牛瑟氏泰勒蟲p23-IL-18融合基因的原核表達
2014-11-23 03:55:38許應天吳艷麗于志云
中國獸藥雜志 2014年7期
白 楊,許應天,吳艷麗,于志云
(1.吉林正業生物制品股份有限公司,吉林吉林132101;2.延邊大學,吉林延吉133002)
牛瑟氏泰勒蟲病是由瑟氏泰勒蟲寄生于牛的紅細胞和單核巨噬系統細胞內引起的一種蜱傳播性血液原蟲病[1]。該病在本地牛多數呈帶蟲,染蟲率較低,外地引進牛和改良牛染蟲率較高,死亡率也很高[2]。牛瑟氏泰勒蟲p23主要表面蛋白是紅細胞感染階段的一種蟲體表面蛋白,當用孢子持續感染牛時,雖然瑟氏泰勒蟲表面抗原不斷發生變化,卻始終都能檢測到 p23表面蛋白的表達[3]。p23因具有較好的免疫原性[4],故是診斷和預防該病的理想抗原之一[5]。
白細胞介素18(interleukin,IL-18)是近年來新發現的一種重要的細胞免疫調節因子,具有多種生物學功能,其應用研究十分活躍[6-9]。IL-18屬于IL-1家族成員,是機體先天性和獲得性免疫的重要調節因子。IL-18在慢性炎癥、自體免疫性疾病、各種各樣的癌變及眾多傳染病的發生過程中都有表達,所以IL-18自發現以來就一直受到各國研究人員的關注。1999年,IL-18基因的克隆和IL-18活性分析由 Shoda[10]完成。2002 年 Nagata T 等[11]將該基因在家蠶中表達,實驗是通過桿狀病毒系統完成。目前,研究較多的也是其在參與機體免疫方面的功能,在提高牛的抗感染力、增強滅活苗或基因工程苗免疫作用方面具有廣闊的應用前景。本研究以pMD18-T-p23-IL-18質粒為材料,構建牛瑟氏泰勒蟲原核表達質粒pET-28a-p23-IL-18,并進行表達,以期為牛瑟氏泰勒蟲病基因工程亞單位疫苗的制備提供了理論依據。
1 材料與方法
1.1 質粒、菌種和載體 質粒pVAXI-p23、pMD18-IL-18、DH5α菌種、pET-28a載體菌液均由延邊大學預防獸醫實驗室保存。
1.2 主要試劑 DNA Ligation Kit Ver.2.1,pMD18-T-Simple 載體,T4 DNA Lingase,BamH I,EcoR I,ExTaq酶,質粒小量抽提試劑盒,逆轉錄酶,DNA Maker,FITC標記的羊抗牛IgG,RNase和巰基乙醇,均為Sigma公司產品。
1.3 引物的設計與合成 根據GenBank中報道的牛瑟氏泰勒蟲p23序列(D84447)及牛白細胞介素18cDNA序列(BC102879)設計了兩對特異性的引物。其中含有長度為45 bp的linker序列為中間接頭(Gly4Ser)3核苷酸,由p2和p3共同編碼,即方框的部分。p2引物與p3引物的5’端有24 bp的堿基互補,為陰影部分。在p1與p4引物上含有酶切位點,設計的引物由上海生工生物工程技術服務有限公司合成。p1和p2用于擴增IL-18基因序列,預測擴增長度為526 bp,重組后目的片段長度約為1048 bp。
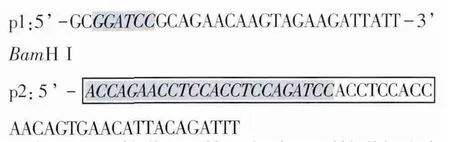
p3和p4用于擴增p23基因序列,預測擴增長度為522 bp。p3:
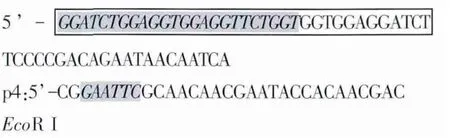
