胃癌細胞株MGC803中LTF基因的表達及其啟動子甲基化相關性研究
2015-01-18 07:47:21陳旭艷諸葛小菊陳仁聘黃智銘
浙江醫學 2015年12期
關鍵詞:胃癌
陳旭艷 諸葛小菊 陳仁聘 黃智銘
胃癌細胞株MGC803中LTF基因的表達及其啟動子甲基化相關性研究
陳旭艷 諸葛小菊 陳仁聘 黃智銘
目的鉬檢測胃癌細胞株MGC803內乳鐵蛋白(LTF)基因的甲基化和表達情況,探討LTF的DNA啟動子區甲基化異常與其在胃癌內表達的相關性。方法鉬用亞硫酸氫鹽測序PCR方法檢測MGC803啟動子區甲基化狀態,使用real-time PCR和Western b lot分別檢測LTF在MGC803內的mRNA和蛋白表達水平。同時使用去甲基化劑5-aza-CdR處理胃癌細胞株,觀察給藥前后的DNA甲基化狀態和mRNA表達情況。結果 MGC803啟動子甲基化率達59.8%,使用5-aza-CdR處理的兩組(2μmol/L組和10μmol/L組)和空白對照組甲基化率差異、LTF的mRNA表達差異均有統計學意義(均P<0.05),并且隨著MGC803啟動子甲基化程度的下降,其mRNA和蛋白表達增加;而不同濃度藥物處理組間差異均無統計學意義(均P>0.05)。結論 LTF基因在胃癌細胞株MGC803內表現為較高甲基化狀態,而且LTF的表達和其啟動子區甲基化情況相關,使用去甲基化劑能逆轉DNA的甲基化情況從而使其m RNA重新表達。
LTF基因 胃腫瘤 5-雜氮脫氧胞苷 DNA甲基化 CpG島
胃癌發病過程受到多基因、多因素的影響。抑癌基因的表觀遺傳學改變是在胃癌癌變演變過程早期非常關鍵的因素之一。表觀遺傳學變化主要包含DNA異常甲基化,雜合性丟失(loss of Heterozygosity,LOH)以及組蛋白修飾等三大方面[1]。Lund等[2]證實DNA甲基化是腫瘤抑制基因(tumor suppressor gene,TSG)失活機制中除基因缺失和基因突變這兩大重要因素外的第三重要機制,有時候甚至越過前兩者成為TSG沉默的唯一機制。
分子遺傳學研究發現,在多種腫瘤內中染色體3p21區存在高頻基因缺失[3],有學者通過基因組雜交技術發現胃癌里3p21-23的缺失率為22%[4]。乳鐵蛋白(lactotransferrin,LTF)基因,作為轉鐵蛋白家族的一員,亦位于人類染色體3p21.3區域,其在鼻咽癌、肺癌、前列腺癌等多種腫瘤及相應的腫瘤細胞株呈現表達降低或缺失的狀態,有研究顯示這與該基因啟動子區域的異常甲基化高度相關[5-8]。5-Aza-2′-deoxycytidine(5-aza-CdR),能結合DNA甲基轉移酶I并使其失活,逆轉抑癌基因的高甲基化狀態,重新激活基因表達。而目前對于LTF基因在胃癌內的研究尚少,因此本實驗選擇胃癌細胞株MGC803,觀察LTF基因的表達和其啟動子甲基化狀態,旨在研究胃癌細胞株內LTF基因表達水平和啟動子異常甲基化相關性,為胃癌臨床診斷治療提供一個新的可能靶點。
