細胞毒性T淋巴細胞相關蛋白4在日本血吸蟲免疫逃避中的作用及其機制研究
2015-02-14 08:08:58唐春蓮申志琴嚴玲霞劉曉宏
中國人獸共患病學報 2015年11期
唐春蓮,申志琴,嚴玲霞,劉曉宏
細胞毒性T淋巴細胞相關蛋白4在日本血吸蟲免疫逃避中的作用及其機制研究
唐春蓮,申志琴,嚴玲霞,劉曉宏
目的 探討細胞毒性T淋巴細胞相關蛋白4(CTLA-4)在日本血吸蟲免疫逃避中的作用及其機制。方法 雌性BALB/c小鼠隨機分成3組,即正常對照組、感染對照組和抗CTLA-4單克隆抗體(Anti-CTLA-4 mAb)組。各感染組每只小鼠經腹部皮膚感染日本血吸蟲尾蚴40條,感染2周后,Anti-CTLA-4 mAb組每只小鼠連續3 d腹腔注射300 μg anti-CTLA-4 mAb,對照組注射等體積的PBS。感染后6周殺鼠沖蟲,計數每只小鼠蟲荷及每克肝臟蟲卵數。流式細胞術檢測各組小鼠脾淋巴細胞中調節性T細胞(CD4+CD25+Tregs)百分比。ELISA法檢測各組小鼠脾淋巴細胞培養上清中細胞因子IFN-γ、IL-4、IL-5和IL-10的含量。肝組織石蠟切片HE染色觀察感染組小鼠肝臟蟲卵肉芽腫病理變化。結果 Anti-CTLA-4 mAb組減蟲率為18.99%,脾淋巴細胞中CD4+CD25+Tregs百分比顯著升高(P<0.05)。Anti-CTLA-4 mAb組脾細胞培養上清中細胞因子IFN-γ、IL-4、IL-5和IL-10的含量均高于感染對照組,肝臟蟲卵肉芽腫直徑較感染對照組明顯增大。結論 Anti-CTLA-4 mAb使用同時增強Th1型和Th2型免疫反應,有利于機體清除日本血吸蟲但以損傷機體為代價。研究結果提示宿主CTLA-4利于日本血吸蟲免疫逃避。
日本血吸蟲; 細胞毒性T淋巴細胞相關蛋白4; 免疫逃避; 細胞因子
Supported by the Schistosomiasis Control Project from Hubei Provincial Health and Family Planning Commission(No. WJ2015XB016) Corresponding author: Liu Xiao-hong, Email: 494527907@qq.com
血吸蟲病是一種常見的人獸共患寄生蟲病,全球76個國家至少23億人感染血吸蟲[1],我國主要是日本血吸蟲。寄生蟲具有免疫逃避功能能在宿主體內長期存在,其逃避機制可能與抗原偽裝、序列變異等相關[2]。新近研究證實,細胞毒性T淋巴細胞相關蛋白-4(cytotoxic T-lympnocyte-associated protein 4,CTLA-4)與病原體逃避宿主免疫攻擊有關,如HIV感染患者,感染利什曼原蟲后導致CD4+CTLA-4+調節性T細胞比例明顯增高,導致原蟲復發[3]。實驗性腦瘧原蟲模型中,FOXP3+細胞損傷了抗原特異性CD4+T細胞及CD8+T細胞反應,因此利于瘧原蟲逃避宿主免疫攻擊。而FOXP3+細胞調節的保護作用依賴于CTLA-4[4]。CTLA-4在日本血吸蟲免疫逃避中的作用尚無報道。本文采用CTLA-4 mAb,觀察其在日本血吸蟲免疫逃避中的作用及其機制。
