
外源葡萄糖對鹽脅迫下黃瓜幼苗光合作用和離子分布的影響
2015-02-28 01:37:24汪菊香
安徽農業科學 2015年1期
汪菊香
(安徽省南陵縣許鎮鎮農業綜合服務中心,安徽蕪湖 241000)
?
外源葡萄糖對鹽脅迫下黃瓜幼苗光合作用和離子分布的影響
汪菊香
(安徽省南陵縣許鎮鎮農業綜合服務中心,安徽蕪湖 241000)
[目的] 為了探討外源葡萄糖緩解鹽脅迫下黃瓜幼苗生理傷害的作用機制。[方法] 采用營養液水培法,以鹽敏感型黃瓜品種‘津春2號’為材料,研究外源葡萄糖對鹽脅迫下黃瓜幼苗生長、葉綠素含量、光合作用以及體內離子分布的影響。[結果] 鹽脅迫(65 mmol/L NaCl)顯著降低了黃瓜幼苗葉片的葉綠素含量、凈光合速率(Pn)、氣孔導度(Gs)和胞間CO2濃度(Ci),限制了黃瓜幼苗植株的生長,而根施2 mmol/L葡萄糖緩解了鹽脅迫誘導的光合速率和生長下降;在鹽脅迫下,黃瓜幼苗葉片、莖和根中Na+和Cl-含量顯著增加,而K+和Mg2+含量明顯降低,外源葡萄糖處理逆轉鹽脅迫誘導黃瓜植株體內離子含量的變化。[結論]外源葡萄糖通過抑制根中Na+和Cl-向地上部的運輸,減輕葉片中Na+和Cl-積累誘導的離子毒害,從而緩解鹽脅迫對光合作用的抑制,進而促進鹽脅迫下黃瓜幼苗的生長。
葡萄糖;黃瓜;鹽脅迫;生長;光合作用;離子分布
鹽脅迫對農業生產的威脅是一個世界性的熱點問題,也是影響全球生態環境的重要因素之一[1]。由于鹽漬土土壤具有相對較高的溶液濃度,易引起植物葉片滲透勢、水勢以及相對含水量下降,導致生長發育受到限制[2]。眾多研究證實,鹽脅迫抑制植株生長,通常與光合作用效率的下降密切相關。Stepien等[3]研究表明,鹽脅迫處理14 d的黃瓜植株凈光合速率(Pn)、氣孔導度(Gs)、蒸騰速率(Tr)、最大光化學效率(Fv/Fm)、光化學淬滅系數(qP)均顯著降低,而非光化學淬滅系數(qN)顯著升高。鹽脅迫下黃瓜葉片PSII實際光化學效率(ФPSII)、氣孔限制值(Ls)和胞間CO2濃度(Ci)下降[4]。研究表明,鹽脅迫導致光合效率下降主要是由非氣孔限制因素所致,如光合色素、葉綠體超微結構、光合電子傳遞、光合膜蛋白復合體以及光合磷酸化和CO2固定等方面受到抑制或損傷,且光合作用的抑制程度取決于鹽害的脅迫程度、持續時間以及不同耐性的植物品種[5]。
葡萄糖作為滲透調節物質,能提高細胞的滲透調節能力,具有維持細胞滲透壓平衡和增強植物響應非生物脅迫的能力。研究表明,水分脅迫下施用外源葡萄糖可通過促進小麥根系生長和提高葉組織的滲透調節能力,改善葉片的水分狀況和光合功能,從而減輕水分脅迫對小麥幼苗生長的抑制作用[6]。經進一步研究,發現在花后干旱和低氮互作脅迫下,外源噴施葡萄糖處理能夠明顯增加小麥單穗結實率和籽粒干物質的積累,延長旗葉的光合效率和灌漿持續期,促進小麥籽粒蛋白質和淀粉積累,進而提高小麥的產量[7]。在鹽脅迫下,施用外源葡萄糖能夠有效增強山楂、甜菜體內抗氧化酶的活性,降低活性氧過量積累誘導的氧化傷害[8]。此外,有研究表明,外源葡萄糖能夠提高鹽脅迫下水稻、玉米和紅小豆種子的發芽率、活力指數和發芽指數,增強種子對鹽脅迫環境的適應能力[9-11]。然而,對于葡萄糖如何增強鹽脅迫下蔬菜作物抗性的生理機制仍缺乏了解。
