布魯菌wzt基因的原核表達(dá)及生物信息學(xué)分析
2014-09-15 04:39:14王秀然康立恒郝芳芳許春曉盧天成周曉麗
東北師大學(xué)報(bào)(自然科學(xué)版) 2014年3期
王秀然,康立恒,安 偉,郝芳芳,陳 思,許春曉,盧天成,周曉麗
(1.吉林農(nóng)業(yè)大學(xué)生命科學(xué)學(xué)院,吉林 長(zhǎng)春 130118;2.吉林大學(xué)白求恩醫(yī)學(xué)部,吉林 長(zhǎng)春 130021;3.吉林大學(xué)第一醫(yī)院,吉林 長(zhǎng)春 130021;4.軍事醫(yī)學(xué)科學(xué)院軍事獸醫(yī)研究所,吉林 長(zhǎng)春 130122)
布魯菌病是一種人畜共患疾病,該病不但嚴(yán)重影響?zhàn)B殖業(yè)的發(fā)展,對(duì)人類健康也構(gòu)成了巨大威脅.人畜布魯菌病難以防控的原因是多方面的,其中,其致病機(jī)理(包括胞內(nèi)寄生機(jī)理)和免疫機(jī)理不清楚是主要原因.
引起布魯菌病的布魯菌是兼性細(xì)胞內(nèi)寄生的革蘭氏陰性球桿菌,不產(chǎn)生芽孢和莢膜.雖然布魯菌不能自主運(yùn)動(dòng),但其攜帶組裝鞭毛的所有基因,也不含趨化系統(tǒng)[1].脂多糖(lipopolysaccharide,LPS)是布魯菌主要的毒力因子,也是引起哺乳動(dòng)物免疫的主要抗原之一[2].目前布魯菌脂多糖合成的途徑并不完全清晰,已經(jīng)發(fā)現(xiàn)的與布魯菌脂多糖合成相關(guān)的關(guān)鍵基因有wbkA,gmd,wzm,wzt,wbkB,wbkC等,其中wzt基因?yàn)锳BC轉(zhuǎn)運(yùn)系統(tǒng)(ABC transporter)的重要組成部分[3],布魯菌LPS-O側(cè)鏈的合成即是通過(guò)ABC轉(zhuǎn)運(yùn)途徑.有研究顯示[4],線性O(shè)多糖大多是通過(guò)這一途徑完成的,這一過(guò)程中糖苷連接到UDP-PP-糖鏈上,在細(xì)胞膜內(nèi)膜上完成聚合物的合成,然后借助ABC轉(zhuǎn)運(yùn)蛋白運(yùn)到膜外,鏈接到核心寡糖和類脂A上;其中,wzt基因編碼ABC型轉(zhuǎn)運(yùn)體(ATP酶結(jié)構(gòu)域)O抗原輸出系統(tǒng)ATP結(jié)合蛋白,其突變導(dǎo)致B.melitensis 16M形成粗糙型突變體,ELISA檢測(cè)S-LPS的O側(cè)鏈可確定O側(cè)鏈缺失.O多糖合成前體由wzt形成的ABC轉(zhuǎn)運(yùn)系統(tǒng)通過(guò)十一異戊烯聯(lián)聚合物轉(zhuǎn)移到膜,在那里它被連接到脂質(zhì)A核心和易位到外膜的周質(zhì)面[5].ABC轉(zhuǎn)運(yùn)系統(tǒng)在細(xì)菌中是一個(gè)主要的細(xì)胞轉(zhuǎn)運(yùn)機(jī)器,由一系列同源基因編碼的操縱子組成[6].該操縱子由ATP結(jié)合蛋白、膜蛋白、亞基結(jié)合蛋白3部分組成,通常情況下,形成由6個(gè)跨膜片段組成的2個(gè)完整的膜蛋白[7].ABC轉(zhuǎn)運(yùn)系統(tǒng)是膜外生物大分子合成的主要運(yùn)輸通道之一,參與胞外多糖的合成[8].此外,ABC轉(zhuǎn)運(yùn)系統(tǒng)在細(xì)菌營(yíng)養(yǎng)成分的運(yùn)輸、獨(dú)立分子的外排等過(guò)程中也起著重要的作用[9].研究發(fā)現(xiàn),wzt基因缺失會(huì)造成布魯菌O側(cè)鏈缺失,同時(shí)降低布魯菌的毒力[10];缺失wzt基因的布魯菌突變株與親本株相比,細(xì)胞免疫水平下降,體液免疫水平上升[11-12].
