基于矢耳石形態的平鲉屬(Sebastes)魚類鑒別及研究方法比較
2015-06-24 14:10:05張鳳俠張秀梅覃樂政李文濤
海洋學報 2015年4期
關鍵詞:差異
張鳳俠, 張秀梅*, 覃樂政, 李文濤
(1.中國海洋大學 教育部海水養殖重點實驗室,山東 青島 266003)
基于矢耳石形態的平鲉屬(Sebastes)魚類鑒別及研究方法比較
張鳳俠1, 張秀梅1*, 覃樂政1, 李文濤1
(1.中國海洋大學 教育部海水養殖重點實驗室,山東 青島 266003)
分析了青島膠州灣鎧平鲉(Sebasteshubbsi)、花斑平鲉(Sebastesnigricans)和許氏平鲉(Sebastesschlegelii) 3種平鲉屬魚類的矢耳石形態特征,探究基于矢耳石形態鑒別3種魚類的可行性。以8個耳石形狀指標和37個橢圓傅里葉系數進行的單因素方差分析結果基本一致,顯示鎧平鲉與許氏平鲉的耳石形態差異最大,鎧平鲉與花斑平鲉間差異最小。利用耳石形狀指標、橢圓傅里葉系數、耳石形狀指標與橢圓傅里葉系數相結合的方法,對3種魚類的綜合判別率均可達95%以上,判別分析散點圖可明顯將3種魚類區分開。耳石長介于4~7 mm的3種魚類小規格組與2齡及以上成魚組耳石樣本的判別結果一致,表明針對不同發育階段平鲉屬魚類,基于耳石形態的種類鑒別均是可行的;僅使用橢圓傅里葉系數鑒別3種魚類時綜合判別率雖高達100%,但難以快速定位3種魚類耳石形態間,在統計上差異顯著而實際上微小的形狀差異所在;而使用耳石形狀指標和橢圓傅里葉系數聯合判別,綜合判別率亦高達99%,且能快速找出3種魚類耳石在矩形趨近率、形態因子、面密度等的形態差異。研究表明,耳石形狀指標和橢圓傅里葉分析聯合判別是平鲉屬魚類種類鑒別及耳石形態差異分析的有效手段。
平鲉屬;耳石形態;耳石形狀指標;橢圓傅里葉系數;鑒別
1 引言
耳石是魚類生長過程中沉淀在內耳中的鈣質結構,分矢耳石(sagittae)、微耳石(lapillus)和星耳石(asteriscus)共3對。耳石隨著魚類的生長而生長[1],一般認為達到性成熟后耳石形態趨于穩定[2-3]。由于耳石具有不易消化的特點,被廣泛應用于食魚性動物的食性分析中[4]。研究表明,耳石形態主要受遺傳[5]、環境[6]、攝食[7]等因素影響,其中遺傳占主導因素[8-9]。Morales-Nin[10]認為魚類耳石在不同空間方向上的沉積導致耳石形狀不規則;Maisey[11]、葉振江[12]指出耳石形態具有高度的物種特異性和顯著的種間差異性;Begg等[13]指出采用先進的數碼圖像分析耳石形狀為種群識別提供可能。眾多研究表明,耳石二維圖像的數學分析可以定量解釋魚類種間、群體間的耳石形態差異,可用于魚類種間、群體間的識別研究。