1 材料和方法
1.1 材料 人胃腺癌細胞株MGC803(中國上海科學院細胞庫),胎牛血清(杭州四季青生物工程材料有限公司),PRMI1640、PBS、青/鏈霉素(100U/ml)購于美國Gibco公司;5-aza-CdR(美國Sigma公司),用培養基充分溶解配成適量濃度的母液,分裝后存于-80℃備用;TRIzol(美國Invitrogen公司);逆轉錄試劑盒,real-time PCR試劑盒(SYBRRGreen Realtime PCRMasterMix,日本TOYOBO公司);亞硫酸氫鈉修飾試劑盒(美國ZYMO公司),Ex Taq HSDNA聚合酶、pMD 19-T Vecto(r日本TaKaRa公司)。抗體anti-Lactoferrin(美國Abcam公司,77 kDa,1∶1 000#ab10110),β-actin antibody(中國碧云天公司,42kDa,1∶1 000)。熒光定量PCR儀(型號CFX96,美國BIO-RAD公司)。
1.2 方法
1.2.1 細胞培養和5-aza-CdR藥物處理 SGC7901細胞用含10%胎牛血清的PRMI-1640培養液放置在37℃、5%C O2的細胞培養箱內培養。以1×105均勻鋪入6孔板內,生長過夜。饑餓處理16 h,然后給予0μmol/L(空白對照組)、2μmol/L、10μmol/L的5-aza-CdR藥物處理(分別為2μmol/L組和10μmol/L組),24h換液1次,培養5d后處理細胞。
1.2.2 real-time PCR檢測藥物干預前后LTF基因mRNA表達 取對照和藥物處理組后回收細胞,用TRIzol提取細胞總RNA。約取1μg總RNA根據逆轉錄試劑盒說明書反應得到20μl體系的cDNA。以得到cDNA為模板,進行real-time PCR反應。引物由上海生工公司設計并合成,用GAPDH作為內參,引物序列分別為上游5′-GACTCATGACCACAGTCCATGC-3′,下游5′-AGAGGCAGGGATGATGTTCTG-3′,擴增產物長度113 bp。LTF引物序列分別為:上游5′-TACAAATCCCAACAAAGCAGTG-3′,下游5′-CTTCCACAGGTCTATCCACACA-3′,產物大小約58bp。