調節性T細胞在poly I:C/D-GalN誘發的肝臟損傷中的免疫調節作用①
2014-11-27 11:15:54王黎麗沈際佳安徽醫科大學基礎醫學院病原生物學教研室合肥230001
中國免疫學雜志 2014年1期
宋 靜 王黎麗 侯 昕 沈際佳 (安徽醫科大學基礎醫學院病原生物學教研室,合肥230001)
急性肝損傷指各種原因引起的肝臟功能的損害,肝損傷是急性肝功能衰竭的基礎,嚴重或持續的肝損傷最終導致肝功能衰竭。其中,病毒性肝損傷并不是肝炎病毒在肝細胞內復制造成的直接損傷,而是病毒感染后機體的免疫反應造成的肝臟損傷。由于缺乏足夠的人類肝臟病變標本和人力實驗操作的困難以及倫理學難題,研究者們建立了模擬人類肝臟病理損傷的動物模型,用來探索肝臟疾病中的免疫致病機理。
1995年,Sakaguchi等人[1]發現在正常人和小鼠外周血及脾臟組織的CD4+T細胞中有一亞群持續高表達CD25分子(IL-2受體α鏈),去除該群細胞可誘導各種自身免疫性疾病的發生[2],據此表明該群細胞是一類重要的免疫調節細胞,將此類細胞命名為調節性T細胞。該細胞亞群具有顯著的免疫抑制作用,可通過不同途徑作用于多種靶細胞,從而對機體免疫應答進行精細的負調節。
本試驗中我們用病毒模擬物Polyinosinic-polycytidylic acid(poly I:C)和致敏劑D-galactosamine(D-GalN)聯合注射建立小鼠爆發性肝炎模型,該損傷主要由天然免疫細胞Kupffer細胞和自然殺傷(NK)細胞介導,天然免疫細胞釋放的 IFN-γ和TNF-α等炎性因子參與了肝損傷的發生發展[3]。鑒于該模型的肝臟免疫損傷可以自愈[3],而調節性T細胞是一群具有負調節作用的T細胞亞群,它不僅可以抑制效應T細胞的反應,而且可以緩解天然免疫細胞引發的病理損傷[4]。故我們推測調節性T細胞可能對該肝炎模型中天然免疫細胞有負向調節作用。
1 材料與方法
1.1 實驗動物 C57BL/6,SPF級,雄性,4周齡,購于安徽醫科大學實驗動物中心。C57BL/6遺傳背景的Rag1-/-小鼠,SPF級,購于南京大學模式動物遺傳中心。實驗動物由安徽醫科大學寄生蟲學教研室動物房飼養(SPF級,22℃,55%濕度,12 h白天/黑夜)。小鼠處理過程和實驗流程遵循實驗動物管理規范條例。
1.2 主要試劑 血清轉氨酶檢測試劑盒購于上海榮盛生物技術有限公司。Poly I:C、TRIzol、M-MLV、RT buffer和MDTT購于InvivoGen公司。D-GalN購于Sigma公司。Tag酶、SYBR Premix EX Tag購于大連寶生物工程有限公司。小鼠 IFN-γ和 TNF-α ELISA檢測試劑盒購于深圳達科為公司。Anti-FITC磁珠及分選儀購于Miltenyi Biotec公司。
1.3 方法