本研究對(duì)源于布魯菌的wzt基因進(jìn)行了克隆并對(duì)原核進(jìn)行了表達(dá),對(duì)其進(jìn)行了生物信息學(xué)分析和結(jié)構(gòu)預(yù)測(cè),以為闡明布魯菌wzt基因的功能提供理論依據(jù).
1 材料與方法
1.1 菌株及載體
布魯菌Brucella abortus S19DNA、感受態(tài)菌株Eschericholia.coli DH5α、E.coli BL21(DE3)由軍事醫(yī)學(xué)科學(xué)院軍事獸醫(yī)研究所五室贈(zèng)送;Trans-T1感受態(tài)、原核表達(dá)載體pEASY-E1expression kit購(gòu)自北京全式金生物技術(shù)有限公司.
1.2 方法
1.2.1 培養(yǎng)基
液體LB培養(yǎng)基:蛋白胨1g,酵母粉0.5g,NaCl 1g,蒸餾水100mL,pH=7.2.
固體培養(yǎng)基(Ampicillin,Ampr):蛋白胨1g,酵母粉0.5g,NaCl 1g,瓊脂1.5g,蒸餾水100mL,pH=7.2.121℃滅菌20min.滅菌后加除菌的氨芐西林至100μg/mL.
1.2.2 引物的設(shè)計(jì)及合成
根據(jù)GenBank中B.abortus S19的序列信息設(shè)計(jì)引物.上游引物(Nde I酶切位點(diǎn))F:catatgATGATCCAGCCATCGATTACCCTGT;下游引物R(Xho I 酶切位點(diǎn)):ctcgagTCATGCTATAGCTCCCATTCCCGAG.引物由長(zhǎng)春華大中天生物技術(shù)有限公司合成.
1.2.3 wzt基因的擴(kuò)增
以B.abortus S19總DNA為模板,反應(yīng)體系50μL:模板DNA 2μL,上游引物1μL,下游引物1μL,Ex Taq酶0.5μL,10×Buffer 5μL,dNTP 4μL,水36.5μL.反應(yīng)條件:94℃5min;94℃1min,58℃30s,72℃1min,30個(gè)循環(huán);72℃延伸10min.
1.2.4 重組表達(dá)質(zhì)粒的構(gòu)建及鑒定
利用凝膠回收試劑盒純化回收PCR產(chǎn)物,將回收產(chǎn)物與pEASY-E1載體連接并轉(zhuǎn)化Trans-T1感受態(tài)細(xì)胞,轉(zhuǎn)化后加入250μL經(jīng)室溫平衡的LB培養(yǎng)基,于37℃,180r/min恒溫?fù)u床中培養(yǎng)1h.取200μL菌液均勻地涂布于準(zhǔn)備好的Ampr固體培養(yǎng)基平板上,37℃培養(yǎng)過(guò)夜.
挑取單菌落接種于LB(Amp)液體培養(yǎng)基中,180r/min,37℃恒溫?fù)u床中培養(yǎng)12h,采用載體正向引物(E Control Forward Primer:GGAATTGTGAGCGGATAACAATTCC)和下游引物R進(jìn)行PCR鑒定.
提取陽(yáng)性菌液質(zhì)粒,進(jìn)行Nde I和Xho I雙酶切鑒定,37℃水浴鍋酶切3h.酶切體系:質(zhì)粒4μL,限制性內(nèi)切酶Nde I 0.5μL,限制性內(nèi)切酶Xho I 0.5μL,Buffer 1μL,無(wú)菌水4μL.將鑒定正確的質(zhì)粒pEASY-wzt進(jìn)行測(cè)序.
