HIV-1膜蛋白gp120 V3環的神經毒性研究
2014-10-25 09:37:02張志麗陳兵剛姜太一喬錄新陳德喜張玉林
首都醫科大學學報 2014年1期
關鍵詞:檢測
張志麗 陳兵剛 姜太一 喬錄新 吳 昊 陳德喜 張玉林*
(1.首都醫科大學附屬北京佑安醫院感染中心,北京100069;2.山東省濱州市中心醫院感染內科,山東濱州251700;3.北京市肝病研究所,北京100069)
在高活性抗反轉錄病毒治療(highly active antiretroviral therapy,HAART)時代,人類免疫缺陷病毒-1(human immunodeficiency virus-1,HIV-1)相關慢性合并癥——HIV-1相關神經損傷(HIV-1 associated neurological impairment,HANI)逐漸顯現,成為影響患者生活質量和預后的重要危險因素[1]。HANI發病機制目前尚不完全清楚。一般認為可能與HIV病毒感染所引起的中樞神經系統(central nervous system,CNS)炎性反應有關。其核心是HIV首先通過CCR5(C-C chemokine receptor 5)和CXCR4(C-X-C chemokine receptor 4)受體于外周感染單核/巨噬細胞,由其攜帶病毒損傷并透過血-腦脊液屏障(blood brain barrier,BBB)進入CNS,定植或感染其中的小膠質細胞和星形膠質細胞,從而導致局部炎性反應,表現為炎性反應細胞激活、細胞因子和趨化因子分泌增加以及病毒顆粒(包括病毒膜蛋白gp120)的釋放和氧化應激反應等,最終使得神經元功能障礙及凋亡,累積發生神經認知損傷(neurocognitive impairment,NCI)[2-4]。本研究擬通過原代神經細胞培養,體外觀察HIV-1包膜蛋白gp120神經毒性。
1 材料與方法
1.1 實驗動物與材料
Balb/C小鼠購自中國軍事醫學科學院〔實驗動物許可證號:SYXK(京)2009-0020〕;重組gp120 V3環多肽購自北京Sino生物公司;D-Glucose、β-actin單克隆抗體購自Sigma公司(美國);神經細胞培養基Neurobasal、B27、青鏈霉素、1 × Hanks液、青/鏈霉素、Glutamax購自Gibco公司(美國);CytoTox96非放射性細胞毒性檢測試劑盒購自Promega公司(美國);抗微管蛋白2(MAP-2)抗體、抗bcl-2單克隆抗體購自Santa Cruz公司(美國);兔抗bax多克隆抗體購自Abcam公司(英國);Cy3標記兔抗小鼠IgG購自BioLegend公司(加拿大);牛血清白蛋白(BSA)、原位細胞凋亡檢測試劑盒(TUNEL)購自羅氏公司(美國);辣根過氧化物酶標記抗小鼠和抗兔二抗購自Jackson公司(美國);其他相關分析純試劑購自國內生產公司。