1.3.1 模型建立及材料收集 從小鼠尾靜脈處注射poly I:C(2 μg/只),同時給小鼠腹腔注射 DGalN(10 mg/只),建立小鼠急性肝損傷模型。于藥物注射后12、18、24和48 h,剖殺小鼠,收集血清,-80℃冰箱保存,用于血清ALT檢測及TNF-α和IFN-γ檢測。摘取小鼠肝臟,留取部分放入4%甲醛溶液中固定,石蠟包埋并切片,用于病理分析。余下肝臟放入-80℃冰箱保存,用于RT-PCR檢測。
1.3.2 血清轉氨酶(ALT)檢測 采集小鼠血清樣本后,按試劑盒說明書進行檢測。通過標準品吸光度值繪制標準曲線,然后計算各個樣品的值。
1.3.3 肝臟HE染色 取同一部位肝臟,福爾馬林固定,脫水,石蠟包埋,連續切片HE染色,顯微鏡下隨機選擇10個低倍視野觀察炎性細胞浸潤情況及肝組織細胞壞死情況。
1.3.4 實時熒光定量PCR檢測肝臟TNF-α和IFN-γ mRNA水平 用TRIzol試劑提取肝臟總RNA,逆轉錄成互補cDNA,以此為模板RT-PCR擴增片段。RT-PCR按SYBY Premis EX Tag II(TaKaRa Code DRR 041A)試劑盒說明書加樣,之后用ABI公司的StepOnePlus Real-time PCR儀檢測。目的基因引物序列 β-actin的上游引物:5'-TGG AAT CCT GTG GCA TCC ATG AAA-3',下游引物:5'-TAA AAC GCA GCT CAG TAA CAG TCC G-3'。TNF-α 的上游引物:5'-ACT GGC AGA AGA GGC ACT C-3',下游引物:5'-CTG GCA CCA CTA GTT GGT TG-3'。IFN-γ的上游引物:5'-AAC GCT ACA CAC TGC ATC T-3',下游引物:5'-GAG CTC ATT GAA TGC TTG G-3'。反應條件:95℃ 10 s,95℃ 15 s,60℃ 40 s,40 個循環進行擴增。采用ΔΔCt方法分析結果。
1.3.5 ELISA法檢測血清IFN-γ和TNF-α蛋白水平 采集小鼠血清樣本后,按試劑盒說明書進行檢測。通過標準品吸光度值繪制標準曲線,然后計算各個樣品的值。
1.3.6 免疫磁珠法分選CD25+細胞 殺鼠取脾臟,無菌分離脾臟單個核細胞,按照磁珠使用說明書分選CD25+單個核細胞和CD25-單個核細胞。
1.3.7 細胞轉輸 將免疫磁珠法分選出的CD25+細胞和 CD25-細胞按 1×106個/只鼠分別給Rag1-/-小鼠尾靜脈注射,然后立刻尾靜脈處注射poly I:C(2 μg/只),腹腔注射 D-GalN(10 mg/只)。
1.4 統計學處理 采用SPSS11.0軟件統計,數據以±s表示,兩組數據均值,選用獨立樣本t檢驗;組間數據比較,采用單因素方差分析。P<0.05為差異有統計學意義。
2 結果
2.1 野生型小鼠較Rag1-/-小鼠的肝臟炎癥損傷明顯加重