1.2.5 誘導(dǎo)表達(dá)
將pEASY-wzt轉(zhuǎn)化到E.coli BL21(DE3).挑取陽(yáng)性單克隆菌落,接種到5mL含有Amp的抗性LB液體培養(yǎng)基中,37℃,180r/min培養(yǎng)6h;至菌液D(600)為0.6時(shí),加入終濃度為1mmol/L 的IPTG,37℃繼續(xù)誘導(dǎo)4h.
1.2.6 聚丙烯酰胺凝膠電泳(SDS-PAGE電泳)分析
收集1.5mL誘導(dǎo)的菌液,以未誘導(dǎo)菌液作為陰性對(duì)照,于室溫12000r/min離心1min,棄上清,用1mL PBS懸浮;12000r/min離心1min,用50μL蒸餾水懸浮沉淀,加入50μL 2×SDS Buffer,在沸水中孵育5~10min.12000r/min離心5min,取上清液進(jìn)行8%SDS-PAGE電泳分析.凝膠用考馬斯亮藍(lán)R-250染色.
1.2.7 表達(dá)蛋白質(zhì)的純化
收集100mL誘導(dǎo)表達(dá)的菌液,4℃,5000r/min離心5min;棄上清,加入100mL的PBS洗滌菌液,4℃,5000r/min離心5min;重復(fù)洗滌一次,棄上清,將菌體置于冰水中,用20mL的Binding Buffer結(jié)合液進(jìn)行重懸.超聲裂解細(xì)菌,超聲10s,間歇10s,功率400W持續(xù)工作30min,直至上清液變清澈.4℃,13000r/min離心10min,取上清,經(jīng)0.22μm的濾膜過(guò)濾后,用1mL HisTrapTMFF親和層析柱純化蛋白.
1.2.8 表達(dá)產(chǎn)物的Western Blotting鑒定
純化產(chǎn)物經(jīng)SDS-PAGE分析后,根據(jù)SDS-PAGE凝膠的大小,剪裁PVDF膜和濾紙.將濾紙放在轉(zhuǎn)膜緩沖液中完全浸透;PVDF膜先在甲醇中敏化15s,隨后放入蒸餾水中浸泡2min.組裝轉(zhuǎn)膜裝置,從陽(yáng)極到陰極依次為:8層濾紙-PVDF膜-凝膠-8層濾紙.PVDF膜要略大于凝膠,以保證所有蛋白能夠轉(zhuǎn)到膜上;并將濾紙、凝膠、PVDF膜對(duì)齊,趕走氣泡.恒流110mA,電壓控制在25V內(nèi),電轉(zhuǎn)25min.用洗滌液清洗PVDF膜3次,每次5min.加入5%脫脂奶粉常溫封閉,在搖床上輕輕搖動(dòng)1~2h.倒掉封閉液,用洗滌液洗膜3次,每次5min.一抗孵育:加入1∶200(體積比)倍稀釋的鼠抗His標(biāo)簽單克隆抗體,37℃孵育2h,倒掉一抗,洗滌液洗膜(方法同上);二抗孵育:加入1∶2000(體積比)倍稀釋的辣根過(guò)氧化物酶標(biāo)記的羊抗鼠IgG,37℃孵育2h.倒掉二抗,洗滌液洗膜(方法同上)[13].加入TMB顯色液顯色10min,加純水終止,掃描保存圖片.
1.3 wzt基因表達(dá)產(chǎn)物的生物信息學(xué)分析與結(jié)構(gòu)預(yù)測(cè)
1.3.1 wzt蛋白結(jié)構(gòu)預(yù)測(cè)
采用Discovery Studio 2.5軟件以Swiss-Model模式對(duì)蛋白質(zhì)結(jié)構(gòu)進(jìn)行預(yù)測(cè)[14].
1.3.2 wzt基因表達(dá)產(chǎn)物的生物信息學(xué)分析
在NCBI數(shù)據(jù)庫(kù)中對(duì)與wzt基因功能相關(guān)的蛋白序列進(jìn)行檢索,并采用Clustalx1.83和MEGA5.0進(jìn)行生物信息學(xué)分析.