20μl PCR體系包括:SYBRRGreen Realtime PCR Master Mix 10μl,Plus solution 2μl,PCR Forward Prime(r10μmol/L)1.2μl,PCR Reverse Primer(10μmol/L)1.2μl,模板 cDNA 2μl,ddH2O 3.6μl。反應條件是95℃預變性30s,95℃5s、62℃10s、72℃15s進行40個循環。擴增結束進行溶解曲線分析來辨別是否有非特異性擴增。實驗重復3次。數據用2-ΔΔCt法分析。
1.2.3 Western blot檢測藥物干預前后LTF基因蛋白表達 回收空白對照組和藥物處理組細胞,加入適量的蛋白抽提 cell lysis buffer(每孔50μg)裂解細胞,然后用BCA法測定蛋白質的濃度。用12%SDS-聚丙烯酰胺凝膠進行電泳,將蛋白用半干式電轉移法轉至硝酸纖維膜上,用5%脫脂牛奶TBST封閉硝酸纖維膜,室溫下搖2 h。與一抗anti-Lactoferrin(77 kDa)結合(以TBS稀釋,濃度依抗體而定),4℃搖過夜,TBST洗4次,每次5min,用β-actin antibody(42kDa)作為內參。加入二抗(以TBS稀釋,濃度依抗體而定),室溫搖1 h,TBST洗4次,每次5min。用碧云天公司的ECL試劑盒進行化學發光檢測。X線片壓片、顯影、定影。
1.2.4 亞硫酸氫鹽修飾DNA和BSP反應 取對照和藥物處理組后回收細胞,按DNA提取試劑盒提取胃癌細胞株的基因組DNA。然后按甲基化修飾試劑盒進行亞硫酸氫鈉修飾,使未甲基化的胞嘧啶(C)轉化成胸腺嘧啶(T),而甲基化的Cm不變。以修飾后基因組DNA為模板,用Ex Taq HSDNA聚合酶進行LTF基因啟動子擴增。LTF特異性引物參考文獻[5],分別是上游5′-TTGAGATTAGAGTTGGGATAGGG-3′,下游:5′-CCCCCAAACACCTACACTCA-3′,產物長度為397bp。其反應體系為50μl,包括:TaKaRa Taq HS 0.25μl,10×PCR Buffer 5μl,dNTPMixture 4μl,上游引物(10μmol/L)1μl,下游引物(10μmol/L)1μl,模板DNA 1μl(≤50ng),ddH2O 37.75μl。反應條件為95℃變性15min,94℃ 30s、56℃30s、72℃30s進行35次循環,然后72℃延伸7min。將獲得的PCR產物用1.5%瓊脂糖凝膠電泳,凝膠圖像分析系統記錄數據。