2.1.1 野生型小鼠和Rag1-/-小鼠poly I:C/DGalN注射后血清ALT檢測和肝臟H-E染色分析Rag1-/-小鼠在注射poly I:C/D-GalN后注射后18、24和48 h血清ALT水平均高于野生型小鼠(P<0.05),可見Rag1-/-小鼠在同等劑量的poly I:C/DGalN聯合注射后肝臟損傷比野生型小鼠更為嚴重。見圖1。肝臟H-E染色可見注射后18和24 h,與野生型小鼠相比Rag1-/-小鼠肝臟呈現嚴重的充血、肝細胞壞死以及炎癥細胞浸潤。注射后48 h,野生型小鼠肝臟損傷基本恢復正常,而Rag1-/-小鼠肝臟仍存在輕微損傷。見圖2所示。

圖1 野生型小鼠和Rag1-/-小鼠poly I:C/D-GalN注射后12、18、24和48 h檢測轉氨酶水平Fig.1 WT mice and Rag1 -/- mice were injected with poly I:C/D-GalN at 12,18,24 and 48 hours after injection,serum ALT levels were measured
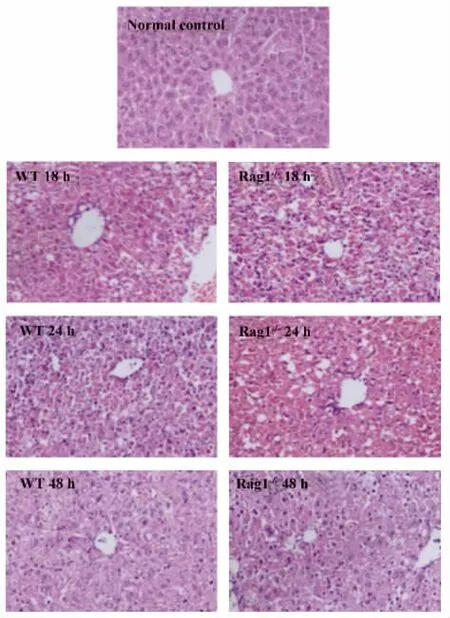
圖2 野生型小鼠和Rag1-/-小鼠poly I:C/D-GalN注射后18、24和48 h肝臟H-E染色(×100)Fig.2 WT mice and Rag1 -/- mice were injected with poly I:C/D-GalN at 18,24 and 48 hours after injection,liver samples from mice were analyzed by HE staining(original magnification×100)

圖3 RT-PCR檢測肝臟中 TNF-α和 IFN-γ的 mRNA水平Fig.3 TNF-α and IFN-γ mRNA expression of hepatic were measured by quantitative real-time RT-PCR
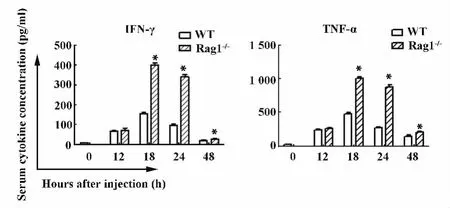
圖4 ELISA檢測血清中TNF-α和IFN-γ的蛋白水平Fig.4 Serum levels of TNF-α and IFN-γ were measured by ELISA
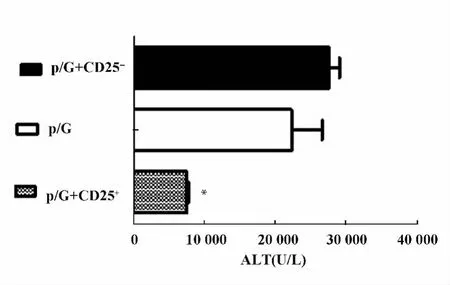
圖5 轉輸CD25+細胞,轉輸CD25-細胞以及未轉輸細胞的Rag1-/-小鼠poly I:C/D-GalN注射后18 h檢測轉氨酶水平Fig.5 Rag1-/- mice with transfer CD25+,transfer CD25-cells and did not transfer cells were injected with poly I:C/D-GalN,at 18 hours after injection,serum ALT levels were measured
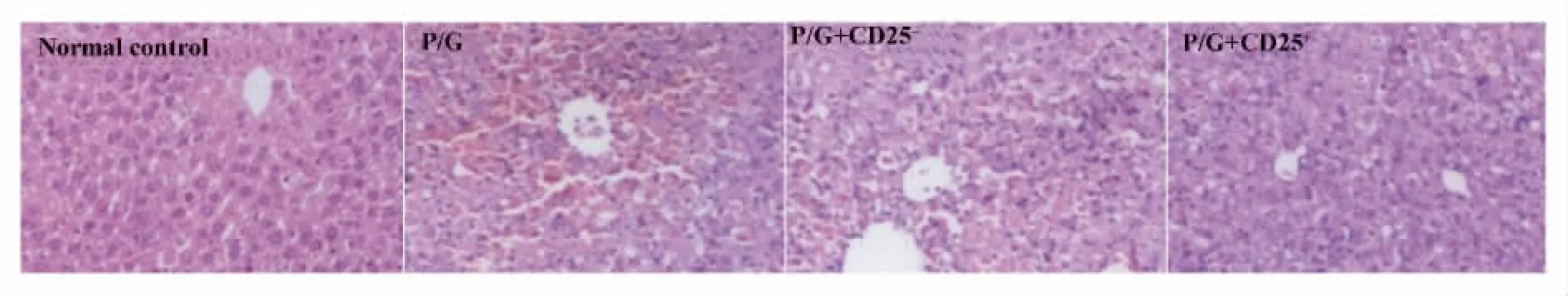