1 材料與方法
1.1 實驗動物分組及處理 18只6~8周齡雌性BALB/c小鼠購自湖北省疾病預防控制中心,隨機分成3組,每組6只。即正常對照組、感染對照組、Anti-CTLA-4 mAb組。日本血吸蟲陽性釘螺(購自江西省寄生所)光照下逸出尾蚴,各感染組每只小鼠經腹部皮膚感染尾蚴40條。感染后2周,抗體組每只小鼠連續3 d腹腔注射300 μg anti-CTLA-4 mAb,對照組注射等體積的PBS。感染后6周剖殺所有小鼠。
1.2 試劑器材 調節性T細胞染色試劑盒試劑盒購自eBioscience公司;細胞因子IFN-γ、IL-4、IL-5、IL-10檢測試劑盒購自eBioscience公司;流式細胞儀(FACSCalibur型)購自美國Becton Dickinson公司。
1.3 蟲荷及每克肝臟蟲卵計數 日本血吸蟲感染后6周,剖殺各感染組小鼠。生理鹽水灌注沖蟲[5],計算各感染組每只小鼠蟲荷及減蟲率。稱取各感染組每只小鼠肝臟右葉0.5 g,研磨,5% KOH 20 mL 37 ℃消化2 h,取250 μL計數,重復4次取平均值,計算每克肝臟蟲卵數。剩余肝組織用于病理學檢測。
1.4 脾細胞流式細胞術檢測無菌取脾,根據文獻報道的方法[6]制備單個脾淋巴細胞懸液。以含10%胎牛血清的RPMI-1640培養液重懸,顯微鏡下計數淋巴細胞數量,調整其濃度為5×106/mL。流式細胞術染色使用小鼠調節性T細胞染色,其中CD4抗體、CD25抗體和Foxp3抗體分別以異硫氰酸熒光素、別藻青蛋白和藻紅蛋白標記。流式細胞儀檢測脾淋巴細胞中CD4+CD25+Tregs的百分比。
1.5 脾細胞培養上清中細胞因子檢測 無菌取脾,研磨,制備脾淋巴細胞懸液并調整細胞濃度至5×106/mL,置37 ℃、5% CO2培養箱培養72 h,收集上清液。雙抗體夾心ELISA法檢測脾細胞培養上清中細胞因子IFN-γ、IL-4、IL-5和IL-10的水平,根據標準曲線計算待測樣品中特定細胞因子含量(pg/mL)。
1.6 病理學檢測 用于病理學檢測的肝組織經10%福爾馬林固定后,石蠟包埋。切片,厚度約5 μm,常規HE染色,觀察各感染組小鼠肝臟蟲卵肉芽腫的變化。
2 結 果
2.1 Anti-CTLA-4 mAb對小鼠體內蟲荷及每克肝臟蟲卵數的影響 Anti-CTLA-4 mAb使用組蟲荷及每克肝臟蟲卵數明顯低于感染對照組(P<0.05)(表1)。

表1 Anti-CTLA-4 mAb對小鼠體內蟲荷及每g肝臟蟲卵數的影響Tab.1 The effect of anti-CTLA-4 mAb on the worm burden and eggs per gram liver in BALB/c mice
注:數值均為均數±標準差。N代表每組6只小鼠。*與感染對照組比較,P<0.05。
Note:All results were presented as mean ± S.D; n represents 6 mice in each group. *P<0.05, versus infected control group.