1.4 重組基因的構建 參考文獻劉娟等的試驗方法[12]以現有質粒pMD18-IL-18為模板,以p1和p2為引物,擴增IL-18基因片段;以pVAXI-p23為模板,以p3和p4為引物,擴增p23基因片段。將以上2次PCR產物混合作為PCR反應模板,以pl和p4為引物,進行PCR擴增,重組基因p23-IL-18。
1.5 克隆載體pMD18-T-p23-IL-18的構建及鑒定 將純化后的p23-IL-18與pMD18-TSimple Vector進行連接。連接體系:PCR回收產物4 μL,pMD18-T-Simple Vector 1 μL,Vector Buffer 5 μL,4℃鏈接過夜。連接產物轉化感受態DH5α大腸桿菌。挑取經Amp抗性篩選的菌落后培養,提取質粒,經PCR、酶切鑒定正確后命名為pMD18-T-p23-IL-18。
1.6 原核表達載體pET-p23-IL-18的構建及鑒定 將pMD18-T-p23-IL-18與pET-28a載體分別用BamH I和EcoR I進行雙酶切,酶切產物純化后用T4 DNA連接酶連接。連接體系為:T4 DNA Lingase 1 μL,T4 DNA ligation buffer 2.5 μL,p23-IL-18 10 μL,pET-28a 2 μL,16 ℃連接過夜。連接產物轉化感受態DH5α大腸桿菌,小量提取質粒。經 PCR(引物為 p1和 p4)、酶切(BamH I和EcoR I)、測序鑒定正確后的重組質粒命名為pET-p23-IL-18。
1.7 重組表達質粒的誘導表達 將鑒定為陽性的重組表達菌50 μL接種于5 mL LB培養液(含Amp 100 μg/mL)中,37℃過夜培養后,按1%接種于新鮮 LB培養液中,振搖至OD600nm為0.6,加入IPTG至終濃度為1 mmol/L,37℃誘導表達。分別在0、2、4、6和8 h各收集2 mL菌液,同時收集未誘導的菌液作為陰性對照。SDS-PAGE電泳后考馬斯亮藍染色觀察,并作Western-blotting分析。
2 結果與分析
2.1 目的基因的擴增 取5 μL的PCR產物使用21.0%瓊脂糖凝膠進行電泳檢測。p23與IL-18基因擴增結果顯示均在500 bp左右出現特異性條帶,與預期大小相符合(圖1);重組后基因擴增結果顯示在1000 bp左右出現特異性條帶,與預期大小相符合(圖2)。
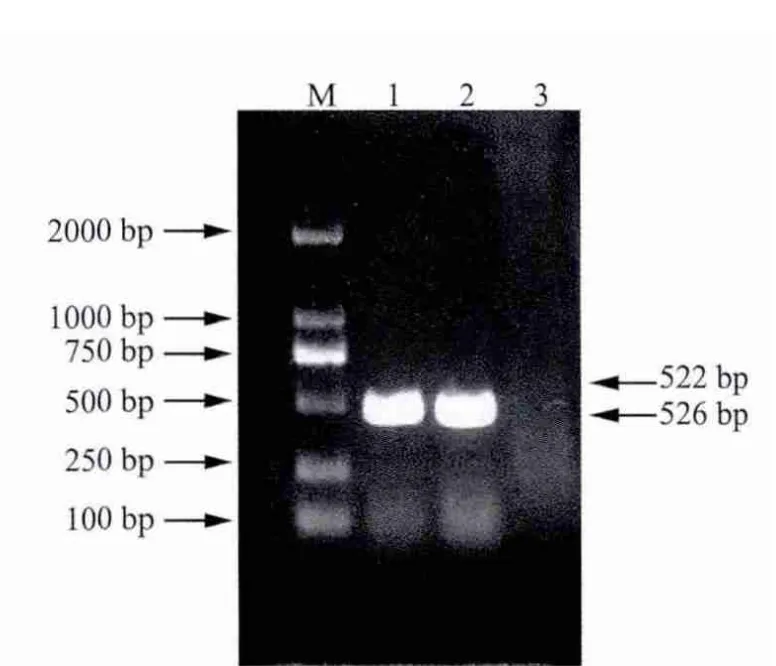
圖1 p23與IL-18基因的PCR擴增
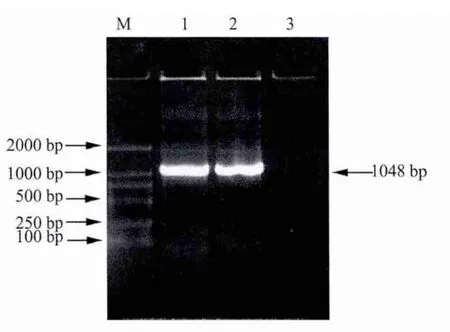
圖2 p23-IL-18融合基因的PCR擴增
2.2 陽性克隆載體的鑒定 重組克隆質粒用特異性引物p1,p4擴增出1000 bp左右出現特異性條帶,然后用 EcoRⅠ和 BamHⅠ酶切出長度為1000 bp左右的片段(圖3),說明目的片段已成功擴增并正確插入到克隆載體pMD-18-T Simple中。測序結果表明,克隆得到的重組基因p23-IL-18基因片段大小為1048 bp。此基因片段與GenBank中已發表的牛瑟氏泰勒蟲p23核苷酸序列、牛IL-18核苷酸序列同源性為99%。