1.2.5 挑選陽性克隆和測序 于紫外燈下切取含目的DNA的條帶,按DNA凝膠回收試劑盒說明書對PCR產物進行回收純化。取回收純化的DNA 4.5μl,與pMD 19-T Vector 0.5μl,solution I 5μl配成10μl體系,輕輕混勻后16℃連T載體12h。取2.5μl連接產物與25μl的大腸桿菌感受態冰浴30min,42℃水浴90s,冰上放置3min,加入不含氨芐青霉素的LB培養基600μl,37℃,250r/min振搖45min,離心將菌液濃縮為15μl,并涂于氨芐(100μg/ml)抗性的LB平板上,先正面向上放置平板1h風干,翻轉倒置平板37℃培養過夜,12~16h后即可長出菌落。隨機挑選15個單克隆菌落,放在500μlLB液體培養基(含有氨芐青霉素100μg/ml),搖渾濁菌液。用普通PCR鑒定陽性重組子,取1μl搖渾的菌液做PCR模板,2×Taq酶MIX 5μl,上下游通用引物各0.2μl,ddH2O 3.6μl構成10μl PCR體系進行擴增反應。用1.5%瓊脂糖凝膠電泳鑒定結果。最后送檢10個克隆于上海生工公司測序。
1.3 統計學處理 采用 SPSS13.0統計軟件,realtime PCR結果和總體甲基化率組間比較采用Oneway ANOVA分析,組間兩兩比較采用LSD-t法。CpG位點甲基化率比較采用行×列表χ2檢驗。
2 結果
2.1 胃癌細胞株MGC803 LTF基因的mRNA表達和啟動子甲基化率 見圖1。
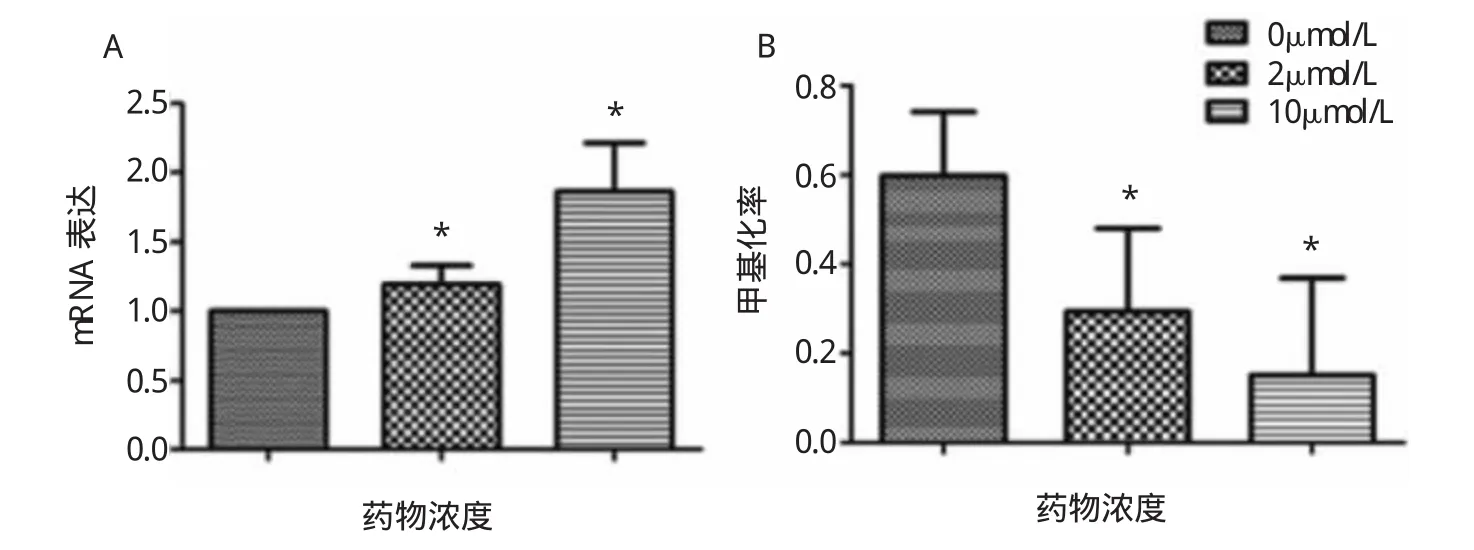
圖1 3組LTFmRNA表達(A)和啟動區甲基化率(B)的比較(與空白對照組比較,*P<0.05)
由圖1可見,不同濃度5-aza-CdR處理胃癌細胞株MGC803后,2μmol/L組和10μmol/L組其mRNA表達較空白對照組明顯增加,差異均有統計學意義(均P<0.05),但2μmol/L組和10μmol/L組間比較差異無統計學意義。甲基化率空白對照組為59.82%,2μmol/L組為29.46%,10μmol/L組為15.17%(P<0.05),但2μmol/L組和10μmol/L組間比較差異無統計學意義(P>0.05)。
2.2 胃癌細胞株MGC803 LTF基因蛋白表達 見圖2。
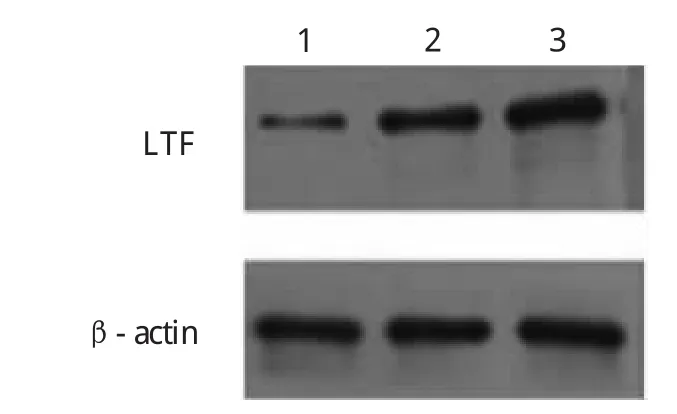
圖2 5-aza-CdR藥物處理前后LTF的蛋白表達(1、2、3分別代表空白對照組、2μmol/L組、10μmol/L組)
由圖2可見,藥物處理前后LTF基因蛋白表達空白對照組較低,兩個不同濃度藥物組均較空白對照組明顯增加。
2.3 胃癌細胞株MGC803藥物處理前后LTF基因甲基化情況 見圖3~5。