黃瓜(CucumissativusL.)具有產量高、營養豐富、效益好等特點,是世界性的重要蔬菜作物之一,也是我國設施栽培面積大、種植范圍廣的主要蔬菜作物之一。但是,由于黃瓜根系分布較淺,再生能力弱,對鹽漬環境敏感。一旦遭受鹽害,黃瓜植株生長發育就會受到顯著抑制,嚴重影響黃瓜的產量和品質。為此,筆者采用營養液水培法,通過根施外源葡萄糖處理鹽脅迫下黃瓜幼苗,研究外源葡萄糖對鹽脅迫下黃瓜幼苗生長、光合作用和體內離子分布的影響,以期探討外源葡萄糖緩解鹽脅迫下黃瓜幼苗生理傷害的作用機制,為利用外源葡萄糖增強黃瓜植株耐鹽性提供理論依據。
1 材料與方法
1.1 幼苗培養試驗于2013年3~6月在塑料大棚內進行。選用鹽敏感型的黃瓜品種‘津春2號’(購自天津市科潤黃瓜研究所)為試驗材料,種子經消毒、浸種后,在29 ℃下催芽24 h,播于裝有基質的塑料盤中育苗。棚內晝溫控制在20~25 ℃,夜溫晝溫控制在15~18 ℃,光合有效量子密度(PPFD)最大為900 μmol/(m2·s)左右,相對濕度維持在55%~75%。在兩片子葉展開后,澆1/2劑量的日本園試配方營養液。待第2片真葉展開后,選取生長整齊一致的幼苗定植于裝有1個劑量園試配方營養液的箱(60 L)中,營養液pH調至6.5±0.1,電導率(EC)2.0~2.2 mS/cm,氣泵通氣20 min/h 。
1.2 試驗處理預培養2 d后,向營養液中添加NaCl。試驗設4個處理:①對照(CK),正常營養液栽培;②對照+葡萄糖(CK+Glu);③鹽脅迫(NaCl);④鹽脅迫+葡萄糖(NaCl +Glu)。當開始NaCl處理時,處理②、④外源葡萄糖直接添加到營養液中。每個處理72株,試驗隨機排列,3次重復。處理7 d后,測定各項指標,并且選擇第 3 片真葉(從基部數)測定光合作用參數和葉綠素含量。該試驗所采用的NaCl脅迫濃度(65 mmol/L)和根施葡萄糖處理濃度(2 mmol/L)均由預備試驗篩選得出。
1.3 指標測定與方法
1.3.1生長指標的測定。株高(莖基部到生長點)用直尺測量;莖粗(莖基部)用游標卡尺測量;鮮樣用去離子水沖洗干凈并吸干水分后稱得鮮重;在烘箱中105 ℃下殺青15 min后,降溫到75 ℃烘干至恒重,稱得干重。
1.3.2光合色素含量的測定。葉綠素含量根據Holden[12]的方法,用乙醇∶丙酮∶水4.5∶4.5∶1.0萃取完整葉片色素含量,利用紫外分光光度計測量。
1.3.3氣體交換參數的測定。用便攜式光合作用測定系統(Li-6400,美國),于上午9:30~11:30測定各處理幼苗第三葉位葉片的光合氣體交換參數。每個處理測定3株。測定時,使用開放氣路,葉室溫度控制在(28±1) ℃,光強控制在800 μmol /(m2· s),參比室CO2濃度為(370±10)μmol/mol,相對濕度為60%~70%。測定指標主要包括葉片的Pn、Tr、Gs和Ci。
1.3.4離子含量分析。提取方法參照王寶山等[13]方法,略加改動。稱取烘干樣品40 mg于試管中,加20 ml去離子水,搖勻,沸水浴2 h,冷卻后過濾于50 ml容量瓶中定容。K+、Na+和Mg2+含量使用日立Z-2000型原子吸收分光光度計進行測定。Cl-含量采用AgNO3滴定法進行測定,以0.22 mol/L K2CrO4和0.03 mol/L K2Cr2O7為混合指示劑。