圖6 轉輸CD25+細胞,轉輸CD25-細胞以及未轉輸細胞的Rag1-/-小鼠poly I:C/D-GalN注射后18 h肝臟H-E染色(×100)Fig.6 Rag1-/- mice with transfer CD25+,transfer CD25-cells and did not transfer cells were injected with poly I:C/DGalN,at 18 hours after injection,liver samples from mice were analyzed by H-E staining(original magnification×100)
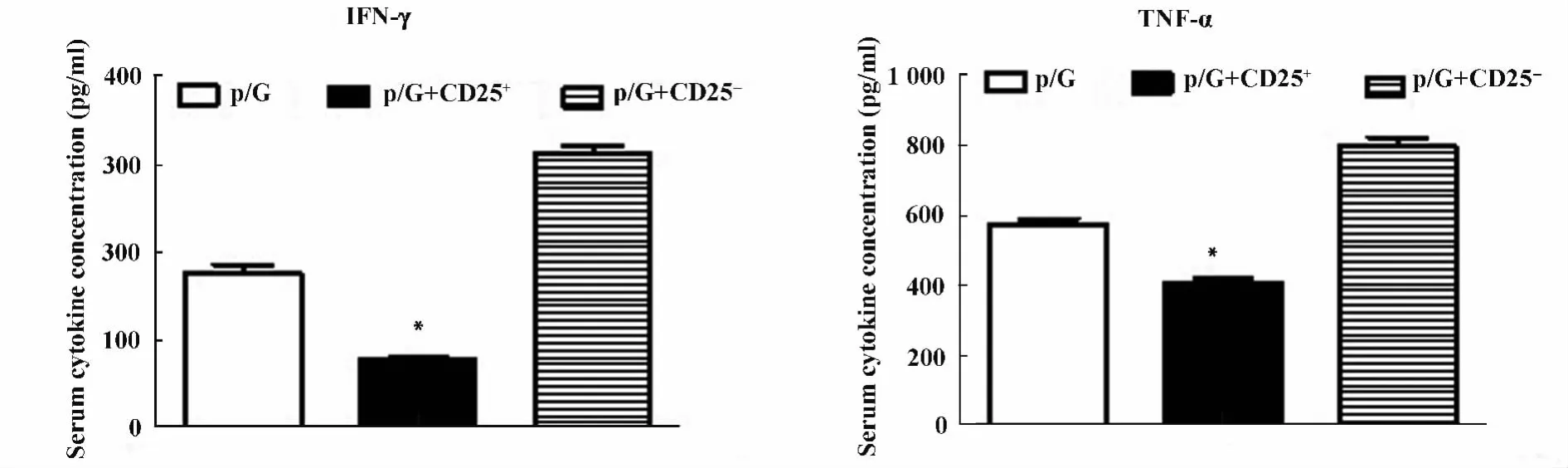
圖7 轉輸CD25+細胞,轉輸CD25-細胞以及未轉輸細胞的Rag1-/-小鼠poly I:C/D-GalN注射后18 h ELISA檢測血清中TNF-α和IFN-γ的蛋白水平Fig.7 Rag1-/- mice with transfer CD25+,transfer CD25-cells and did not transfer cells were injected with poly I:C/DGalN,at 18 hours after injection,serum levels of TNF-α and IFN-γ were measured by ELISA
2.1.2 野生型小鼠和Rag1-/-小鼠poly I:C/DGalN注射后肝臟炎癥損傷相關的炎癥因子TNF-α和IFN-γ mRNA水平和蛋白水平的動態表達Poly I:C/D-GalN 注射后12、18、24和48 h Rag1-/-小鼠肝臟IFN-γ的 mRNA水平明顯高于 WT小鼠(P<0.05),但是 Rag1-/-小鼠肝臟 TNF-α的 mRNA水平與野生型小鼠相比無統計學意義(P>0.05)。見圖3。同野生型小鼠相比,注射后18、24和48 h Rag1-/-小鼠血清 TNF-α和 IFN-γ含量明顯升高(P<0.05),而在注射后12 h兩種小鼠血清TNF-α和IFN-γ水平均無明顯差異(P>0.05)。見圖4。
2.2 過繼轉輸調節性T細胞減輕poly I:C/D-GalN誘導的損傷 我們通過免疫磁珠分選細胞系統純化得到脾臟CD25+單個核細胞純度大于90%,而且一半以上表達Foxp3,是調節性T細胞。將分離純化的脾臟CD25+單個核細胞過繼轉輸給Rag1-/-小鼠。同過繼轉輸 CD25-細胞和未轉輸細胞的Rag1-/-小鼠相比,轉輸CD25+細胞的Rag1-/-小鼠的血清ALT水平明顯降低(P<0.05),見圖5。肝臟H-E染色分析轉輸CD25-細胞和未轉輸細胞的Rag1-/-小鼠肝臟可見大量充血,肝細胞死亡以及大量炎性細胞浸潤。而過繼轉輸了CD25+細胞的Rag1-/-小鼠肝臟充血減輕,僅有小片的壞死區域以及少量的炎性細胞浸潤。見圖6。而且轉輸CD25+細胞的Rag1-/-小鼠的血清中炎性因子IFN-γ和TNF-α含量明顯下降。見圖7。