1.2 原代神經細胞培養和gp120 V3環神經毒性試驗
動物實驗嚴格按照首都醫科大學動物保護及實驗動物操作規范執行。取孕16 d后balb/C胎鼠腦組織,在顯微鏡下于低溫解剖分離液(1×Hanks平衡鹽溶液+2.5 mmol/L Hepes+30 mmol/L D-Glucose+2 mmol/L CaCl2+0.5 mmol/L MgSO4+2 mmol/L NaHCO3+0.03%BSA+1%青/鏈霉素)中剝除硬腦膜,分離出大腦皮質。解剖分離液清洗2遍后輕柔吹打,200目篩網過濾后,1 500 r/min離心3 min,棄上清液,加神經細胞培養液(Neurobasal+1×Glutamax+2%B27+1%青/鏈霉素),接種入右旋多聚賴氨酸包被的細胞培養板。對于預加蓋玻片的培養板,預先依次將蓋玻片在100%丙酮中洗滌40 min,無水乙醇洗滌20 min,0.1 mol/L鹽酸洗滌40 min,蒸餾水洗滌10 min,高壓滅菌25 min;然后將蓋玻片放入培養板孔中經由右旋多聚賴氨酸包被后使用。每孔分別加培養液0.5 mL(24孔板)、1 mL(12孔板)和2 mL(6孔板)。細胞接種濃度分別為每孔2.5×105(24孔板)、5.0×105(12孔板)和12×105(6孔板),置于37℃、體積分數為0.05 CO2恒溫培養箱溫育,每3 d更換一半培養液。依據既往文獻[5]說明,于神經細胞培養第7天,分別向培養基中加入 0、200、500、1 000 pmol/L重組gp120蛋白,37℃下孵育24 h,進行神經毒性試驗。以上實驗都在超凈臺內無菌條件下進行,相同條件重復3次,且繼續以下實驗。
1.3 神經細胞毒性及凋亡檢測
通過FITC熒光標記的TUNEL檢測試劑盒評估細胞凋亡:4%多聚甲醛固定細胞,用磷酸鹽緩沖液沖洗后按TUNEL檢測試劑盒說明書操作,DAPI染色觀察細胞核形態。同時,通過測定培養基中的乳酸脫氫酶(LDH)來定量評估gp120的細胞毒性,培養基中LDH值越高則細胞凋亡/死亡比例就越高。按照商用乳酸脫氫酶試劑盒CytoTox96廠家說明書操作,簡言之,收集神經細胞培養基與反應底物各50 μL加入96孔板中室溫孵育30 min,再加終止液使反應停止,通過Emax精密酶標儀(Molecular Devices)測定490 nm波長下樣本的吸光度。設定純神經細胞培養基為陰性對照(0%死亡),加入裂解液后細胞完全裂解的培養基為陽性對照(100%死亡),每樣本測量3次取平均值。檢測樣本死亡細胞比例(%)=[(檢測樣本吸光度-陰性對照吸光度)/(陽性對照吸光度-陰性對照吸光度)]×100%。