依據耳石形態,在鮨屬(Serranus)[14]、鯉科(Cyprinidae)[15]、鱭屬(Coilia)[16]、鱒屬(Salmo)[17]、四大家魚仔稚魚[18]以及熱帶鳀科(Engraulididae)[19]等魚類的種類鑒別中取得了較好的鑒別效果。但在耳石形態分析時,參數選擇和數據分析方法不同,會顯著影響鑒別結果。目前,常用的耳石分析方法包括文字描述法[2]、可量性狀比較法[16]、地標法[20]、耳石形狀指標法[9]與橢圓傅里葉分析法[21]。耳石形狀指標法運用于九帶鮨(Serranuscabrilla)[22]、鱚屬(Sillago)[20]等的鑒別研究中;耳石橢圓傅里葉分析已成功進行竹筴魚(Trachurustrachurus)[23]和條紋婢(Latrislineata)[24]的群體鑒別;耳石形狀指標法結合橢圓傅里葉分析,已在黑線鱈(Melanogrammusaeglefinus)[25]和大西洋鱈(Gadusmorhua)[26]的鑒別中應用。
平鲉屬(Sebastes)隸屬于鲉形目(Scorpaeniformes)鲉亞目(Scorpaenoidei)的鲉科(Scorpaenidae),我國常見分布種為鎧平鲉(Sebasteshubbsi)、花斑平鲉(Sebastesnigricans)和許氏平鲉(Sebastesschlegelii),主要分布在黃海、渤海海域,為巖礁區底層經濟魚類。目前3種魚類的鑒別主要依據背鰭鰭棘數、鰭條數和體側斑點等生物學特征。依據外部形態特征的種類鑒別,當樣品保存不當時,種類鑒別則受到限制,此時形態穩定性較好的耳石則可作為分類的輔助手段。截至目前,基于耳石形態的平鲉屬魚類的種類鑒別尚無報道,本研究以青島膠州灣的平鲉屬魚類為研究對象,旨在探究基于耳石形態鑒別平鲉屬魚類的可行性,為傳統形態分類提供佐證和輔助資料;并比較分析不同方法對鑒別結果的影響,嘗試找出適合平鲉屬魚類耳石分類的研究方法,為今后開展相關耳石研究提供基礎資料。
2 材料和方法
2.1 實驗材料
實驗所用平鲉屬樣本于2013年5-8月采自青島膠州灣海域地籠網和拖網漁獲物。種類鑒定后測量體長(精確到1 mm)、體質量(精確到0.01 g)(表1)。取出左、右矢耳石(以下皆以耳石簡稱),洗凈后保存于離心管中待進一步分析。