2.3 重組表達質粒pET-28a-p23-IL-18的鑒定 用BamHⅠ和EcoRⅠ對初步篩選為陽性的重組質粒進行酶切鑒定,酶切產物于1%瓊脂糖凝膠中電泳。結果顯示,經BamHⅠ和EcoRⅠ酶切,得到了約3000 bp和1048 bp的兩條片段(圖4),說明目的基因片段已正確地克隆到pET-28a原核表達載體中。
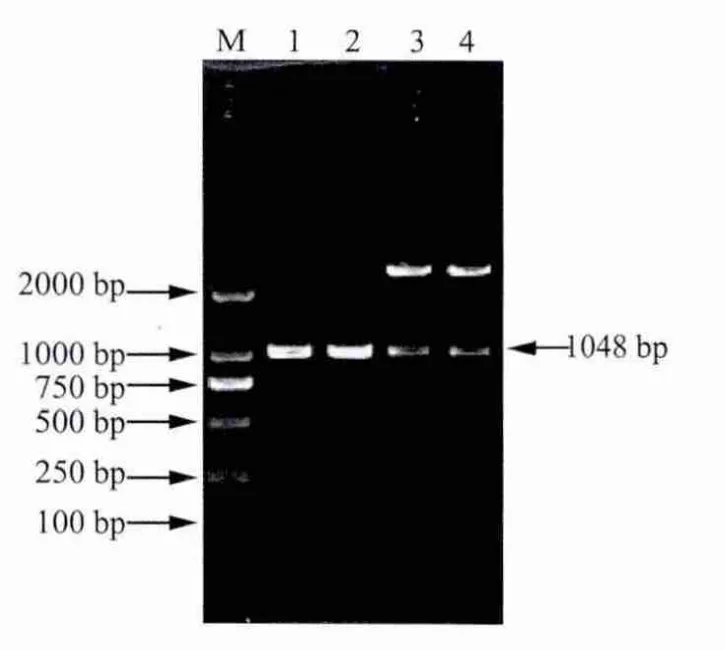
圖3 重組質粒pMD18-T-p23-IL-18的PCR及酶切鑒定結果

圖4 重組質粒pET-28a-p23-IL-18的PCR及酶切鑒定圖
2.4 目的基因的誘導表達 誘導的pET-28ap23-IL-18陽性菌在約48 kDa處出現明顯特異性條帶(圖5),與預期結果相符,重組表達菌在6 h時表達量最高。
2.5 表達產物的Western-blotting檢測 表達產物經SDS-PAGE電泳后轉移到PVDF膜上進行Western-blotting檢測,在PVDF膜上出現特異性條帶,表達產物能被牛瑟氏泰勒蟲陽性血清所識別,證明目的基因的表達產物具有較好的反應原性(圖6)。
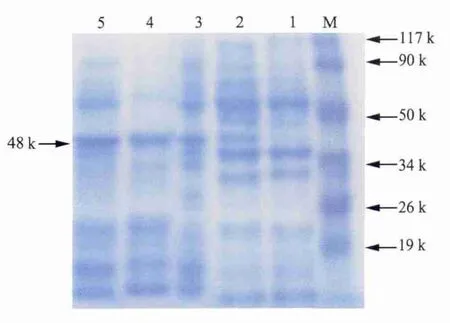
圖5 表達產物的SDS-PAGE電泳

圖6 表達產物的Western-blotting分析
3 討論
細胞因子通過復雜網絡調節機體免疫,細胞因子失衡或調節異常都與疾病有著密切的關系。IL-18是一種很強的IFN-γ誘導劑,其生物學活性是通過調節IFN-γ的表達來提高機體細胞免疫水平。因此BolIL-18可望成為疫苗的一種重要的細胞免疫佐劑[13-15],在提高牛的抗感染力、增強滅活苗或基因工程苗免疫作用方面具有廣闊的應用前景。本實驗利用現有質粒為模板,將牛白細胞介素18與牛瑟氏泰勒蟲的p23表面蛋白基因通過重疊延伸拼接聚合酶鏈式反應(SOE-PCR)將兩段基因串連在一起,并連接到原核表達載體上,構建原核表達重組質粒。Western-blotting結果證實表達產物能被牛瑟氏泰勒蟲陽性血清所識別,證明表達產物具有較好的反應原性。
牛瑟氏泰勒蟲p23基因全長672 bp,具有完整的開放閱讀框,編碼223個氨基酸,N末端有28個氨基酸殘基的信號肽序列,疏水區多纈氨酸,C末端是跨膜區域,69~71位有一個潛在的N糖基化位點,為ASn-Ile-Ser,p23是單拷貝基因,并有等位基因。化學組成分析酪氨酸含量較多,彌補了p23作為抗原蛋白分子量較小的不足,且親水性較強,膜表面抗原決定簇豐富,具有疫苗候選分子的基礎。
在進行原核細胞表達時,多以融合蛋白為表達形式作為載體,這樣既可防止宿主菌的蛋白醇對所表達目的蛋白的降解,也有利于表達蛋白的純化,最終保護目的蛋白的活性。故本試驗選用pET-28a為載體,可將外源蛋白表達在細胞質內,在N端和C端都能加His-tag,pET-28a沒有本身溶解性髙的多肽融合蛋白,也沒有催化二硫鍵形成的酶融合蛋白,而且不含信號肽序列,這為純化目的蛋白提供了有利條件。同時pET-28a上的大部分酶切位點都不會造成閱讀框移位。
基于上述研究,本實驗將IL-18基因和p23基因融合在一起,成功構建了雙基因融合的基因工程菌株,并探討p23和IL-18雙基因融合產物作為亞單位疫苗成分的可行性[16],為探索牛瑟氏泰勒蟲雙基因工程疫苗奠定了理論依據。
[1]蔣金書.動物原蟲病學[M].北京:中國農業大學出版社,2001.