1.4 數據分析試驗數據均采用SAS13.1統計分析軟件進行分析。采用Duncan多重比較法進行統計分析,P<0.05。
2 結果與分析
2.1 葡萄糖對鹽脅迫下黃瓜幼苗生長的影響由表1可知,在正常營養液栽培條件下,根施外源葡萄糖對黃瓜幼苗生長指標沒有明顯的影響。65 mmol/L NaCl脅迫處理7 d后,黃瓜幼苗株高、莖粗、鮮重和干重明顯下降,與對照相比分別下降37.04%、17.48%、40.85%和37.55%。鹽脅迫下,外源葡萄糖顯著增加黃瓜幼苗株高、莖粗、鮮重和干重,分別比單純鹽脅迫處理提高47.78%、19.22%、51.39%和40.17%,說明外源葡萄糖緩解鹽脅迫誘導黃瓜幼苗生長的抑制。
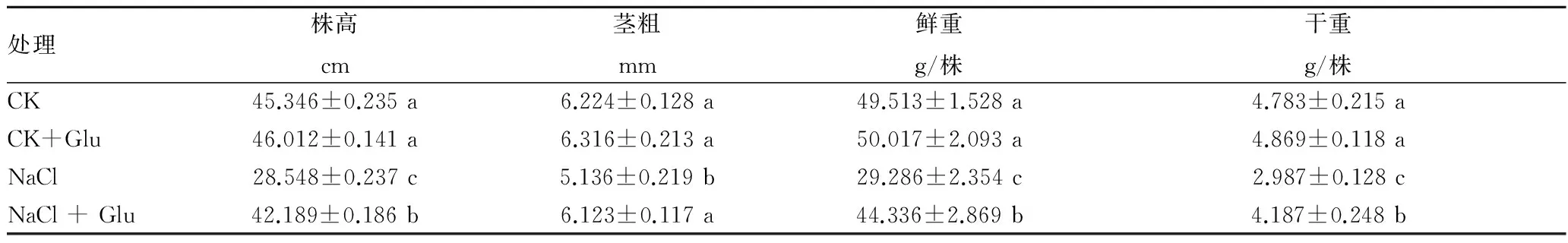
表1 外源葡糖糖對鹽脅迫下黃瓜幼苗生長的影響
注:每個值都是由3個重復的平均值±標準誤差。不同字母表示差異達0.05顯著水平。
2.2 葡萄糖對鹽脅迫下黃瓜幼苗光合色素含量的影響由表2可知,鹽脅迫下,黃瓜幼苗葉片葉綠素a(Chl a)、Chl(a+b)含量和Chl a/Chl b顯著下降,與對照相比分別降低了39.98%、26.16%和49.46%,而葉綠素b(Chl b)含量顯著增加。根施葡萄糖處理顯著提高鹽脅迫下Chl a、Chl(a+b)和Chl a/Chl b的值,與鹽脅迫相比分別增加了51.31%、23.62%和93.48%,而Chl b含量顯著下降。在正常營養液栽培條件下,根施葡萄糖對黃瓜幼苗光合色素含量沒有明顯的影響。

表2 外源葡萄糖對鹽脅迫下黃瓜幼苗光合色素含量的影響
注:每個值都是由3個重復的平均值±標準誤差。不同字母表示差異達0.05顯著水平。
2.3 葡萄糖對鹽脅迫下黃瓜幼苗光合作用的影響由圖1可知,鹽脅迫下,黃瓜幼苗葉片Pn、Gs和Tr顯著下降,與正常營養液栽培相比分別下降了62.44%、57.44%和65.68%,而鹽脅迫顯著增加Ci,比對照增加了19.70%。外源葡萄糖處理顯著提高了黃瓜葉片Pn、Gs和Tr,與單純鹽脅迫處理相比分別增加了100.15%、61.10%和145.58%,并且降低了Ci。在正常營養液栽培條件下,外源葡萄糖處理對黃瓜幼苗葉片光合作用無明顯影響。
2.4 葡萄糖對鹽脅迫下黃瓜植株體內離子分布的影響由表3可知,鹽脅迫下,黃瓜不同器官中Na+和Cl-含量明顯增加,而Cl-增加的幅度高于Na+;與正常營養液栽培相比,鹽脅迫下黃瓜幼苗植株葉片、莖中和根中Na+含量分別增加了585.12%、508.87%和258.04%,而葉片、莖中和根中Cl-含量分別增加了2 043.62%、2 419.39%和1 239.47%;鹽脅迫下,黃瓜不同器官中K+和Mg2+含量均明顯下降,而K+下降的幅度高于Mg2+;與對照相比,葉片、莖中、根中K+和Mg2+含量分別下降了62.02%、81.77%、72.93%和44.57%、59.22%、74.07%。外源葡萄糖處理明顯逆轉了鹽脅迫下黃瓜植株體內離子含量的變化。葡萄糖處理明顯降低鹽脅迫下黃瓜植株中Na+和Cl-含量,與單純鹽脅迫相比黃瓜葉片、莖、根中Na+、Cl-含量分別降低了58.26%、35.53%、52.70%和41.89%、35.76%、33.99%;同時,葡萄糖明顯增加鹽脅迫下黃瓜葉片、莖、根中K+和Mg2+含量,與鹽脅迫相比分別增加了111.64%、216.72%、97.10%和54.90%、61.91%、92.86%。在正常營養液栽培條件下,施用外源葡萄糖對黃瓜幼苗不同組織中的離子含量無明顯的影響。