2.2 Anti-CTLA-4 mAb對脾淋巴細胞中CD4+CD25+Tregs百分比的影響 CTLA-4表達于CD4+CD25+Tregs,為了研究日本血吸蟲感染模型中CTLA-4在CD4+CD25+Tregs中的作用,流式細胞術檢測各組小鼠脾淋巴細胞中的CD4+CD25+Tregs百分比。既然Foxp3是CD4+CD25+Tregs的特異性標志,因此,用CD4+CD25+Foxp3+T 細胞表示CD4+CD25+Tregs。無菌取抗體處理組和對照組小鼠脾臟,常規制備脾淋巴細胞懸液,小鼠調節性T細胞染色試劑盒抗體標記。流式檢測結果見圖1,anti-CTLA-4 mAb組脾淋巴細胞中CD4+CD25+Tregs比例明顯高于對照組(P<0.05)。
2.3 脾細胞培養上清中細胞因子的含量 Anti-CTLA-4 mAb組脾淋巴細胞中細胞因子IFN-γ、IL-4、IL-5和IL-10的含量均明顯高于感染對照組(圖2)。
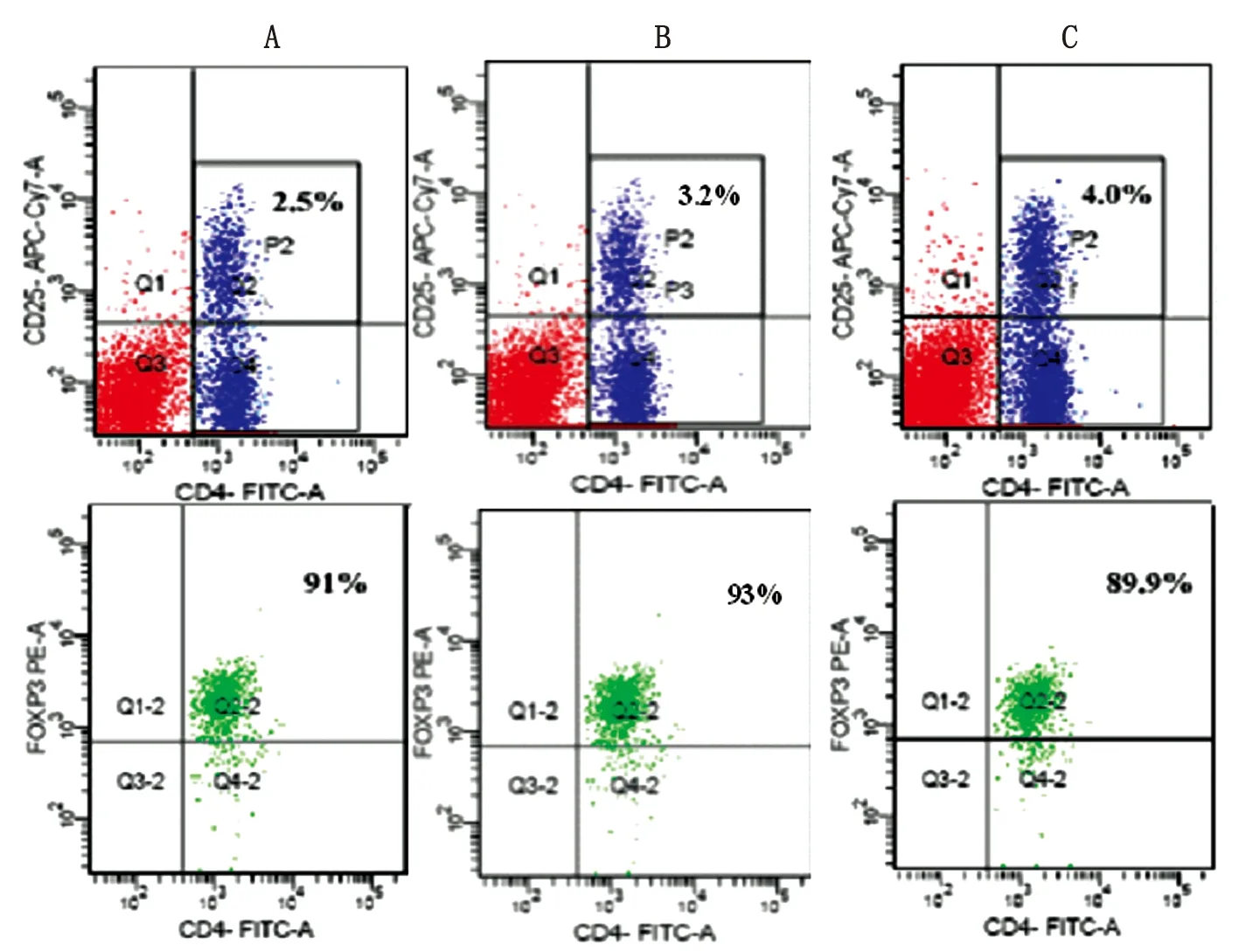
正常對照組(A), 感染對照組(B)和Anti-CTLA-4 mAb組 (C).P2門為CD4+CD25+T細胞在脾淋巴細胞中的百分比,Q2-2為P2門中的Foxp3+T細胞的百分比(即CD4+CD25+Foxp3+%)。
Normal control (A), infected control (B) and anti-CTLA-4 mAb (C). Upper panels, the cells in P2 gate denote the percentage of CD4+CD25+T cells in splenic lymphocytes. The cells in Q2-2 gate in the lower panels denote the percentage of Foxp3+lymphocytes in P2 gate.