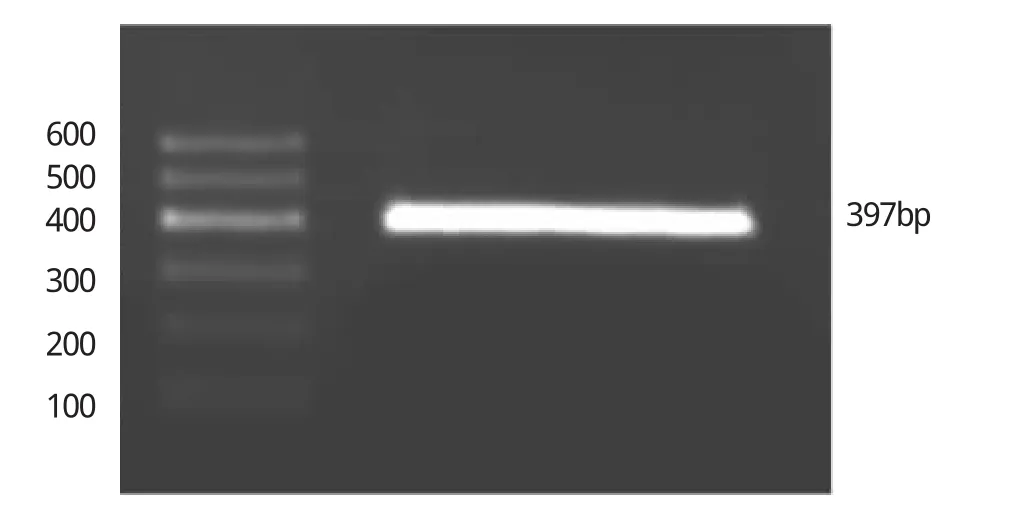
圖3 LTF基因的BSP法電泳圖結果
由圖3可見,目的條帶397bp清晰。
由圖4可見,所選陽性單克隆概率達83.3%(5/6)。
由圖5可見,其中3,4,5,6,11,12,13號CpG位點甲基化程度超過50%,呈高甲基化狀態。而用5-aza-CdR處理后,與空白對照組相比,發現14號位點甲基化情況變化不大。3、4、5、6、10、11、12、13號這8個CpG位點甲基化率較空白對照組明顯降低,均有統計學意義(均P<0.05)。
3 討論
LTF基因位于染色體短臂3p21.3的CER1區,編碼包含17個外顯子長約2.4kb的mRNA,翻譯一個分子質量約為80kD的鐵結合蛋白,命名為乳鐵蛋白(lactoferrin,LF)[9]。該基因在多種惡性腫瘤內表現為候選的抑癌基因。
LTF基因在鼻咽癌,肺癌,前列腺癌等多種腫瘤及相應的腫瘤細胞株呈表達降低或缺失的狀態,有研究顯示這種現象與該基因啟動子區域的甲基化異常高度相關[5-8]。為研究LTF基因啟動子甲基化對LTF在胃癌里表達的影響,我們使用BSP方法來檢測胃癌細胞株的甲基化情況。BSP法是公認的分析DNA甲基化的金標準[10-11]。本研究結果顯示LTF基因在胃癌細胞株MGC803內是呈高甲基化的(總體甲基化率是59.82%),這與學者在鼻咽癌和肺癌里的報道一致[6-7]。去甲基化劑5-aza-CdR作用后隨著藥物處理組LTF基因啟動子區總體甲基化程度下降,其mRNA重新表達。這些結果進一步證明LTF基因啟動子區異常高甲基化是引起LTF基因在胃癌組織和細胞株里沉默的關鍵表觀遺傳學機制,同時這種“疾病狀態”是可以被去甲基化劑逆轉的。但是兩個藥物處理組間的甲基化率和mRNA表達差異無統計學意義,這可能和5-aza-CdR對BGC823的去甲基化作用達到平臺期相關。該段DNA啟動子區含有14個CpG島,CpG島是一段長約1kb的富含飾CpG和GpC序列的DNA結構。進一步分析發現其中3,4,5,6,11,12,13號CpG位點甲基化程度超過50%,呈高甲基化狀態。而用5-aza-CdR處理后,與空白對照組相比,發現14號位點甲基化情況變化不大。3,4,5,6,10,11,12,13號這8個CpG位點甲基化率較空白對照組明顯降低,且有統計學意義(P<0.05)。我的猜測這些位點和LTF在胃癌的表達缺失及逆轉其表達可能相關。
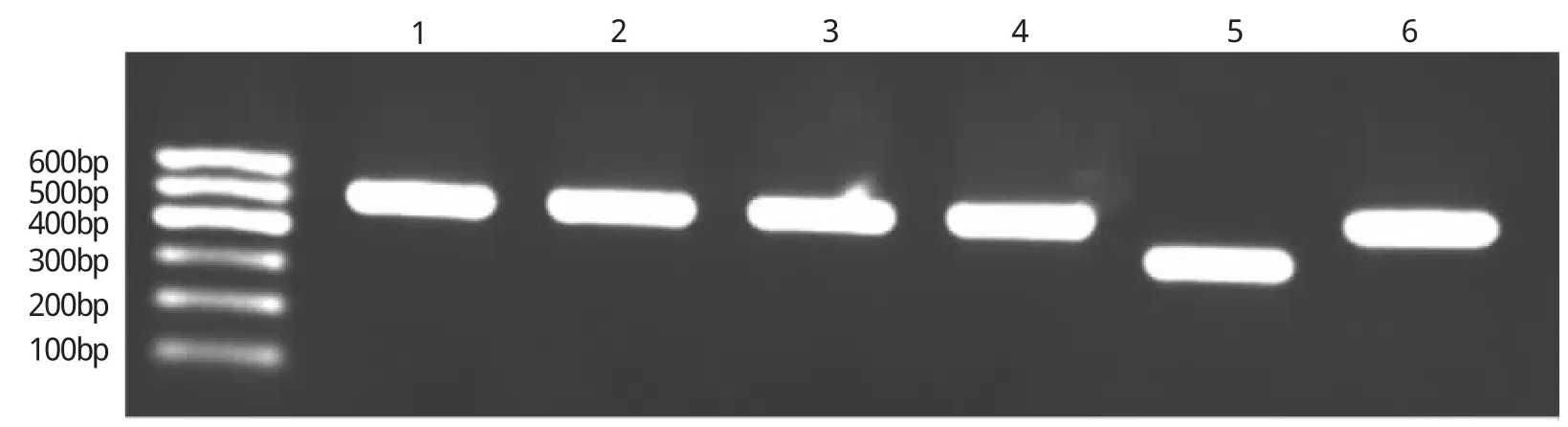
圖4 LTF甲基化后目的DNA連T載體挑選陽性克隆電泳圖(1,2,3,4,6為陽性克隆,成功連接T載體,509 bp;5為陰性克隆,未連接T載體,397 bp)