3 討論
本研究我們通過給小鼠注射poly I:C/D-GalN誘導急性肝臟損傷模型,重點探討了調節性T細胞在此模型中的免疫調節作用。在這個模型中我們發現沒有調節性T細胞存在的Rag1-/-小鼠由poly I:C/D-GalN誘導的肝臟損傷更為嚴重,而給Rag1-/-小鼠過繼轉輸調節性T細胞可以減輕poly I:C/DGalN誘導肝臟損傷。
根據前期的研究,我們確定了poly I:C/D-GalN可以誘導小鼠急性爆發性肝損傷模型,而且該損傷在2 d內基本自愈[3]。我們推測調節性T細胞可能限制了poly I:C/D-GalN誘導的急性肝臟損傷。因為調節性T細胞可以調節機體免疫反應,維持自身耐受和避免過度免疫反應。近年來的不少研究證實調節性T細胞在急性炎癥損傷如ConA誘導的急性肝炎[5],AdHBV 感染的急性肝炎[2],LPS 誘導的急性肺損傷[6]和腎臟缺血再灌注損傷[7]等早期均可被誘導而且具有免疫調節作用,限制炎癥損傷。于是我們首先比較了野生型小鼠和Rag1-/-小鼠在poly I:C/D-GalN注射后肝臟損傷輕重的不同,Rag1-/-小鼠在同等劑量的poly I:C/D-GalN注射后,肝臟炎癥損傷加重。同時Rag1-/-小鼠肝臟局部炎性因子IFN-γ,血清中TNF-α和IFN-γ分泌均高于野生型小鼠。將分離純化的CD25+細胞過繼轉輸給Rag1-/-小鼠,發現同過繼轉輸CD25-細胞以及未轉輸細胞的Rag1-/-小鼠相比轉輸調節性T細胞的小鼠肝臟炎癥損傷明顯減輕。以上結果均證實沒有調節性T細胞的存在,poly I:C/D-GalN誘導的急性炎癥反應加重,損傷時間延長,且與天然免疫細胞分泌的炎性因子TNF-α和IFN-γ有關,而轉輸了調節性T細胞后能夠明顯抑制炎性細胞因子的分泌和肝臟損傷。
調節性T細胞被認為主要通過抑制自身反應性T細胞的活化和擴增,得以限制變態反應疾病、感染、移植、移植物抗宿主反應和癌癥中的免疫反應強度,達到調節免疫應答的目的[8-11]。但是近年來越來越多的研究顯示調節性T細胞同樣可以通過抑制天然免疫細胞來發揮免疫調節作用。如調節性T細胞可以抑制NK細胞的細胞毒作用同時降低NK細胞分泌的IFN-γ[12]。調節性T細胞可以抑制中性粒細胞的功能,促進它們凋亡和死亡[13]。調節性T細胞還可以抑制巨噬細胞促炎因子反應[14]。腫瘤免疫中清除調節性T細胞可以促進依賴IFN-γ的巨噬細胞的聚集和活化從而抑制淋巴瘤細胞[15]。本研究中,poly I:C/D-GalN誘導的肝臟損傷是由天然免疫細胞介導,在缺失獲得性免疫細胞的Rag1-/-小鼠中依然能夠復制,并且損傷更加嚴重。而給Rag1-/-小鼠轉輸調節性T細胞后能夠明顯抑制肝臟損傷和TNF-α和IFN-γ的分泌。我們以前的研究表明TNF-α和IFN-γ分別是由肝臟中的巨噬細胞和NK細胞分泌。因此我們的結果提示調節性T細胞可以負向調節天然免疫細胞。