圖1 流式細胞染色顯示CD4+CD25+Tregs在脾淋巴細胞中的的百分比。
Fig.1 FACS analysis demonstrates the percentage of CD4+CD25+Tregs in splenocytes
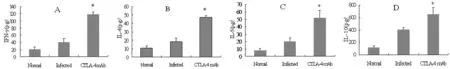
* 與感染對照組比較,P<0.05。* P<0.05, versus infected control group.圖2 BALB/c小鼠脾細胞培養上清中IFN-γ(A)、IL-4(B)、IL-5(C)和IL-10(D)的含量Fig.2 The expression of IFN-γ, IL-4, interleukin-5 and IL-10 in splenic lymphocytes culture supernatant
2.4 肝組織病理切片結果 肝組織HE染色結果見圖3,anti-CTLA-4 mAb組肝組織蟲卵肉芽腫直徑(503.2±53.6)較感染對照組(356.8±43.6)增大,組間差異有統計學意義。
3 討 論
CTLA-4為共刺激分子,與CD28分子結構具有76%的同源性。CTLA-4能競爭性抑制CD28分子與B7復合物結合,防止CD28分子促進T細胞激活[7]。CTLA-4最重要的功能是抑制T細胞激活,CTLA-4基因缺陷小鼠出現淋巴增殖及多器官淋巴細胞浸潤,出生后3~4周死亡[8]。Blank等[6]報道,采用抗CTLA-4單克隆抗體(anti-CTLA-4 mAb)能增強機體的免疫反應,有利于機體抗黑色素瘤免疫反應。CTLA-4在轉移性的黑色素瘤患者中,有利于腫瘤細胞的免疫逃避,使用相應抗體阻斷則有利于病情改善[9]。Martins等[10]報道,錐蟲感染模型中采用anti-CTLA-4 mAb有利于機體對病原體的清除,使寄生蟲血癥下降及利于宿主存活。本實驗證實,anti-CTLA-4 mAb使用時機體對日本血吸蟲的清除率為18.99%。CTLA-4可能有利于日本血吸蟲在宿主體內的免疫逃避。
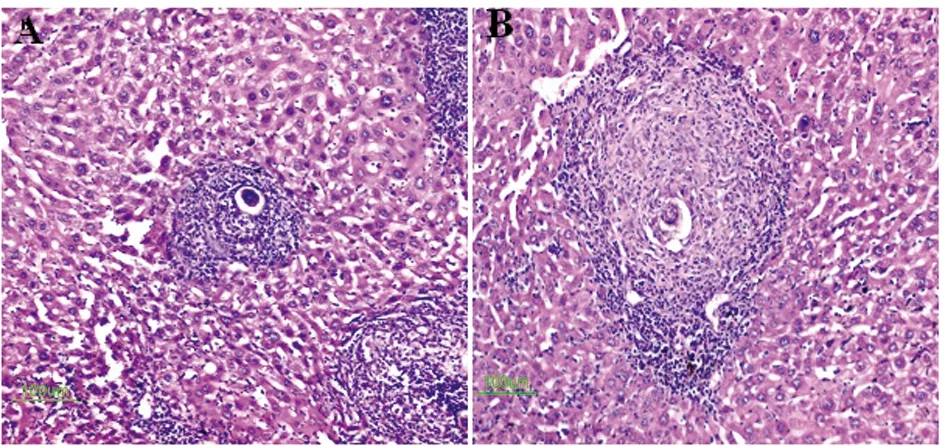
感染對照組(A)、anti-CTLA-4 mAb組(B)Infected control group (A), anti-CTLA-4 mAb group (B).圖3 各感染組蟲卵肉芽腫Fig.3 Schistosoma egg granuloma in infected groups
既然CTLA-4表達于CD4+CD25+Tregs,則很容易認為其免疫逃避作用是通過CD4+CD25+Tregs來實現的。本實驗使用流式方法檢測anti-CTLA-4 mAb對CD4+CD25+Tregs的影響,結果顯示:CD4+CD25+Tregs不降反升,表明CTLA-4在日本血吸蟲免疫逃避中的作用不是通過CD4+CD25+Tregs來實現的,Suarez N等[11]也報道,封閉CD4+CD25+Tregs與CTLA-4阻斷為不同的作用機制,兩者能起協同作用利于機體清除腫瘤。抗CTLA-4抗體作用的靶點在于增強效應T細胞的反應或者增強其對CD4+CD25+Tregs的抵抗。
CTLA-4為抑制性受體,能提高效應T細胞激活的閾值[12],用相應抗體阻斷后,則能使各種類型細胞增殖及各種類型細胞因子分泌增加。本實驗證實,anti-CTLA-4 mAb引起脾細胞培養上清中細胞因子IFN-γ明顯上升。Hoffman等[13]報道,Th1型免疫反應對機體清除日本血吸蟲很重要。因此,抗體使用后的殺蟲效應可能是通過增強Th1型免疫反應來實現的。
病理組織學檢測結果顯示,與感染對照組相比較,anti-CTLA-4 mAb及聯合組單個蟲卵肉芽腫直徑明顯增大。蟲卵肉芽腫主要是由日本血吸蟲卵引起的Th2型免疫反應[14],anti-CTLA-4 mAb導致Th2型細胞因子上升,因而引起蟲卵肉芽腫反應增強。Torino F等[15]報道,anti-CTLA-4 mAb具有明顯地抗腫瘤反應,同時加重對機體的病理損傷。Anti-CTLA-4 mAb增強機體免疫反應有利于機體清除寄生蟲的同時,以對機體的損傷增強為代價。