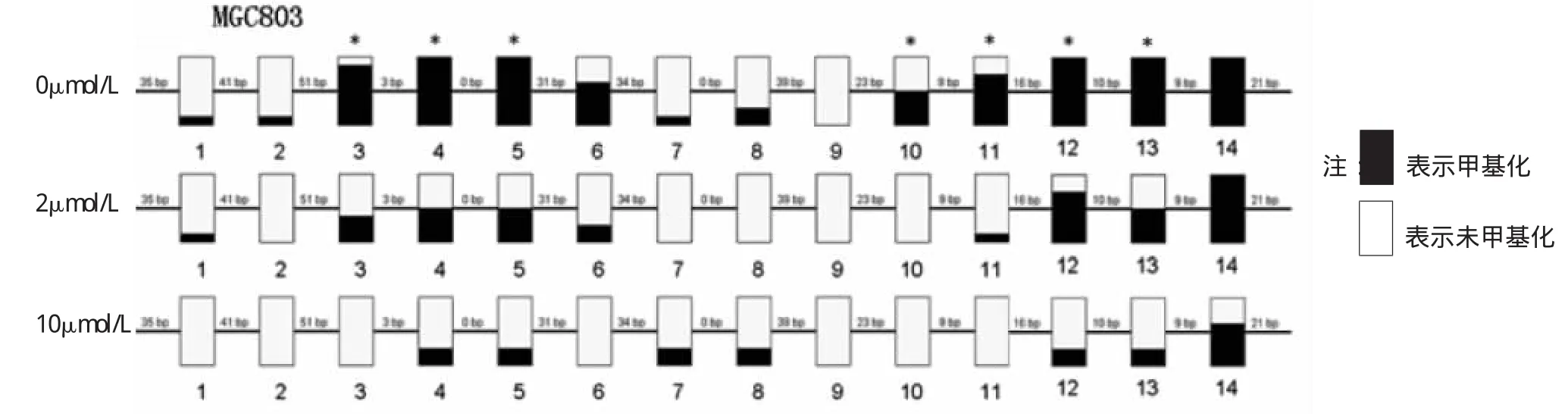
圖5 3組LTF基因啟動子區14個CpG島的甲基化率改變(與空白對照組比較,*P<0.05)
大量研究證明LTF在體內外實驗中有抗腫瘤作用[12-14],但是其機制目前并不明確。大量文獻報道在乳腺癌,頭頸部腫瘤和其他腫瘤細胞內,LTF蛋白可以激活細胞周期相關蛋白,進而阻滯惡性腫瘤細胞從G1期到S期的過渡,最終抑制腫瘤細胞的增殖[15-16]。這提示LTF蛋白對這些腫瘤有化學預防作用。有些研究認為LTF的蛋白能調節細胞周期相關蛋白,如細胞周期蛋白cyclin、細胞周期蛋白依賴性激酶(cyclin-dependent kinase,CDK)、細胞周期蛋白依賴性激酶抑制劑(cyclin dependent kinase inhibitor,CKI)和抑癌基因(p53和Rb)的表達,進而影響細胞周期的整個進程[17]。但是也有學者持不同意見,認為LTF蛋白補給治療能調節MAPK(mitogen-activated protein kinase)通路卻不會影響p53的表達[18]。Oh等[19]證實LTF蛋白參與激活NF-kB(nuclear factor-κB,NF-kB)信號通路,進而激活NF-kB的相關靶基因,同時激活的NF-kB能抑制p53的轉錄,p53能反向調節下游基因mdm2和p21的表達,最終遏制腫瘤細胞的生長。Xu等[20]報道LTF在SGC7901里能夠抑制蛋白激酶的活化,調節其下游蛋白的磷酸化然后誘導腫瘤細胞凋亡。另有學者報道LTF在結腸癌里促進細胞凋亡的機制與激活Fas信號通路,增強caspase-8和caspase-3表達相關[21]。
在本研究中,我們發現LTF基因在胃癌里是低表達的,而且與LTF啟動子甲基化高度相關。DNA甲基化作為表觀遺傳學重要機制之一,其本身優勢在于不涉及基因序列的改變就可以調控基因的表達,并且能夠被甲基化轉移酶抑制劑逆轉的。5-aza-CdR,作為一種甲基化轉移酶,已成功應用于白血病的臨床治療[22],這對其他機制和抑癌基因甲基化異常相關的腫瘤來說是個成功的鼓舞。LTF可作為胃癌內的候選抑癌基因,由于LTF在胃癌內表達和基因甲基化機制相關,為未來可能
通過甲基化轉移酶抑制劑來進行腫瘤的預防和早期治療提供理論依據。
[1]Choi IS,Wu T T.Ep igenetic alterations in gastric carcinogenesis [J].CellRes,2005,15(4):247-254.