2 結(jié)果與分析
2.1 wzt基因擴(kuò)增產(chǎn)物的鑒定
以B.abortus S19總DNA為模板,PCR擴(kuò)增wzt基因,成功獲得目的基因,擴(kuò)增產(chǎn)物經(jīng)1%瓊脂糖凝膠電泳檢查,得到與目的基因大小相符的約756bp的條帶(見(jiàn)圖1).
2.2 重組表達(dá)質(zhì)粒的酶切鑒定
將重組質(zhì)粒pEASY-wzt經(jīng)Xho I單酶切、Nde I和Xho I雙酶切進(jìn)行鑒定,得到的結(jié)果同理論值相符,表明重組表達(dá)質(zhì)粒pEASY-wzt構(gòu)建正確.經(jīng)測(cè)序,與GenBank注冊(cè)序列一致.

圖1 目的基因wzt的PCR擴(kuò)增產(chǎn)物凝膠電泳圖
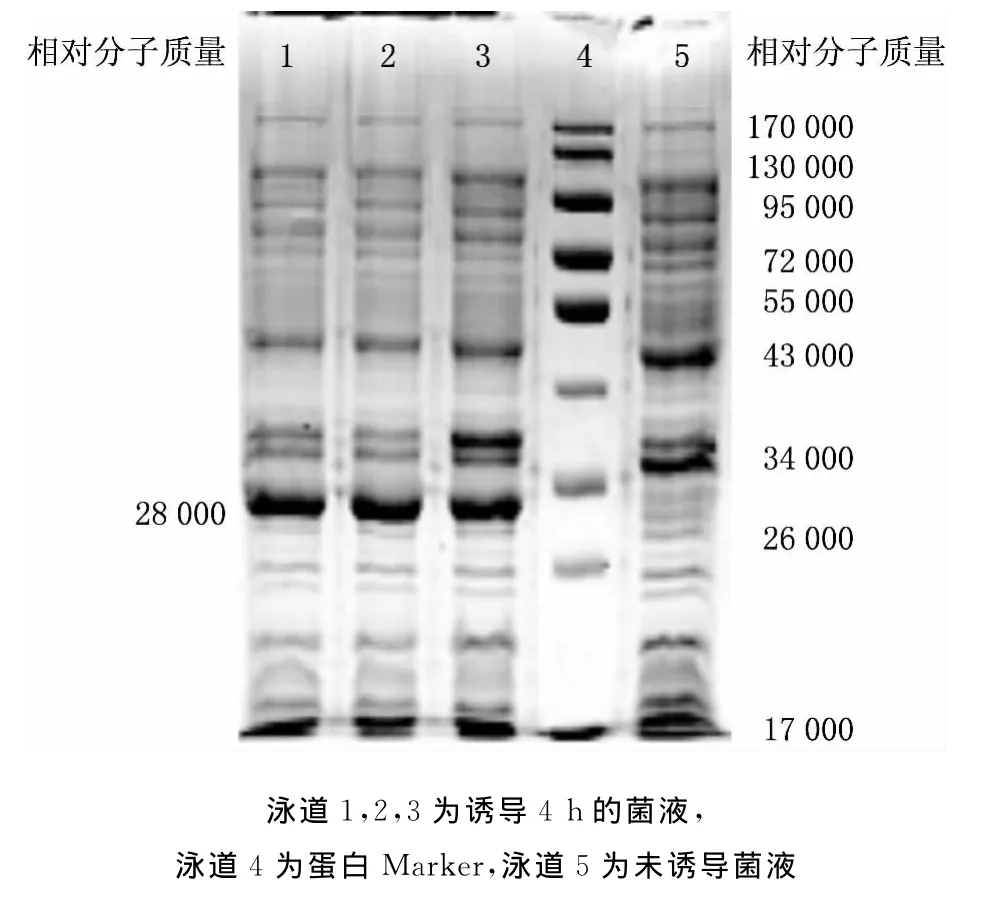
圖2 表達(dá)產(chǎn)物的SDS-PAGE分析
2.3 表達(dá)產(chǎn)物的鑒定
將構(gòu)建好的重組質(zhì)粒pEASY-wzt轉(zhuǎn)到大腸桿菌BL21中,加入IPTG使其終濃度為1.0mmol/L,37℃,誘導(dǎo)4h.通過(guò)8%SDS-PAGE分析鑒定,表明重組質(zhì)粒能夠成功表達(dá)目的蛋白,在相對(duì)分子質(zhì)量約28000處可見(jiàn)差異蛋白條帶,與理論值相符(見(jiàn)圖2),表明wzt基因成功表達(dá).