表1 膠州灣3種平鲉屬魚類樣品的體長與體質量
2.2 研究方法
2.2.1 年齡鑒定及耳石樣本的選取
將洗凈的耳石浸入盛水的培養皿中,凹面向上,置于Nikon SMZ8000型體式顯微鏡下,透射光觀察,可見清晰的輪紋,其中一個完整的明帶和暗帶計為一個年輪。左、右耳石的年齡鑒定結果一致,實驗首先選取2齡及以上平鲉屬魚類耳石用于后續分析。
2.2.2 耳石形態參數的選取
將選取的耳石置于50℃烘箱中干燥12 h以上至恒重。干燥后的耳石用精密電子天平稱重,單位精確到0.01 mg。耳石凸面向上、凹面向下,用連接于體式顯微鏡的Nikon DN100數碼相機拍攝耳石的二維圖像(見圖1)。
采用Image-ProPlus6.0圖像分析軟件分析耳石圖像,測量耳石形態學參數,共得出8個尺寸參數:耳石面積、耳石周長、耳石長、耳石寬、最大Feret直徑、最小Feret直徑、最大半徑和最小半徑。其中半徑測量的是通過耳石幾何中心的半徑長,Feret直徑指耳石按照某一角度擺放時,經過形心的耳石外緣兩點間的距離,面積為耳石的二維投影面積。根據潘曉哲和高天翔[20]、徐勝勇等[27]、Tuset等[28]、Canas等[29]相關研究,結合平鲉屬魚類的耳石形態特征,將8個耳石尺寸參數和耳石質量通過計算公式的轉換,得出平鲉屬魚類的8個耳石形狀指標(Otolith shape indices)(見表2)。耳石形狀指標在一定程度上可消除因耳石放置位置不同、耳石大小不同等因素對圖像分析結果的影響。
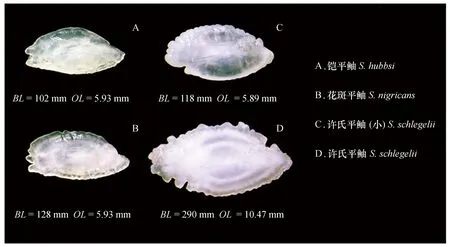
圖1 膠州灣3種平鲉屬魚類左耳石內側面照片(BL代表體長,OL代表耳石長)Fig.1 Left sagittal otoliths of three Sebastes collected in the Jiaozhou Bay shown with the proximal surface up (BL represents body length, OL represents otolith length)
表2 平鲉屬魚類耳石尺寸參數和形狀指標
Tab.2 Size parameters and size based shape indices with calculation formulas forSebastesspecies