另外,有實驗證明調節性T細胞行使調節功能需要通過細胞-細胞接觸。這種接觸可能通過調節性T細胞表面的CTLA-4及其在效應性T細胞的配體相互作用而完成[16]。也有一些實驗發現調節性T細胞可以通過分泌免疫抑制細胞分子IL-10和TGF-β以及誘導其他細胞分泌這些細胞因子來發揮調節作用[6,17-19]。在本研究模型中究竟調節性T細胞如何發揮免疫調節作用來抑制poly I:C/D-GalN誘導的急性爆發性肝損傷還需進一步研究。
綜上所述,我們的實驗結果顯示poly I:C/DGalN誘導的急性肝損傷伴隨著調節性T細胞的顯著上升。調節性T細胞在poly I:C/D-GalN誘導的肝損傷中對限制肝臟免疫損傷起了重要作用。目前急性肝損傷治療并無特效的藥物治療,多以支持治療為主,調節性T細胞對肝臟免疫損傷的抑制作用可以為急性肝損傷的臨床治療、藥物研發開辟新的視角。
[1]Sakaguchi S,Sakaguchi N,Asano M,et al.Immunologic self-tolerance maintained by actived T cells expressing IL-2 receptor alpha-chains(CD25)breakdown of a single mechanism of self-tolerance causes various auto-immune diseases[J].J Immunol,1995,155(3):1151-1164.
[2]Leonhard Stross,Johannes Gunther,Georg Gasteiger,et al.Foxp3+regulatory T cells protect the liver from immune damage and compromise virus control during acute,experimental hepatitis B virus infection[J].Hepatology,2012,56(3):873-883.
[3]Hou X,Zhou RB,Wei HM,et al.NKG2D-retinoic acid early inducible-1 recognition between natural killer cells and kupffer cells in a novel murine natural killer cell-dependent fulminant hepatitis[J].Hepatology,2009,49(3):940-949.
[4]Fehervari Z,Sakaguchi S.Development and function of CD4+CD25+regulatory T cells [J].Nat Immunol,2004,16(2):203-208.