1.4 神經突起測量
通過抗MAP-2單克隆抗體免疫熒光顯像觀察神經突起形態學變化,具體實驗步驟如下:4%多聚甲醛室溫固定15 min;1×PBS洗滌15 min;1%Triton穿孔15 min;1×PBS洗滌15 min×2次;1%BSA+2%羊血清+1×PBS 37℃封閉1 h;1∶1 000濃度MAP-2單克隆抗體(封閉液稀釋)4℃過夜;1×PBS洗滌15 min×3次;1∶1 000濃度Cy3標記抗小鼠二抗(封閉液稀釋)37℃溫育1 h;1×PBS洗滌20 min×3次;DAPI封片后于德國Leicadm500b型正向熒光顯微鏡下閱片并攝片(每個點至少選取3個視野,取平均數);使用Image-Pro Plus 6.0圖像分析軟件分析神經元突起數量和長度。
1.5 統計學方法
細胞計數、神經元突起長度分析使用Image-Pro Plus 6.0圖像分析軟件。使用PASW statistics 18統計分析軟件進行統計分析。計量資料用均數±標準差(±s)表示,且給出雙側95%可信區間,組間比較采用單因素方差分析和多重均數比較。以P<0.05為差異有統計學意義。
2 結果
2.1 gp120可誘導神經細胞凋亡
HIV包膜蛋白gp120能夠通過V3環與神經元表面表達的趨化因子受體CCR5、CXCR4相互作用,直接或間接誘導神經細胞凋亡。本實驗中,神經細胞培養第8 天,向培養基中加入 0、200、500、1 000 pmol/L 重組gp120 V3環多肽37℃孵育24 h。TUNEL檢測顯示隨著gp120 V3環重組蛋白濃度增加,凋亡細胞數量明顯增加(圖1A)。通過培養基LDH檢測得出的不同濃度V3環細胞毒性所致細胞死亡比例分別為:(21.16±3.56)%(對照組)、(35.23 ±10.42)%(200 pmol/L組)、(57.45±9.26)%(500 pmol/L 組)、(62.21±14.35)%(1 000 pmol/L組),除了500 pmol/L組與1 000 pmol/L組之間差異無統計學意義外(P>0.05),其余各組間差異均有統計學意義(P<0.01)(圖1B)。
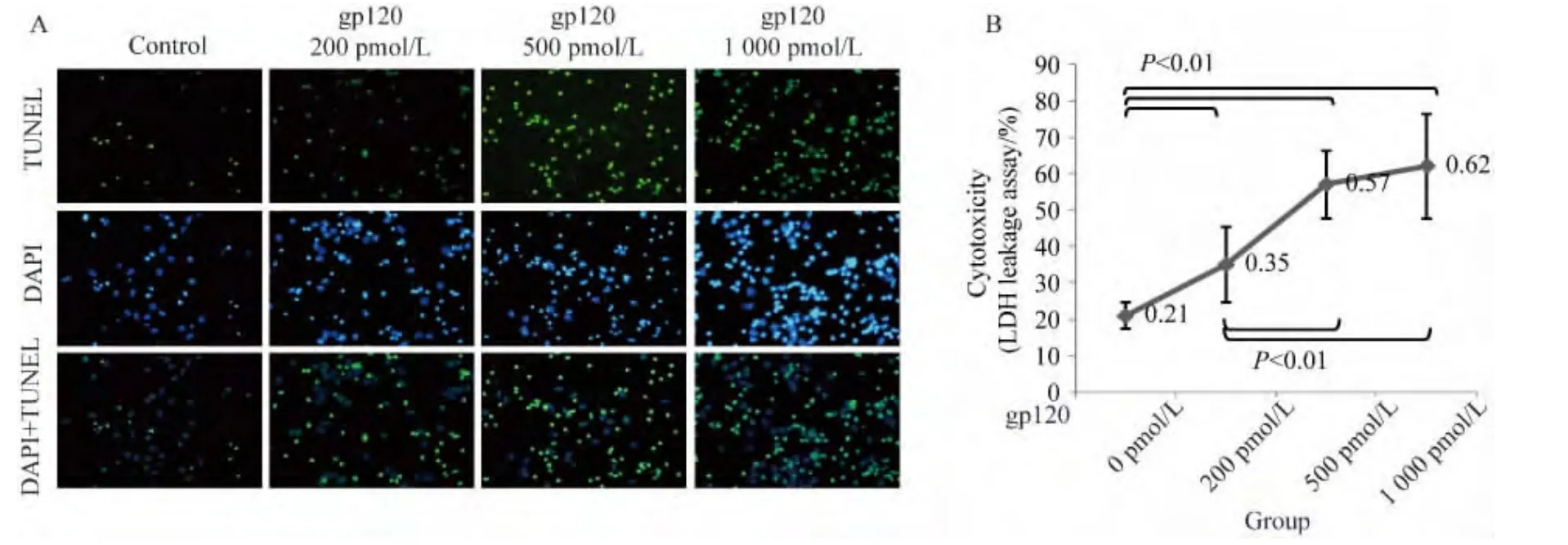