[1]Colley DG, Bustinduy AL, Secor WE, et al. Human schistosomiasis[J]. Lancet, 2014, 383(9936): 2253-2264.
[2]Higgins MK, Carrington M. Sequence variation and structural conservation allows development of novel function and immune evasion in parasite surface protein families[J]. Protein Sci, 2014, 23(4): 354-365.
[3]Vallejo A, Abad-Fernández M, Moreno S, et al. High levels of CD4+CTLA-4+Treg cells and CCR5 density in HIV-1-infected patients with visceral leishmaniasis[J]. Eur J Clin Microbiol Infect Dis, 2015, 34(2): 267-275.
[4]Haque A, Best SE, Amante FH, et al. CD4+natural regulatory T cells prevent experimental cerebral malaria via CTLA-4 when expandedinvivo[J]. PLoS Pathog, 2010, 6(12): e1001221.
[5]Tang CL, Lei JH, Wang T, et al. Effect of CD4+ CD25+ regulatory T cells on the immune evasion ofSchistosomajaponicum[J]. Parasitol Res. 2011, 108(2): 477-480.
[6]Mo HM, Liu WQ, Lei JH, et al.Schistosomajaponicumeggs modulate the activity of CD+4CD+25Tregs and prevent development of colitis in mice[J]. Exp Parasitol, 2007, 116(4): 385-389.
[7]Dilek N, Poirier N, Hulin P, et al. Targeting CD28, CTLA-4 and PD-L1 costimulation differentially controls immune synapses and function of human regulatory and conventional T-cells[J]. PLoS One, 2013, 8(12): e83139.
[8]Waterhouse P, Penninger JM, Timms E, et al. Lymphoproliferative disorders with early lethality in mice deficient in Ctla-4[J]. Science, 1995, 270(5238): 985-988.
[9]Matikas A, Mavroudis D. Beyond CTLA-4: novel immunotherapy strategies for metastatic melanoma[J]. Future Oncol, 2015, 11(6): 997-1009.