2.4 表達(dá)產(chǎn)物的分離純化
SDS-PAGE結(jié)果表明,蛋白表達(dá)成功,經(jīng)親和層析純化可獲得純化的重組蛋白,經(jīng)Lowry法測(cè)定純化蛋白的質(zhì)量濃度為0.43mg/mL.
2.5 目的蛋白的鑒定
Western Blotting分析顯示,在相對(duì)分子質(zhì)量約28000處可見(jiàn)特異反應(yīng)條帶(見(jiàn)圖3),表明純化的重組蛋白能被His標(biāo)簽單克隆抗體所識(shí)別,表明所表達(dá)蛋白為wzt蛋白.
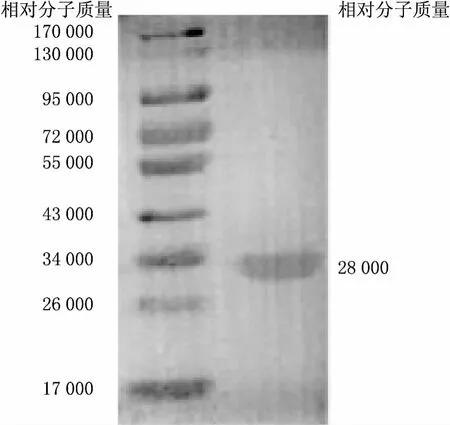
圖3 純化產(chǎn)物的Western blot分析
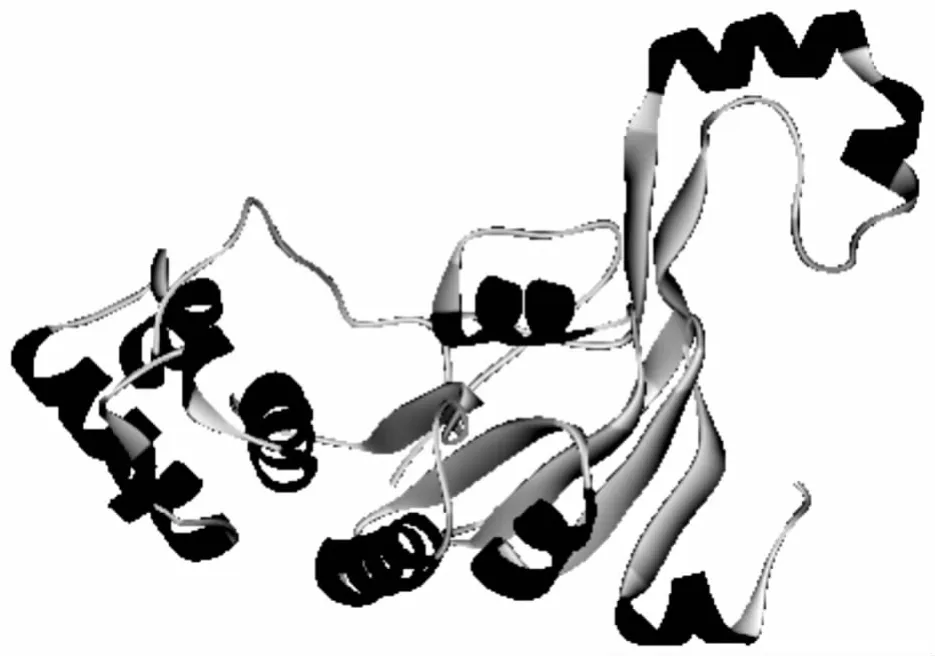
圖4 ABC轉(zhuǎn)運(yùn)系統(tǒng)結(jié)構(gòu)圖
2.6 wzt編碼蛋白的結(jié)構(gòu)預(yù)測(cè)
由Discovery Studio 2.5生成、通過(guò)Swiss-Model構(gòu)建的結(jié)構(gòu)示意圖見(jiàn)圖4.模板選擇1oxv,與來(lái)自Sulfolobus solfataricus的蔗糖ABC轉(zhuǎn)運(yùn)系統(tǒng)的ABC-ATP酶的序列一致性僅23.11%.三維構(gòu)象顯示,該蛋白可能由8個(gè)α螺旋、7個(gè)β片層構(gòu)成.