[2]Schnittger Leonhard,Katzer Frank,Biermann Reinhild,et al.Characterization of a polymorphic Theileria annulata surface protein(TaSP)closely related to PIM of Theileria parva:implications for use in diagnostic tests and subunit vaccines[J].Molecularandbiochemicalparasitology, 2002, 120(2):247-256.
[3]Wen Zhong Zhuang,Chihiro Sugimoto,Shuichi Kubota,et al.Antigenic alteration in major piroplasm surface proteins of Theileria sergenti during infection[J].Veterinary Parasitology,1995,60(3/4):191-198.
[4]金春梅,許應天,張守發,等.牛瑟氏泰勒蟲P33主要表面蛋白基因的克隆與序列分析[J].中國預防獸醫學報,2007,(2):112-114.
[5]Zhang W,Sugimoto C,Matsuba T,et al.Analysis of antigentic and genetic of Theileria sergenti major piroplasm surface proteins[J].Net Med Sci,1994,56:469-473.
[6]Bokarewa M,Hultgren O.Is interleukin-18 useful for monitoring rheumatoid arthritis?[J].Scandinavian journal of rheumatology,2005,34(6):433-436.
[7]Yano Ai,Nakao Kazushi,Sarai Ai,et al.Elevated serum interleukin-18 levels might reflect the high risk of hospitalization in patients on peritoneal dialysis[J].Nephrology(Carlton ,Vic.),2005,10(6):576-582.
[8]Thong-Ngam D,Tangkijvanich P,Lerknimitr R,et al.Diagnostic role of serum interleukin.1 8 in gastric cancer patients[J].World J Gastroenterol,2006,12(28):4473-4477.
[9]Shinoda Masahiro,Wakabayashi Go,Shimazu Motohide,et al.Increased serum and hepatic tissue levels of interleukin-18 in patients with fulminant hepatic failure[J].Journal of gastroenterology and hepatology,2006,21(11):1731-1736.
[10]Shoda L K,Zarlenda D S,Hirano A.Cloning of a cDNA encoding bovine interleukin-18 and analysis of IL-18 expression in macrophages and its IFN-gamma-inducing activity[J].J Interferon Cytokine Res,1999,(10):1169-1177.
[11]Nagata T,Ishikawa S,Shimokawa E,et al.High level expression and purification of bioactive bovine interleukin-18 using a baculovirus system[J].Vet Immunity,2002,87(8):65-72.
[12]劉 娟,劉明遠,于 錄,等.RV-G/LTB雙基因融合真核質粒表達載體的構建并在vero細胞中的表達[J].中國生物制品學雜志,2011,24(3):255-258.
[13]邢秀娟.牛IL-18真核表達載體的構建及其對FMD疫苗免疫增強作用的研究[D].泰安:山東農業大學,2008.
[14]葉麗萍,王春鳳.細胞因子免疫佐劑效應機制及其在獸醫臨床中的應用[J].中國畜牧獸醫2012,(11):196-199.
[15]劉建文,李月輝,王健春.基因疫苗及其免疫佐劑研究進展[J].中國獸醫雜志,2007,(3):41-43.
[16]孫平杰.牛瑟氏泰勒蟲p23表面蛋白基因與牛白細胞介素18基因在原核和真核細胞中的串聯表達[D].延邊:延邊大學,2012.