[10]Martins GA, Tadokoro CE, Silva RB, et al. CTLA-4 blockage increases resistance to infection with the intracellular protozoanTrypanosomacruzi[J]. J Immunol, 2004, 172(8): 4893-4901.
[11]Suarez N, Alfaro C, Dubrot J, et al. Synergistic effects of CTLA-4 blockade with tremelimumab and elimination of regulatory T lymphocytesinvitroandinvivo[J]. Int J Cancer, 2011, 129(2): 374-386.
[12]Egen JG, Kuhns MS, Allison JP. CTLA-4: new insights into its biological function and use in tumor immunotherapy[J]. Nat Immunol, 2002, 3(7): 611-618.
[13]Mendez S, Reckling SK, Piccirillo CA, et al. Role for CD4(+) CD25(+) regulatory T cells in reactivation of persistent leishmaniasis and control of concomitant immunity[J]. J Exp Med, 2004, 200(2): 201-210.
[14]Tian Z, Wang XY, Zhou YF, et al.SchistosomajaponicumscFv-IL18 fusion DNA ameliorates hepatic fibrosis in schistosomiasis-infected mice via improving local concentration of IL-18 in liver[J]. Exp Parasitol, 2013, 134(4): 447-454.
[15]Torino F, Barnabei A, De Vecchis L, et al. Hypophysitis induced by monoclonal antibodies to cytotoxic T lymphocyte antigen 4: challenges from a new cause of a rare disease[J]. Oncologist, 2012, 17(4): 525-535.
Effect and mechanism of CTLA-4 on the immune evasion ofSchistosomajaponicumin mice
TANG Chun-lian,SHEN Zhi-qin,YAN Lin-xia,LIU Xiao-hong
(DepartmentofClinicalLaboratory,WuchangHospital,Wuhan430063,China)
In order to explore the effect and mechanism of cytotoxic T-lymphocyte-associated protein 4 (CTLA-4) on the immune evasion ofSchistosomajaponicum(S.japonicum) in mice, the 18 female BALB/c mice were divided randomly into 3 groups: normal control, infected control and anti-CTLA-4 mAb-treated group. Each mouse in infected control and anti-CTLA-4 groups was challenged with 40 cercarie ofS.japonicum. Mice in anti-CTLA-4 mAb group were treated with 300 μg of anti-CTLA-4 mAb for 3 days consecutively, equal volume PBS as control. At 6 weeks post-infection, all mice were succumbed to measure worm burden and eggs per gram liver. The levels of IFN-γ, IL-4, IL-5 and IL-10 were detected by sandwich-ELISA. Flow cytometry was performed to measure the percentages of CD4+CD25+tregs in splenic cells. Liver sections were stained with hematoxylin and eosin to detect egg granuloma. The results showed that reduction rate of worm burden in anti-CTLA-4 mAb group were 18.99%. Compared with that of control group, the percentage of CD4+CD25+Tregs was extremely higher in anti-CTLA-4 mAb group. The egg granuloma size in anti-CTLA-4 mAb group were extremely larger than those of infected control group, while the levels of IFN-γ, IL-4, IL-5, IL-10 in anti-CTLA-4 mAb group were extremely higher than those of control group. It’s indicated that anti-CTLA-4 mAb favors the host to clearS.japonicumby enhancing Th1 type immune response, but at the cost of elevated pathological damage by enhancing Th2 type immune response. It hints that CTLA-4 contribute to the escape ofS.japonicumfrom the host immune responses.
Schistosomajaponicum; CTLA-4; immune evasion; cytokines
劉曉宏,Email: 494527907@qq.com
武漢市武昌醫院檢驗科,武漢 430063
10.3969/j.issn.1002-2694.2015.11.016
R383
A
1002-2694(2015)11-1061-04
2015-05-08;
2015-08-26
湖北省衛生和計劃生育委員會血防專項(No. WJ2015XB016)資助
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
海峽科技與產業(2016年3期)2016-05-17 04:32:12
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