圖1 gp120神經細胞毒性Fig.1 HIV-1 envelope protein gp120 induced neuronal apoptosis
2.2 gp120可抑制神經突起形成
本實驗中通過免疫熒光Cy3標記的抗神經纖維特異性MAP-2抗體檢測gp120 V3環對神經突起造成的損傷(圖2A)。對照組、200 pmol/L組、500 pmol/L組和1 000 pmol/L組平均每個神經元突起數量分別為6.26±2.12、4.85±1.68、4.23±1.08和 3.81±0.86,除200 pmol/L組與500 pmol/L組,500 pmol/L組與1 000 pmol/L組外(P>0.05),其余各組間差異有統計學意義(P<0.05)(圖2B)。各組平均神經突起長度為(900.24±169.06)μm(對照組)、(679.32±157.37)μm(200 pmol/L 組)、(445.64 ±56.13)μm(500 pmol/L組)和(173.39±29.58)μm(1 000 pmol/L組)。神經突起長度隨gp120濃度增加而縮短(P<0.01)(圖2C)。因此,HIV包膜蛋白gp120不僅誘導神經細胞凋亡,還會抑制神經突起形成。
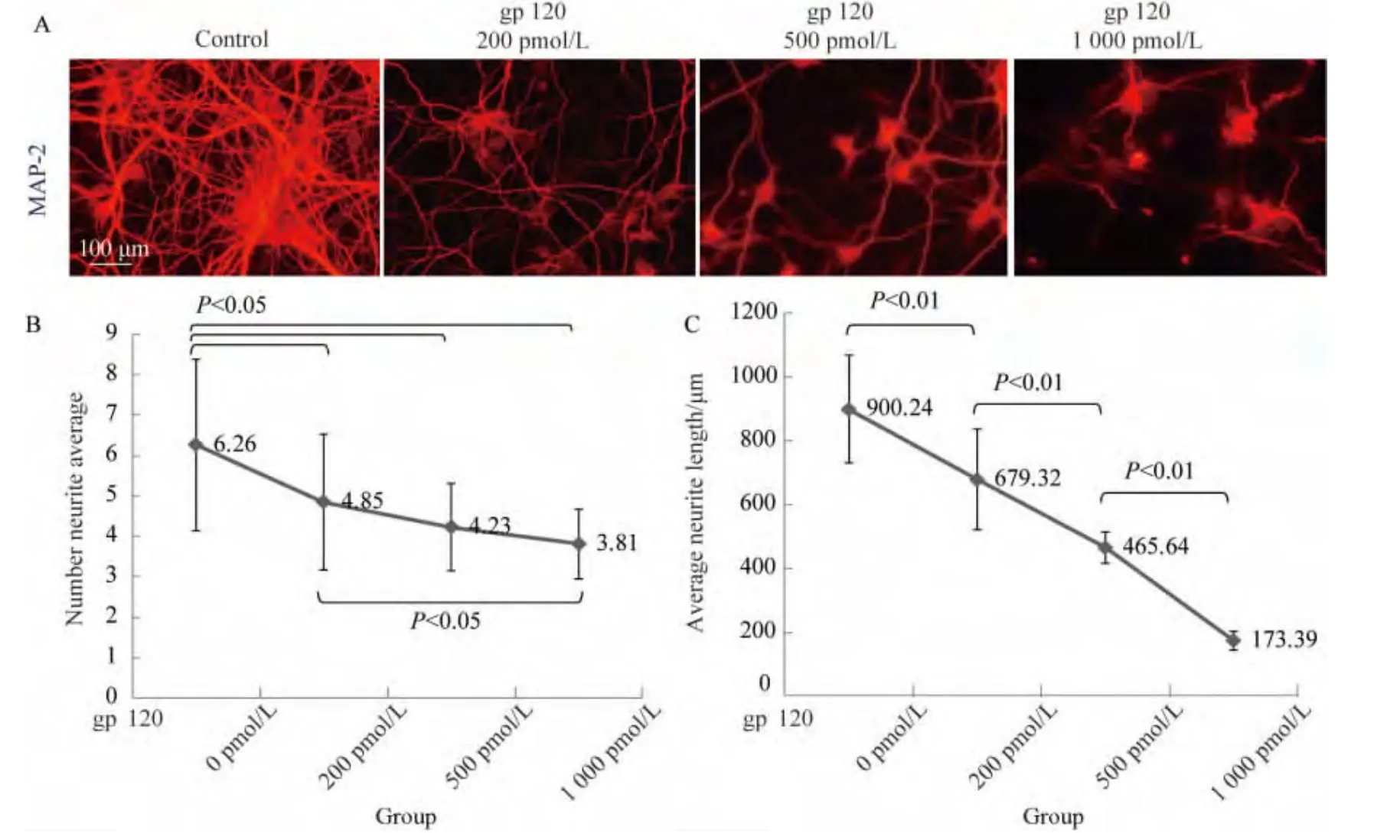
圖2 gp120抑制神經突起形成Fig.2 HIV-1 envelope protein gp120 inhibited neurite outgrowth
2.3 gp120 V3環的神經毒性與bcl-2低表達有關
Gp120誘導bcl-2蛋白低表達,但對bax蛋白低表達無影響(圖3)。可見gp120 V3環通過抑制bcl-2表達來誘導神經細胞凋亡。
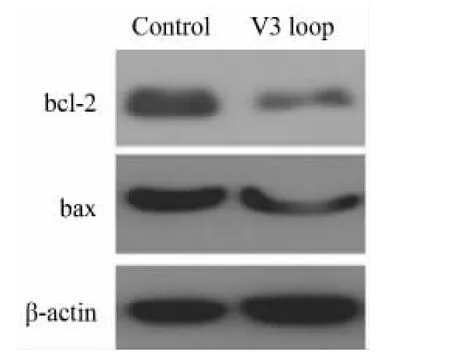
圖3 蛋白印跡檢測不同條件下凋亡調控蛋白bcl-2與bax蛋白表達水平的變化Fig.3 The change of bcl-2 and bax protein expression under different experimental conditions via Western blotting
3 討論
HIV相關神經認知障礙與感染HIV的單核細胞透過血-腦脊液屏障,在CNS產生炎性反應有關[6]。除了受HIV感染的炎性反應(單核/巨噬細胞系統和膠質細胞)細胞釋放的各種炎性介質外,感染的炎性反應細胞還可以釋放HIV-1相關蛋白,繼而產生病毒蛋白相關的興奮性神經毒性,導致癥狀輕重不等的神經認知障礙[7-8]。HIV病毒調控元件包括長末端重復序列(long terminal repeat,LTR)、基因編碼的病毒蛋白Tat、Vpr、Nef和膜基因編碼的包膜蛋白 gp120與gp41 等[9]。
