一步研究來證實。
本研究還發(fā)現(xiàn)在hPASMCs中,抑制miR-210的表達(dá)可以減少低氧誘導(dǎo)的細(xì)胞凋亡,但miR-210的過表達(dá)卻并不影響細(xì)胞的增殖情況,由此說明,低氧下miR-210可負(fù)性調(diào)節(jié)hPASMCs的增殖,然而,近年來有另外一項研究報道[17],miR-210在hPASMCs中發(fā)揮抗凋亡的作用而并不影響細(xì)胞的增殖,這似與本研究結(jié)果有所不同,究其原因可能有如下兩方面。一是,這2個實驗所采用的培養(yǎng)細(xì)胞的氧濃度不同,本研究采用的氧濃度為1%,而另一研究采用的氧濃度為3%。二是,另有其它研究發(fā)現(xiàn)miRNA對靶基因呈非線性、劑量依賴性的選擇性調(diào)節(jié)作用[18],本研究中miR-210抑制劑的量與另一研究中所采用的劑量也不同,這可能也是原因之一。而且,miR-210抑制劑對細(xì)胞增殖的負(fù)性調(diào)節(jié)作用需依靠MKP-1來發(fā)揮,具體機(jī)制仍需進(jìn)一步研究。
綜上所述,本研究發(fā)現(xiàn)在低氧下的hPASMCs中,MKP-1是miR-210的一個新的靶基因,MKP-1可以介導(dǎo)miR-210抑制劑對hPASMCs增殖的負(fù)性調(diào)節(jié)作用,這有望成為治療低氧性動脈肺高壓的新的靶點。
[參 考 文 獻(xiàn)]
[1] Pushparaj PN, Aarthi JJ, Kumar SD, et al. RNAi and RNAa: the Yin and Yang of RNAome[J]. Bioinformation, 2008, 2(6): 235-237.
[2] Lodish HF, Zhou B, Liu G, et al. Micromanagement of the immune system by microRNAs[J]. Nat Rev Immunol, 2008, 8(2):120-130.
[3] Hale AE, White K, Chan SY. Hypoxamirs in pulmonary hypertension: breathing new life into pulmonary vascular research [J]. Cardiovasc Diagn Ther, 2012, 2(3):200-212.
[4] Huang X, Le QT, Giaccia AJ. MiR-210: micromanager of the hypoxia pathway [J]. Trends Mol Med, 2010, 16(5):230-237.
[5] Kulshreshtha R, Ferracin M, Wojcik S, et al. A microRNA signature of hypoxia [J]. Mol Cell Biol, 2007, 27(5):1859-1867.
[6] Fasanaro P, D’Alessandra Y, Di Stefano V, et al. MicroRNA-210 modulates endothelial cell response to hypoxia and inhibits the receptor tyrosine kinase ligand Ephrin-A3[J]. J Biol Chem, 2008, 283(23):15878-15883.
[7] Won Kim H, Haider HK, Jiang S, et al. Ischemic preconditioning augments survival of stem cells via miR-210 expression by targeting caspase-8-associated protein 2 [J]. J Biol Chem, 2009, 284(48):33161-33168.
[8] Yang S, Banerjee S, de Freitas A, et al. miR-21 regulates chronic hypoxia-induced pulmonary vascular remodeling[J]. Am J Physiol Lung Cell Mol Physiol, 2012, 302(6): L521-L529.
[9] Seta KA, Kim R, Kim HW, et al. Hypoxia-induced regulation of MAPK phosphatase-1 as identified by subtractive suppression hybridization and cDNA microarray analysis [J]. J Biol Chem, 2001, 276(48):44405-44412.
[10] Jin Y, Calvert TJ, Chen B, et al. Mice deficient in Mkp-1 develop more severe pulmonary hypertension and greater lung protein levels of arginase in response to chronic hypoxia[J]. Am J Physiol Heart Circ Physiol, 2010, 298(5): H1518-H1528.
[11] Li B, Yang L, Shen J, et al. The antiproliferative effect of sildenafil on pulmonary artery smooth muscle cells is mediated via upregulation of mitogen-activated protein kinase phosphatase-1 and degradation of extracellular signal-regulated kinase 1/2 phosphorylation [J]. Anesth Analg, 2007, 105(4): 1034-1041.
[12] Chan SY, Zhang YY, Hemann C, et al. MicroRNA-210 controls mitochondrial metabolism during hypoxia by repressing the iron-sulfur cluster assembly proteins ISCU1/2[J]. Cell Metab, 2009, 10(4):273-284.
[13] Muralimanoharan S, Maloyan A, Mele J, et al. MIR-210 modulates mitochondrial respiration in placenta with preeclampsia[J]. Placenta, 2012, 33(10): 816-823.
[14] Puisségur MP, Mazure NM, Bertero T, et al. miR-210 is overexpressed in late stages of lung cancer and mediates mitochondrial alterations associated with modulation of HIF-1 activity[J]. Cell Death Differ, 2011, 18(3): 465-478.
[15] Bennett AM, Tonks NK. Regulation of distinct stages of skeletal muscle differentiation by mitogen-activated protein kinases[J]. Science, 1997, 278(5341): 1288-1290.
[16] Liu Y, Shepherd EG, Nelin LD. MAPK phosphatases: regulating the immune response[J]. Nat Rev Immunol, 2007, 7(3):202-212.
[17] Gou D, Ramchandran R, Peng X, et al. miR-210 has an antiapoptotic effect in pulmonary artery smooth muscle cells during hypoxia[J]. Am J Physiol Lung Cell Mol Physiol, 2012, 303(8):L682-L691.
[18] Shu J, Xia Z, Li L, et al. Dose-dependent differential mRNA target selection and regulation by let-7a-7f and miR-17-92 cluster microRNAs[J]. RNA Biol, 2012, 9(v10):1275-1287.
猜你喜歡
體育科技文獻(xiàn)通報(2022年3期)2022-05-23 13:46:54
天津外國語大學(xué)學(xué)報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機(jī)設(shè)計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學(xué)報(2017年2期)2017-07-05 08:13:02
國際商務(wù)財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