表3 外源葡萄糖對鹽脅迫下黃瓜幼苗根、莖和葉中離子含量的影響
注:每個值都是由3個重復的平均值±標準誤差。不同字母表示差異達0.05顯著水平。
3 討論
研究表明,65 mmol/L NaCl脅迫處理7 d后,黃瓜幼苗生長受到顯著抑制,而施用2 mmol/L葡萄糖可明顯提高鹽脅迫下黃瓜幼苗的株高、干重和鮮重。這可能是由于葡萄糖改善了鹽脅迫下黃瓜幼苗的光合作用。
光合作用是地球上所有生物體獲取能量和物質的基礎。提高光合作用的效率是提高農作物產量的又一重要途徑。研究表明,鹽脅迫導致植物光合效率下降主要包括氣孔因素和非氣孔因素2個方面。判斷的依據主要是看Pn、Ci和Gs三者的變化趨勢[14]。如果Pn下降,那么伴隨著Ci和Gs的同時下降,基本上可以確定凈光合速率的下降主要是由氣孔因素引起的;反之,如果Gs下降,而Ci維持不變甚至上升,則Pn的下降是由葉肉細胞同化能力下降或光合器官組分等非氣孔因素所致。該試驗表明,在鹽脅迫下,黃瓜幼苗葉片Pn、Ci和Gs顯著降低,而Ci顯著升高,表明鹽脅迫引起光合速率的下降主要是由非氣孔因素所致。施用外源葡萄糖處理可顯著緩解鹽脅迫誘導黃瓜幼苗葉片Pn、Ci和Gs的下降,并降低Ci的濃度。這與于翠等[15]在葡萄糖緩解鹽脅迫下山楂光合特性上的研究結果相一致,表明葡萄糖提高鹽脅迫下黃瓜幼苗葉片光合速率主要是通過作用于非氣孔因素。
光合色素是一類含脂的色素,存在于葉綠體內基粒類囊體上,主要包含葉綠素、反應中心色素和輔助色素,在光合作用中參與光能吸收、傳遞或原初光化學反應等過程。高等植物和大部分藻類的色素主要是由葉綠素 a、葉綠素 b和類胡蘿卜素(包括胡蘿卜素和葉黃素)組成[1]。研究表明,鹽脅迫對高等植物光合色素含量的影響依賴于試驗材料、鹽處理濃度及處理時間。王加真等[16]通過研究不同鹽脅迫程度下溝葉結縷草Chl a和 Chl b含量的變化,發現在高濃度鹽脅迫條件(1.5、2.0 mmol/L NaCl)下所有光合色素含量都減少,中等脅迫濃度(1 mmol/L NaCl)下光合色素含量先增加后減少,而在低濃度條件(0.5 mmol/L NaCl)下光合色素含量有增加的趨勢。試驗中,65 mmol/L NaCl處理顯著降低黃瓜幼苗葉片Chl a、Chl a+b和Chl a/b的值,暗示鹽脅迫可能誘導葉綠素降解酶活性的增加,從而促進葉綠素降解[4]。外源施用葡萄糖處理顯著緩解鹽脅迫下黃瓜幼苗Chl a的降低,并且提高葉綠素a/b的比值,表明葡萄糖可能抑制葉綠素降解酶的活性,保持其結構的穩定性,并且使黃瓜葉片具有較高的葉綠素含量,有助于黃瓜葉片對光能的吸收,提高光能轉化效率,從而保證鹽脅迫下黃瓜幼苗光合作用的正常進行[1]。