[2]Lund A H,van Lohuizen M.Ep igenetics and cancer[J].Genes Dev,2004,18(19):2315-2335.
[3]Hesson L B,Cooper W N,Latif F.Evaluation of the 3p21.3 tumour-supp ressorgene c luster[J].Oncogene,2007,26(52):7283-7301.
[4]Sakakura C,MoriT,Sakabe T,etal.Gains,losses,and amp lifications ofgenom icmaterials in p rimary gastric cancers analyzed by comparative genom ic hyb rid ization[J].Genes Chrom osomes Cancer,1999,24(4):299-305.
[5]YiH M,LiH,Peng D,et al.Genetic and ep igenetic alterations o f LTFat3p21.3 in nasopharyngealcarcinoma[J].OncolRes,2006,16 (6):261-272.
[6]Zhang H,Feng X,Liu W,etal.Underlying m echanism s for LTF inactivation and its functionalanalysis innasopharyngea lcarcinom a celllines[J].JCellBiochem,2011,112(7):1832-1843.
[7]Iijim a H,Tom izawa Y,IwasakiY,etal.Genetic and ep igenetic inactivation of LTF gene at3p21.3 in lung cancers[J].Int JCancer, 2006,118(4):797-801.
[8]Shaheduzzaman S,Vishwanath A,Furusato B,et al.Silencing o f Lactotransferrin exp ression bym ethylation in p rostate cancerp rog ression[J].Cancer BiolTher,2007,6(7):1088-1095.
[9]No jim a Y,SuzukiY,IguchiK,etal.Development of poly(ethylene g lycol)con jugated lactoferrin for oraladm inistration[J].Bioconjug Chem,2008,19(11):2253-2259.
[10]Clark S J,Harrison J,PaulC L,eta l.High sensitivitym app ing of methylated cytosines[J].Nuc leic Acids Res,1994,22(15):2990-2997.
[11]Frommer M,McDonald L E,Millar D S,et al.A genom ic sequenc ing p rotocol that yields a positive d isp lay of 5-m ethylcytosine residues in ind ividualDNA strands[J].Proc NatlAcad Sci U SA,1992,89(5):1827-1831.
[12]Tsuda H,Kozu T,Iinuma G,et al.Cancer p revention by bovine lacto ferrin:from animal stud ies to hum an trial[J].Biometals, 2010,23(3):399-409.
[13]Yamada Y,Sato R,KobayashiS,etal.The antip roliferative effect o fbovine lactoferrin on caninem ammary g land tum or cells[J].J VetMed Sci,2008,70(5):443-448.
[14]Zhou Y,Zeng Z,Zhang W,etal.Lactotransferrin:a cand idate tum or supp ressor-Deficient exp ression in humannasopharyngeal carcinom a and inhibition of NPC cell p roliferation by modu lating the m itogen-activated p rotein kinase pathway[J].Int JCancer,2008,123(9):2065-2072.
[15]Wang J,LiQ,Ou Y,etal.Inhibition of tumor g row th by recom binant adenovirus containing human lactoferrinthrough inducing tumor cell apop tosis in m ice bearing EMT6 b reast cancer[J]. Arch Pharm Res,2011,34(6):987-995.