尺寸參數耳石形狀指標面積(A)圓度=(4A)/(πOL2)周長(P)矩形趨近率=A/(OL×OW)耳石長(OL)橢圓率=(OL-OW)/(OL+OW)耳石寬(OW)形態因子=(4πA)/P2耳石質量(OW′)幅形比=OL/OW最大Feret直徑(Fmax)Feret直徑比=Fmax/Fmin最小Feret直徑(Fmin)半徑比=Rmax/Rmin最大半徑(Rmax)面密度=OW′/A最小半徑(Rmin)
注:文中以OW′代表耳石質量。面積度量單位為mm2,長度度量單位為mm,質量度量單位為mg。
橢圓傅里葉分析(Elliptic Fourier analysis)能夠有效對耳石輪廓進行描述,定量獲取耳石外輪廓的有效信息[30]。研究表明,傅里葉諧值越多越能精確地描述物體的外輪廓[31],但前10~20個傅里葉特征系數能夠描述耳石的主要形態輪廓[32]。本研究使用SHAPE1.3軟件包前10組傅里葉諧波對耳石圖像進行分析。每組諧波由4個形態變量an、bn、cn、dn組成,因此,每個耳石共由40個傅里葉特征系數描述。對10個諧值進行標準化后,前3個特征系數為定值:a1=1,b1=c1=0,故有效傅里葉特征系數為37個。
2.2.3 數據處理
本研究分別對3種平鲉屬魚類左、右耳石各原始測量指標進行獨立樣本T檢驗,以檢驗3種魚類種內左、右耳石間的差異性。同樣使用獨立樣本T檢驗,檢測3種魚類雌、雄個體間耳石形態的差異情況。顯著性水平取α=0.05。
鑒于魚類個體生長差異可能對檢驗結果產生影響,為實現無偏分析,在樣本選擇時除選取2齡及以上的性成熟個體外,還在數據分析時引入協變量(體長)以降低或消除個體生長差異對耳石形態參數的影響。本文采用Pearson相關性分析確定耳石形態參數(8個耳石形狀指標和37個橢圓傅里葉系數)與體長間關系。當體長與耳石形態參數存在線性相關時,參照竇碩增[33]的協方差校正方法,利用回歸斜率按照體長校正公式校正各耳石形態參數:
(1)
以單因素方差分析檢驗各個耳石形態參數在3種魚類間的差異性,用多重比較驗證差異來源。以上統計分析過程中,對具有方差齊性的變量采用LSD法,不具方差齊性的采用Tamhane’s T2法,取顯著性水平α=0.05。
利用SPSS18.0對3種平鲉屬魚類作判別分析,本文采用3種方式進行對比分析:(1)僅使用8個耳石形狀指標;(2)僅使用37個橢圓傅里葉特征系數;(3)8個耳石形狀指標并結合37個橢圓傅里葉特征系數。
2.2.4 對比分析樣本的補充