植物光合作用在鹽脅迫下的響應與植物自身阻止鹽分離子向體內運輸以及維持細胞內的離子平衡密切相關[17]。當細胞外Na+和Cl-含量累積到一定臨界值時,細胞內的水分子快速地從水通道離開細胞,造成細胞失水。與此同時,Na+、Cl-可分別通過Na+/K+、Cl-通道進入細胞質,然后進入葉綠體體內,雖然離子在細胞中運動的具體機制目前仍不清楚,但是光合器官中較高濃度的Na+和Cl-含量將抑制光合酶活性[18]。另外,植株體內Na+和Cl-的過量積累將會抑制K+和Mg2+的吸收,而K+和Mg2+又是植物光合作用中多種酶的活化劑,其含量的減少勢必會進一步加劇光合作用的抑制。Li等[19]研究表明,隨著鹽脅迫處理濃度的增加,紅樹林植株葉片中積累的Na+和Cl-含量也相應增加,使得植物細胞體內養分吸收動態平衡發生紊亂,導致光合作用受到抑制。該試驗結果表明,施用外源葡萄糖處理可有效降低黃瓜植株體內Na+和Cl-含量,并且抑制Na+和Cl-由植株體內向地上部的運輸,提高植株體內K+和Mg2+含量,說明葡萄糖作為一種滲透調節物質和能量物質,可直接或間接地維持細胞的離子選擇性,通過調節過量鹽分離子進入植株體內,維持黃瓜幼苗體內養分離子的吸收平衡,緩解鹽分離子對光合機構的毒害,進而提高鹽脅迫下黃瓜幼苗光合效率。而對于葡萄糖如何調節鹽分離子在植株體內的運輸和轉運還需進一步的研究。
[1] 束勝,郭世榮,孫錦,等.鹽脅迫下植物光合作用的研究進展[J].中國蔬菜,2012(18):53-61.
[2]SUDHIR P R,MURTHY S D S.Effects of salt stress on basic processes of photosynthesis[J].Photosynthetica,2004,42(4):481-486.
[3]STEPIEN P,KLOBUS G.Water relations and photosynthesis inCucumissativusL.Ieaves under salt stress[J].Biologia Plantarum,2006,50(4):610-616.
[4]ZHANG R H,LI J,GUO S R,et al.Effects of exogenous putrescine on gas-exchange characteristics and chlorophyll fluorescence of NaCl-stressed cucumber seedlings[J].Photosynthesis Research,2009,100(3):155-162 .
[5]MEGDICHE W,HESSINI K,GHARBI F,et al.Photosynthesis and photosystem II efficiency of two salt-adapted halophytic seashoreCakilemaritimaecotypes[J].Photosynthetica,2008,46(3):410-419.
[6] 胡夢蕓,李輝,李倩影,等.花后干旱和低氮脅迫下外源葡萄糖對小麥產量性狀的影響[J].中國農學通報, 2012,28(27):118-123.
[7] 胡夢蕓,李輝,張穎君,等.水分脅迫下葡萄糖對小麥幼苗光合作用和相關生理特性的影響[J].作物學報,2009,35(4):724-732.