本研究發現gp120除了能夠誘導神經細胞凋亡外,還能影響神經突起的發育和生長,表明gp120在誘導神經元細胞凋亡之前可能已經抑制神經突觸傳遞,從而影響神經元功能[10-11]。與神經細胞凋亡相比,gp120或其他病毒蛋白對神經軸突與樹突的損傷性抑制,對于無癥狀性和輕度神經認知障礙的發生可能更為重要[12-13]。除了誘導神經細胞凋亡外,有研究[14-15]報道gp120還可通過蛋白激酶C途徑以及受體介導的Ca2+釋放,改變人腦微血管內皮細胞中緊密連接蛋白的表達,導致細胞骨架的改變、單核細胞內移增加。多項研究[16-18]證明gp120擾亂神經細胞鈣穩態、破壞線粒體膜完整性,導致細胞色素C釋放、激活caspase和核酸內切酶,而gp120對神經元鈣穩態的破壞是通過干擾質膜與內質網中的鈣調控系統來實現的[19-20]。
Bcl-2家族蛋白通過調控線粒體外膜的透化作用,使神經細胞暴露于多種不同的促死亡信號[21]。Bcl-2家族中主要抗凋亡成員Bcl-2的表達會降低線粒體外膜的透化作用,以及線粒體死亡誘導因子如Ca2+及細胞色素C向胞質的釋放[22]。而bcl-2家族中主要促凋亡成員bax的表達作用相反。因此,線粒體膜的透化作用是gp120誘導神經細胞凋亡的通路。本研究發現,重組人gp120 V3環多肽可誘導神經元凋亡和抑制神經突起形成,并呈濃度依賴性。同時,gp120 V3環誘導神經細胞凋亡與抑制bcl-2表達有關。
[1]Heaton R K,Clifford D B,Franklin D R Jr,et al.HIV-associated neurocognitive disorders persist in the era of potent antiretroviral therapy:CHARTER Study[J].Neurology,2010,75(23):2087-2096.
[2]Wu D T,Woodman S E,Weiss J M,et al.Mechanisms of leukocyte trafficking into the CNS[J].J Neurovirol,2000,6 Suppl 1:S82-85.
[3]Gras G,Kaul M.Molecular mechanisms of neuroinvasion by monocytes-macrophages in HIV-1 infection[J].Retrovirology,2010,7:30.