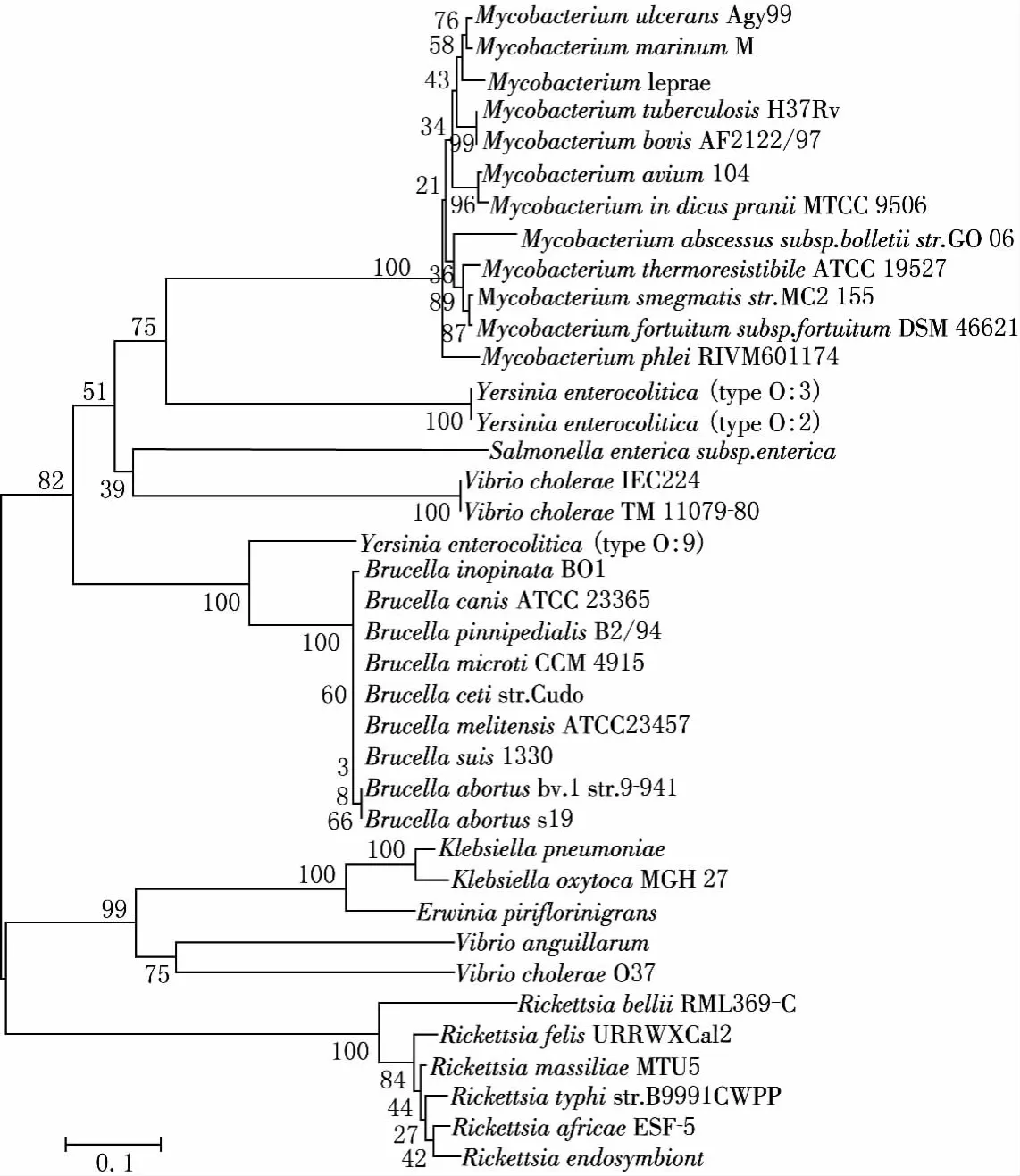
圖5 wzt基因及部分功能相同的基因編碼的蛋白序列的系統(tǒng)進(jìn)化分析
2.7 wzt編碼蛋白的同源性分析
將wzt基因編碼的蛋白序列與GenBank中功能相關(guān)的基因序列進(jìn)行同源性比較,發(fā)現(xiàn)該基因編碼的蛋白質(zhì)序列在多數(shù)種屬中具有較高的種屬特異性.布魯菌屬不同種的該基因蛋白同源性接近100%(見(jiàn)圖5),與其他種屬相關(guān)蛋白的差異較大,但與耶爾森氏菌的O:9菌株的該蛋白序列同源性較高.