[8] 曹光峰,郭洪海,田高中,等.施加外源葡萄糖對甜菜響應鹽脅迫的影響[J].安徽農業科學,2012,40(8):4494-4495.
[9] 周紅菊.水楊酸和葡萄糖對鹽脅迫下玉米種子萌發的緩解作用[J].湖北農業科學,2012,51(8):1539-1540.
[10] 尹相博,于立芝,湯麗娟.Ca2+和葡萄糖對鹽脅迫下紅小豆種子萌發的影響[J].廣東農業科學,2013(21):39-42.
[11] 凌騰芳,宣偉,樊穎瑞,等.外源葡萄糖、果糖和NO供體(SNP)對鹽脅迫下水稻種子萌發的影響[J].植物生理與分子生物學學報,2005,31(2):205-212.
[12]HOLDEN M.Chlorophylls[M]//GOODWIN T W.Chemistry and Biochemistry of Plant Pigments.London.London,Academic Press,1965:461-488.
[13] 王寶山,趙可夫.小麥葉片中Na、K提取方法的比較[J].植物生理學通訊,1995,31(1) :50-52.
[14] FARQUHAR G D,SHARKEY T D.Stomatal conductance and photosynthesis[J].Annual Review of Plant Physiology,1982,33:317-345.
[15] 于翠,呂德國,楊磊,等.葡萄糖對鹽脅迫下山楂幼苗光合熒光特性的影響[J].沈陽農業大學學報,2009,40(4):408-412.
[16] 王加真,夏更壽,李建龍,等.高鹽脅迫對溝葉結縷草葉片光合色素含量的影響[J].上海交通大學學報,2007,25(6):583-586.
[17]SIMON P,ROBINSON W,JOHN S D.Photosynthesis and ion content of leaves and isolated chloroplasts of salt-stressed spinach[J].Plant Physiol,1983,73:238-242.
[18]WANG R,CHEN S,DENG L,et al.Leaf photosynthesis,fluorescence response to salinity and the relevance to chloroplast salt compartmentation and anti-oxidative stress in two poplars[J].Trees,Struct and Funct,2007,21(5):581-591.
[19]LI N Y,CHEN S L,ZHOU X Y,et al.Effect of NaCl on photosynthesis,salt accumulation and ion compartmentation in two mangrove species,Kandelia candeln and Bruguiera gymnorhiza[J].Aquatic Botany,2008,88:303-310.
Effects of Exogenous Glucose on Photosynthesis and Ion Distribution of Cucumber Seedlings under Salt Stress
WANG Ju-xiang
(Xuzhen Comprehensive Agricultural Service Center of Nanling County,Wuhu,Anhui 241000)
[Objective] The research aimed to investigate the mechanism of the relief of exogenous glucose on cucumber seedlings exposed to salt stress.[Method] Taken sensitivity pattern cucumber ‘Jinchun 2’as material,the effects of exogenous glucose on plant growth,chlorophyll content,gas exchange parameter and ion distribution of cucumber seedlings exposed to salt stress were studied by hydroponics.[Result] 65 mmol/L NaCl salt stress induced a severe reduction of growth,chlorophylls content and photosynthesis.Application of exogenous glucose (2 mmol/L glucose) alleviated the reduction of growth and net photosynthetic rate (Pn) in salt stressed-cucumber plants.Salt stress resulted in a significant accumulation of Na+and Cl-in roots,stems and leaves,but decreased the levels of K+and Mg2+.However,application of glucose to salt-stressed plants counteracted the adverse effects of excess ion accumulation.[Conclusion] Exogenous glucose inhibited the transportation of Na+and Cl-from roots to leaves,decreased the accumulation of Na+and Cl-in different tissues under salt stress,which was associated with an improvement in photosynthetic efficiency.
Glucose; Cucumber; Salt stress; Growth; Photosynthesis; Ion distribution
汪菊香(1975-),女,安徽蕪湖人,農藝師,從事瓜果蔬菜技術方面的研究。
2014-11-21
S 642.2
A
0517-6611(2015)01-007-04