[16]Xiao Y,Monitto C L,Minhas K M,et al.Lacto ferrin down-regulates G1 cyc lin-dependentkinases during grow th arrestofhead and neck cancer cells[J].Clin Cancer Res,2004,10(24):8683-8686.
[17]Son H J,Lee SH,ChoiSY.Human lactoferrin controls the levelof retinob lastoma p rotein and its ac tivity[J].Biochem Cell Biol, 2006,84(3):345-350.
[18]Zhou Y,Zeng Z,ZhangW,etal.Lactotransferrin:a candidate tumor supp ressor-Deficientexp ression in humannasopharyngeal carcinoma and inhibition of NPC cell p roliferation by modulatingthem itogen-activated p rotein kinase pathway[J].Int JCancer,2008,123(9):2065-2072.
[19]Oh SM,Pyo CW,Kim Y,etal.Neutrophil lactoferrin up regulates the human p53 gene through induc tion ofNF-kappaB ac tivation cascade[J].Oncogene,2004,23(50):8282-8291.
[20]Xu XX,Jiang H R,LiH B,etal.Apop tosis ofstomach cancer cell SGC-7901 and regulation of Akt signaling wayinduced by bovine lac toferrin[J].JDairy Sci,2010,93(6):2344-2350.
[21]Fujita K,Matsuda E,Sekine K,et al.Lactoferrin enhances Fas exp ression and apop tosis in the co lon mucosa of azoxym ethane-treated rats[J].Carcinogenesis,2004,25(10):1961-1966.
[22]Kantarjian H,Oki Y,Garcia-Manero G,et a l.Results of a random ized study o f 3 schedules of low-dose decitabine inhigher-riskmyelodysp lastic synd rome and chronic myelom onocytic leukem ia[J].Blood,2007,109(1):52-57.
Expression of LTF gene and its promotor region methylation in human gastric cancer cell lines MGC803
CHEN Xuyan,ZHUGE Xiao-ju,CHEN Renpin,et al.Department of Gastroenterology,the First Affiliated Hospital of Wenzhou Medical University,Wenzhou 325000,China
Objective To investigate the exp ression of LTF(lactotransferrin)gene and its p romoter region in human gastric cancer cell line MGC803.Methods The exp ression of LTFm RNA and p rotein levels in MGC803 cells were detected by real-time PCR and Western b lot,respec tively;LTF p romotormethylation status was determ ined by Bisulfite genom ic sequencing PCR(BSP).MGC803 cells were treated w ith 5-aza-CdR to reversemethylation status,Results LTF p romotormethylation rate in MGC803 was 59.8%.Compared to control g roup,methylation status and m RNA exp ression of LTF in 5-aza-CdR treatment groups(2μmol/L and 10μmol/L)were reversed(P<0.05);however there was no d ifference between d ifferent treatmentg roups(P>0.05).Conclusion The LTFmethylation in p romoter region is high and LTFm RNA exp ression in MGC803 cells is low,5-aza-CdR can reverse themethylation status and up-regulate LTFm RNA exp ression.
Stomach neop lasms Lac totransferrin 5-aza-CdR DNAmethylation CpG island
2015-03-06)
(本文編輯:沈昱平)
溫州市科技局項目(Y20140275)
325000溫州醫科大學附屬第一醫院消化內科(陳旭艷、陳仁聘、黃智銘);溫州醫科大學附屬第三醫院消化內科(諸葛小菊)
黃智銘,E-mail:w yyyhzhim ing@126.com
猜你喜歡
昆明醫科大學學報(2022年1期)2022-02-28 07:43:36
昆明醫科大學學報(2021年5期)2021-07-22 07:32:22
基層中醫藥(2020年2期)2020-07-27 02:46:06
中國組織化學與細胞化學雜志(2016年3期)2016-02-27 11:15:35
中國衛生標準管理(2015年3期)2016-01-14 03:41:46
中外醫療(2015年18期)2016-01-04 06:51:55
吉林大學學報(醫學版)(2015年4期)2015-12-17 07:48:25
醫學研究雜志(2015年6期)2015-07-01 17:40:49
醫學研究雜志(2015年9期)2015-07-01 17:28:27
中國當代醫藥(2015年20期)2015-03-01 02:04:29