2齡及以上3種平鲉屬魚類間體長、耳石長存在較大差異,而近岸捕撈生產中常見低于1齡的許氏平鲉幼魚。因此,為驗證鑒別方法的有效性,本實驗對比加入不足1齡的許氏平鲉幼魚樣本,即補充加入耳石長介于4~7 mm的小個體許氏平鲉樣本。將耳石長介于4~7 mm的3種平鲉屬魚類單獨進行判別分析,以充分探討耳石形態在3種魚類種類鑒別中的可行性。在此,采用耳石長作為協變量引入協方差校正方程,校正與耳石長顯著相關的參數,校正后的參數用于后續判別分析,進而比較交叉檢驗結果。
為有效區分,下文中2齡及以上許氏平鲉樣本統一稱為許氏平鲉;耳石長介于4~7 mm的小個體許氏平鲉統一稱為許氏平鲉(小)。下文中,若未詳細說明,則3種平鲉屬魚類均指2齡及以上、耳石形態趨于穩定的樣本;耳石長介于4~7 mm的3種魚類樣本信息為:鎧平鲉(n=61,體長范圍73~119 mm,耳石長范圍4.45~6.58 mm)、花斑平鲉(n=14,體長范圍102~165 mm,耳石長范圍4.97~6.64 mm)、許氏平鲉(小)(n=45,體長范圍101~131 mm,耳石長范圍4.29~5.93 mm)。
3 結果
3.1 種內耳石形態的差異性
對左、右耳石的8個測量指標和耳石質量分別進行T檢驗,結果顯示3種平鲉屬魚類的左、右耳石在這些測量指標上均無顯著性差異。
對雌、雄個體耳石的8個測量指標和耳石質量分別進行T檢驗,結果顯示3種平鲉屬魚類雌、雄個體的耳石在這些測量指標上均無顯著性差異。
因此,下文中的耳石形態研究不分性別統一使用左耳石。表3為以鎧平鲉為例,左、右耳石以及雌、雄個體間耳石差異性的檢驗結果。
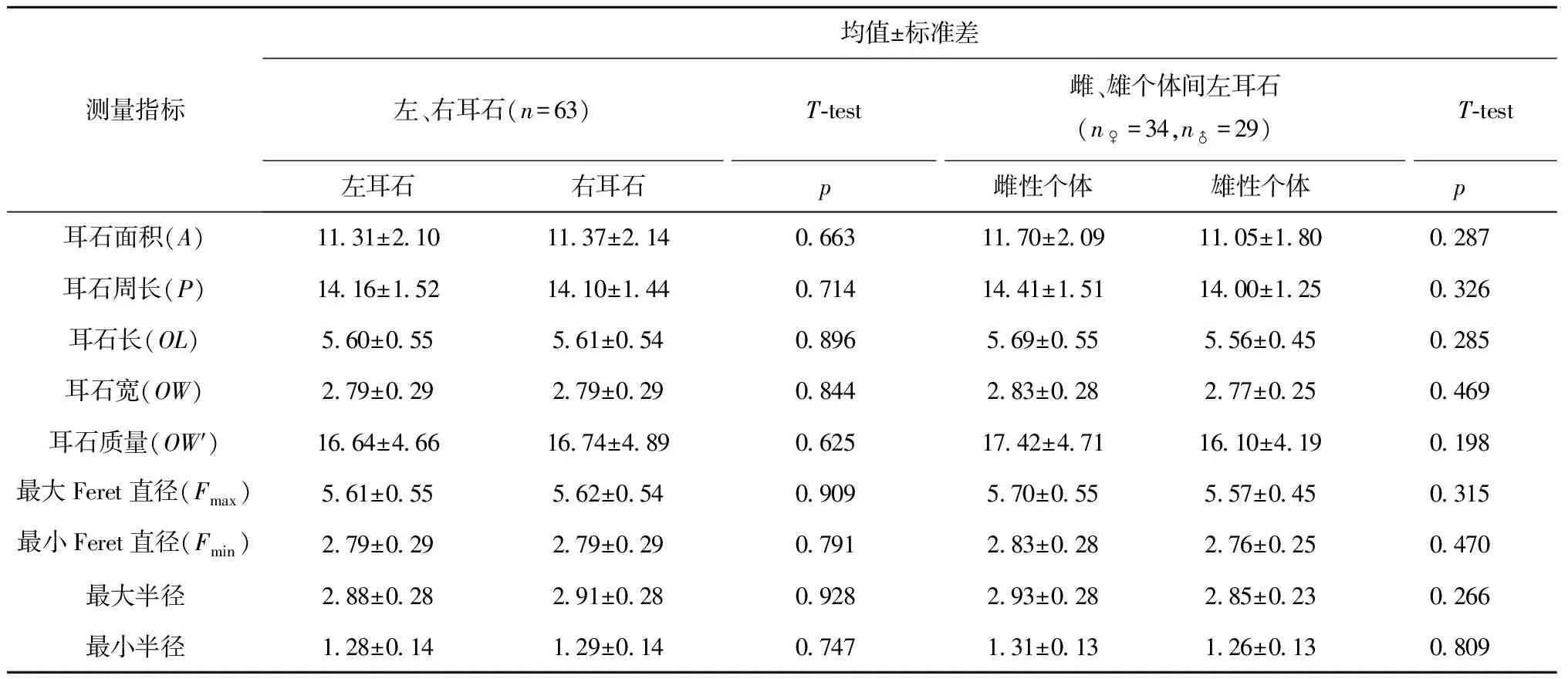
表3 鎧平鲉左右耳石、雌雄個體間耳石差異性比較
注:n♀表示鎧平鲉雌性個體的數量,n♂表示雄性個體的數量。
3.2 單因素方差分析
Pearson相關分析得出3個耳石形狀指標(橢圓率、形態因子、面密度)及14個橢圓傅里葉系數(h4、h6、h11、h12、h14、h17、h18、h19、h22、h23、h25、h29、h30、h37)與體長顯著相關,這些變量使用協方差校正方法校正后進行后續的數據分析。
8個耳石形狀指標及單因素方差分析結果如表4所示。鎧平鲉和許氏平鲉,花斑平鲉和許氏平鲉間均有7個耳石形狀指標存在顯著差異;鎧平鲉和花斑平鲉有3個指標存在顯著差異。圓度在3種平鲉屬魚類間差異均不顯著,3種魚類兩兩之間在矩形趨近率、形態因子、面密度間均差異顯著。
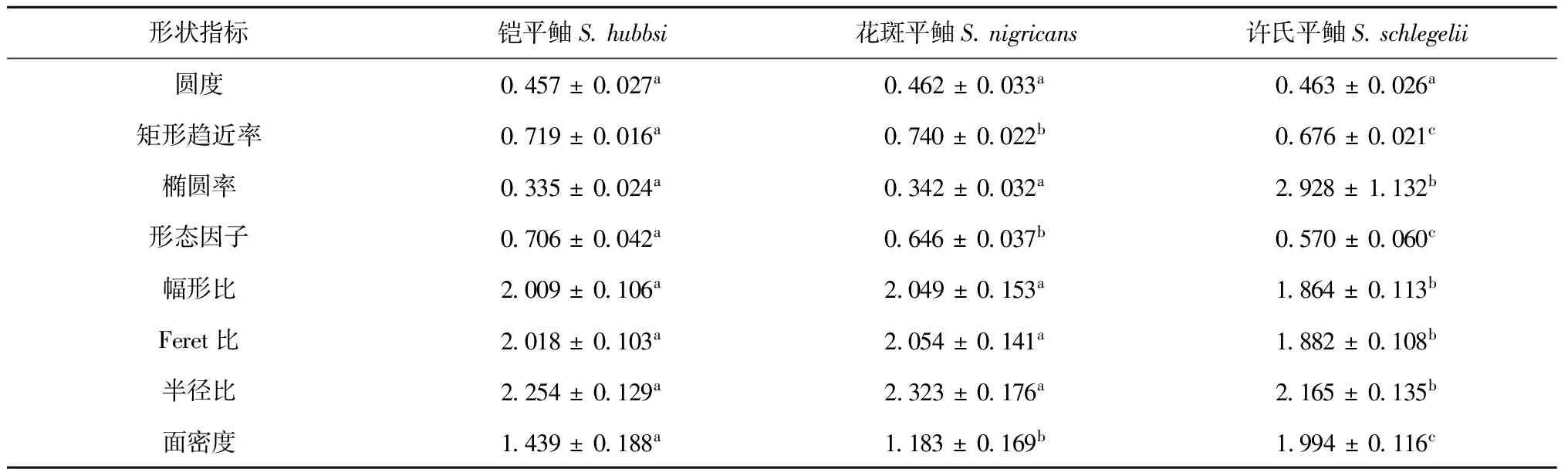