3 討論
布魯菌是一種重要的人畜共患病病原,其LPS既是其主要的毒力因子,同時(shí)也是引起免疫的重要抗原,wzt基因作為其合成的關(guān)鍵基因,在O多糖跨膜合成過(guò)程中起著重要的作用.
O多糖是在內(nèi)膜上由糖基轉(zhuǎn)移酶催化形成的脂質(zhì)鏈接的O側(cè)鏈重復(fù)單位,可通過(guò)某種機(jī)制將脂質(zhì)鏈接的O多糖傳遞到細(xì)胞周質(zhì)空間,并將O多糖鏈接到核心寡糖上形成LPS[5].在細(xì)菌中參與O多糖合成的基因往往形成RFB基因[15-16],RFB基因編碼合成新型糖苷前體的糖基轉(zhuǎn)移酶,作為運(yùn)輸過(guò)程所需要的酶,因此,RFB基因座位的多態(tài)性影響O多糖的結(jié)構(gòu)多樣性[17].
目前已知有3種途徑參與O多糖的合成運(yùn)輸過(guò)程,分別為WZY依賴型途徑、ABC轉(zhuǎn)運(yùn)型和合酶依賴型途徑.在WZY依賴型途徑中,由wzx蛋白同源類似物完成跨膜運(yùn)輸;而在ABC轉(zhuǎn)運(yùn)型途徑中則由ABC轉(zhuǎn)運(yùn)子完成運(yùn)輸,其ATP結(jié)合域的序列高度保守,特別是核苷酸結(jié)合區(qū)[18].本研究的結(jié)果顯示,ABC轉(zhuǎn)運(yùn)系統(tǒng)中的wzt蛋白、ATP結(jié)合域,在布魯菌中具有高度保守的種屬特異性,因此ATP結(jié)合域的保守性具有屬的特異性.目前,有關(guān)O多糖合成的ABC轉(zhuǎn)運(yùn)子的確切組織形式和模式尚未見(jiàn)報(bào)道,沒(méi)有結(jié)構(gòu)數(shù)據(jù)可以參考,大部分信息是來(lái)自大腸桿菌等細(xì)菌的2型莢膜合成過(guò)程的研究和假說(shuō)[18-19].本研究發(fā)現(xiàn)wzt蛋白具有8個(gè)α螺旋、7個(gè)β片層結(jié)構(gòu),為進(jìn)一步研究ABC轉(zhuǎn)運(yùn)系統(tǒng)的組織形式及形成模式提供了依據(jù).
本研究成功表達(dá)了wzt蛋白,并對(duì)其結(jié)構(gòu)進(jìn)行了初步的預(yù)測(cè),研究結(jié)果為進(jìn)一步揭示wzt基因在LPS合成過(guò)程中的作用機(jī)制,闡述其在細(xì)菌組成物質(zhì)的合成和免疫調(diào)節(jié)中的作用具有重要的意義.
[1]ALTON G G,JONES L M,ANGUS R D,et al.Techniques for the Brucellosis Laboratory[R].Institut National de la Recherche Agronomique,1988.