[4]Wang H,Sun J,Goldstein H.Human immunodeficiency virus type 1 infection increases the in vivo capacity of peripheral monocytes to cross the blood-brain barrier into the brain and the in vivo sensitivity of the blood-brain barrier to disruption by lipopolysaccharide[J].J Virol,2008,82(15):7591-7600.
[5]Meucci O,Miller R J.gp120-induced neurotoxicity in hippocampal pyramidal neuron cultures:protective action of TGF-beta1[J].J Neurosci,1996,16(13):4080-4088.
[6]丁渭,張玉林,喬錄新,等.氧化損傷在HIV相關神經認知損害中的作用[J].首都醫科大學學報,2012,33(5):602-609.
[7]Cicala C,Arthos J.Virion attachment and entry:HIV gp120 env biotinylation,gp120 Env,or integrin ligandbinding assay[J].Methods Mol Biol,2014,1087:3-12.
[8]Tuzer F,Madani N,Kamanna K,et al.HIV-1 Env gp120 structural determinants for peptide triazole dual receptor site antagonism[J].Proteins,2013,81(2):271-290.
[9]Roberts A J,Maung R,Sejbuk N E,et al.Alteration of methamphetamine-induced stereotypic behaviour in transgenic mice expressing HIV-1 envelope protein gp120[J].J Neurosci Methods,2010,186(2):222-225.
[10]張紅梅,孫玉,柳雅立,等.gp120轉基因小鼠制備及初步鑒定[J].首都醫科大學學報,2010,31(6):695-700.
[11]蓋祥云,趙瑛,劉琳,等.腦缺血后神經細胞凋亡致血管性癡呆的研究進展[J].中國腦血管病雜志,2010,7(11):614-616.
[12]Sarkar R,Mitra D,Chakrabarti S.HIV-1 Gp120 protein downregulates nef induced IL-6 release in immature dentritic cells through interplay of DC-SIGN[J].PLoS One,2013,8(3):e59073.
[13]焦艷梅,吳昊.HIV-1潛伏庫研究進展[J].首都醫科大學學報,2009,30(5):639-642.
[14]Kanmogne G D,Schall K,Leibhart J,et al.HIV-1 gp120 compromises blood-brain barrier integrity and enhances monocyte migration across blood-brain barrier:implication for viral neuropathogenesis[J].J Cereb Blood Flow Metab,2007,27(1):123-134.
[15]Tintori C,Selvaraj M,Badia R,et al.Computational studies identifying entry inhibitor scaffolds targeting the Phe43 cavity of HIV-1 gp120[J].Chem Med Chem,2013,8(3):475-483.
[16]Li W,Galey D,Mattson M P,et al.Molecular and cellular mechanisms of neuronal cell death in HIV dementia[J].Neurotox Res,2005,8(1-2):119-134.
[17]Mattson M P,Haughey N J,Nath A.Cell death in HIV dementia[J].Cell Death Differ,2005,12 Suppl 1:893-904.
[18]Shah A,Kumar S,Simon S D,et al.HIV gp120-and methamphetamine-mediated oxidative stress induces astrocyte apoptosis via cytochrome P450 2E1[J].Cell Death Dis,2013,4:e850.
[19]Haughey N J,Mattson M P.Calcium dysregulation and neuronal apoptosis by the HIV-1 proteins Tat and gp120[J].J Acquir Immune Defic Syndr,2002,31 Suppl 2:S55-61.
[20]Scharf L,West A P Jr,Gao H,et al.Structural basis for HIV-1 gp120 recognition by a germ-line version of a broadly neutralizing antibody[J].Proc Natl Acad Sci U S A,2013,110(15):6049-6054.
[21]Adams J M,Cory S.The Bcl-2 apoptotic switch in cancer development and therapy[J].Oncogene,2007,26(9):1324-1337.
[22]Ow Y P,Green D R,Hao Z,et al.Cytochrome c:functions beyond respiration[J].Nat Rev Mol Cell Biol,2008,9(7):532-542.
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