[5]Wei HX,Chuang YH,Li BF,et al.CD4+CD25+Foxp3+regulatory T cells protect against T cell-mediated fulminant hepatitis in a TGF-β dependent manner in mice[J].J Immunol,2008,181(10):7221-7229.
[6]D’Alessio FR,Kenji Tsushima,Aggarwal NR,et al.CD4+CD25+Foxp3+Tregs resolve experimental lung injury in mice and are present in humans with acute lung injury[J].J Clin Invest,2009,119(10):2898-2913.
[7]Gilbert R Kinsey,Rahul Sharma,Liping huang,et al.Regulatory T cells suppress innate immunity in kidney ischemia-reperfusion injury[J].J Am Soc Nephrol,2009,20(8):1744-1753.
[8]Viglietta V,Baecher-Allan C,Weiner HL,et al.Loss of functional suppression by CD4+CD25+regulatory T cells in patients with multiple sclerosis[J].J Exp Med,2004,199(7):971-979.
[9]Aandahl EM,Michaelsson J,Moretto WJ,et al.Human CD4+CD25+regulatory T cells control T-cell responses to human immunodeficiency virus and cytomegalovirus antigens [J].J Virol,2004,78(5):2454-2459.
[10]Tonkin DR,Kathryn Haskins.Regulatory T cells enter the pancreas during suppression of type 1 diabetes and inhibit effector T cells and macrophages in a TGF-β-dependent manner[J].Eur J Immunol,2009,39(5):1313-1322.
[11]Hisaeda H,Maekawa Y,Iwakawa D,et al.Escape of malaria parasites from host immunity requires CD4+CD25+regulatory T cells[J].Nat Med,2004,10(11):29-30.
[12]Hao Zhou,Li-juan Chen,Yong You,et al.Foxp3-transduced polyclonal regulatory T cells suppress NK cell functions in a TGF-β dependent manner [J].Autoimmunity,2010,43(4):299-307.
[13]Przemyslaw Lewkowicz,Natalia Lewkowicz,Andrzej Sasiak,et al.Lipopolysaccharide-activated CD4+CD25+T regulatory cells inhibit neutrophil function and promote their apoptosis and death[J].J Immunol,2006,177(10):7155-7163.
[14]Li M,Lin J,Wang ZX,et al.Oxidized low-density lipoproteininduced proinflammatory cytokine response in macrophages are suppressed by CD4+CD25+Foxp3+regulatory T cells through downregulating toll like receptor 2-mediated activation of NF-κB[J].Cell Physiol Biochem,2010,25(6):649-656.
[15]Galani IE,Marco Wendel,Ana Stojanovic,et al.Regulatory T cells control macrophage accumulation and activation in lymphoma[J].Int J Cancer,2010,127(5):1131-1140.
[16]von Boehmer H.Mechanisms of suppression by suppressor T cells[J].Nat Immunol,2005,6(4):338-344.
[17]Asseman C,Mauze S,Leach MW,et al.An essential role for interleukin 10 in the function of regulatory T cells that inhibit intestinal inflammation[J].J Exp Med,1999,190(7):995-1004.
[18]Nakamura K,Kitani A,Fuss I,et al.TGF-beta 1 plays an important role in the mechanism of CD25+CD4+regulatory T cells activity in both humans and mice[J].J Immunol,2004,172(2):834-842.
[19]Piccirillo CA,Letterio JJ,Thornton AM,et al.CD25+CD4+regulatory T cells can mediate suppressor function in the absence of transforming growth factor beta 1 production and responsiveness[J].J Exp Med,2002,196(2):237-246.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中老年保健(2021年3期)2021-08-22 06:50:04
天津醫科大學學報(2021年2期)2021-03-29 05:31:08
現代臨床醫學(2021年1期)2021-01-26 00:56:02
學苑創造·A版(2020年9期)2020-10-13 09:41:02
海峽科技與產業(2016年3期)2016-05-17 04:32:12
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