[2]CARDOSO P G,MACEDO G C,AZEVEDO V,et al.Brucellaspp noncanonical LPS:structure,biosynthesis,and interaction with host immune system[J].Microb Cell Fact,2006,23:5-13.
[3]GODFROID F,CLOECKAERT A,TAMINIAU B,et al.Genetic organisation of the lipopolysaccharide O-antigen biosynthesis region of Brucella melitensis16M (wbk)[J].Res Microbiol,2000,151:655-668.
[4]CLOECKAERT A,GRAYON M,VERGER J M,et al.Conservation of seven genes involved in the biosynthesis of the lipopolysaccharide O-side chain in Brucellaspp[J].Res Microbiol,2000,151:209-216.
[5]RAETZ C R H ,WHITFIELD C.Lipopolysaccharide endotoxin[J].Annu Rev Biochem,2002,71:635-700.
[6]JIANG X,LEONARD B,BENZON R,et al.Macrophage control of Brucella abortus:role of reactive oxygen intermediates and nitric oxide[J].Cell Immunol,1993,151:309-319.
[7]WANG Z,NIU J R,WANG S S,et al.In vivo differences in the virulence,pathogenicity and induced protective immunity of wboA mutants from genetically different parent Brucellaspp.[J].Clin Vaccine Immunol,2013,20(2):174-180.
[8]ADONE R,F(xiàn)RANCIA M,PLSTOIA C,et al.Protective role of antibodies induced by Brucella melitensis B115against B.melitensis and B.abortus infections in mice[J].Vaccine,2012,30(27):3992-3995.
[9]TOMII K ,KANEHISA M.A comparative analysis of ABC transporters in complete microbial genomes[J].Genome Res,1998,8(10):1048-1059.
[10]GODFROID F,TAMINIAU B,DANESE I,et al.Identification of the perosamine synthetase gene of Brucella melitensis 16M and involvement of lipopolysaccharide O side chain in Brucella survival in mice and in macrophages[J].Infect Immun,1998,66:5485-5493.
[11]WANG X,WANG L,LU T,et al.Effects of partial deletion of the wzm and wzt genes on lipopolysaccharide synthesis and virulence of Brucella abortus S19[J].Mol Med Rep,2014,9(6):2521-2527.
[12]WANG X R,YAN G M,ZHANG R,et al.Immunogenic response induced by wzm and wzt gene deletion mutants from Brucella abortus S19[J].Mol Med Rep,2014,9(2):653-658.
[13]張瑞,王秀然,夏力亮,等.牛布氏菌virB8基因的克隆及原核表達(dá)[J].中國(guó)生物制品學(xué)雜志,2012,25(7):841-843.
[14]劉艷.二甲氧基硫代磷酸酯類農(nóng)藥單克隆抗體研制及單鏈抗體三維結(jié)構(gòu)模擬[D].南京:南京農(nóng)業(yè)大學(xué),2009.
[15]POPOFF M Y,LE MINOR L.Expression of antigenic factor O:54is associated with the presence of a plasmid in Salmonella[J].Ann Inst Pasteur Microbiol,1985,136B:169-177.
[16]KEENLEYSIDE W J,PERRY M B,MACLEAN L L,et al.A plasmid-encoded rfbO:54gene cluster is required for biosynthesis of the O:54antigen in Salmonella enterica serovar Borreze[J].Mol Microbiol,1994,11:437-448.
[17]AL-HENDY A,TOIVANEN P,SKURNIK M.The effect of growth temperature on the biosynthesis of Yersinia enterocolitica O:3lipopolysaccharide:temperature regulates the transcription of the rfb but not of the rfa region[J].Microb Pathog,1991,10:81-86.
[18]MANTING E H,DRIESSEN A J.Escherichia coli translocase:the unravelling of a molecular machine[J].Mol Microbiol,2000,37:226-238.
[19]CHU S,NOONAN B,CAVAIGNAC S,et al.Endogenous mutagenesis by an insertion sequence element identifies Aeromonas salmonicida AbcA as an ATP-binding cassette transport protein required for biogenesis of smooth lipopolysaccharide[J].Proc Natl Acad Sci USA,1995,92:5754-5758.