表4 平鲉屬魚類8個耳石形狀指標均值及單因素方差分析結果
注:表中耳石形狀信息為原始數據,顯著性判定為依據校正后的數據。上述字母不同表示差異顯著,字母相同表示差異不顯著(p<0.05)。
對3種魚類的37個傅里葉特征系數進行單因素方差分析,將兩兩比較具有差異性顯著的特征系數的個數及其所占總系數(37)的比例列入表5。可以看出,3種魚類在多個特征系數上的差異具有明顯的統計意義。其中,鎧平鲉和許氏平鲉間的差異最大,有16個特征系數存在顯著性差異,占總系數的43.2%;鎧平鲉和花斑平鲉之間的差異較小,有12個特征系數存在差異,占總系數的32.4%。

表5 平鲉屬魚類耳石傅里葉特征系數顯著個數及比例
3.3 種類識別
分別采用3種方式進行平鲉屬魚類種類識別的判別分析結果如表6所示。交叉驗證結果表明:
僅耳石形狀指標,典則判別函數一能夠解釋91.9%的魚種間差異,可以有效地將許氏平鲉與鎧平鲉和花斑平鲉區分開;典則判別函數二能夠解釋8.1%的魚種間差異,可以將鎧平鲉與花斑平鲉區分開(見圖2a)。對判別函數一貢獻最大的參數為圓度,對判別函數二貢獻最大的參數為形態因子。僅通過耳石形狀指標進行的種類劃分,鎧平鲉與花斑平鲉之間有少許的誤判,而許氏平鲉與鎧平鲉、花斑平鲉之間無誤判出現。表6可以看出,3種魚類判別正確率為93.3%~100%,綜合判別正確率為96.2%。
僅橢圓傅里葉系數,典則判別函數一能夠解釋60.2%的魚種間差異,可以有效地將鎧平鲉和許氏平鲉區分開;典則判別函數二能夠解釋39.8%的魚種間差異,可以有效地將花斑平鲉與鎧平鲉和許氏平鲉區分開(見圖2b)。對判別函數一貢獻最大的參數為第12號傅里葉系數,對判別函數二貢獻最大的參數為第17號傅里葉系數。通過橢圓傅里葉系數進行的判別分析,3種魚類無任何誤判,各魚種判別正確率均為100%(見表6)。
結合耳石形狀指標和橢圓傅里葉系數,典則判別函數一能夠解釋97.3%的魚種間差異,可以非常有效地將許氏平鲉與鎧平鲉和花斑平鲉區分開;典則判別函數二能夠解釋2.7%的魚種間差異,可以有效地將花斑平鲉與鎧平鲉和許氏平鲉區分開(見圖2c)。對判別函數一貢獻最大的參數為圓度,對判別函數二貢獻最大的參數為Feret比。結合耳石形狀指標和橢圓傅里葉系數的判別分析,3種魚類間無錯判,判別正確率均為100%(見表6)。
綜上,僅傅里葉分析以及耳石形狀指標結合傅里葉系數的判別結果,綜合判別正確率均高達100%;僅采用耳石形狀指標進行的判別分析,綜合判別率雖未達100%,但亦達較高正確率。
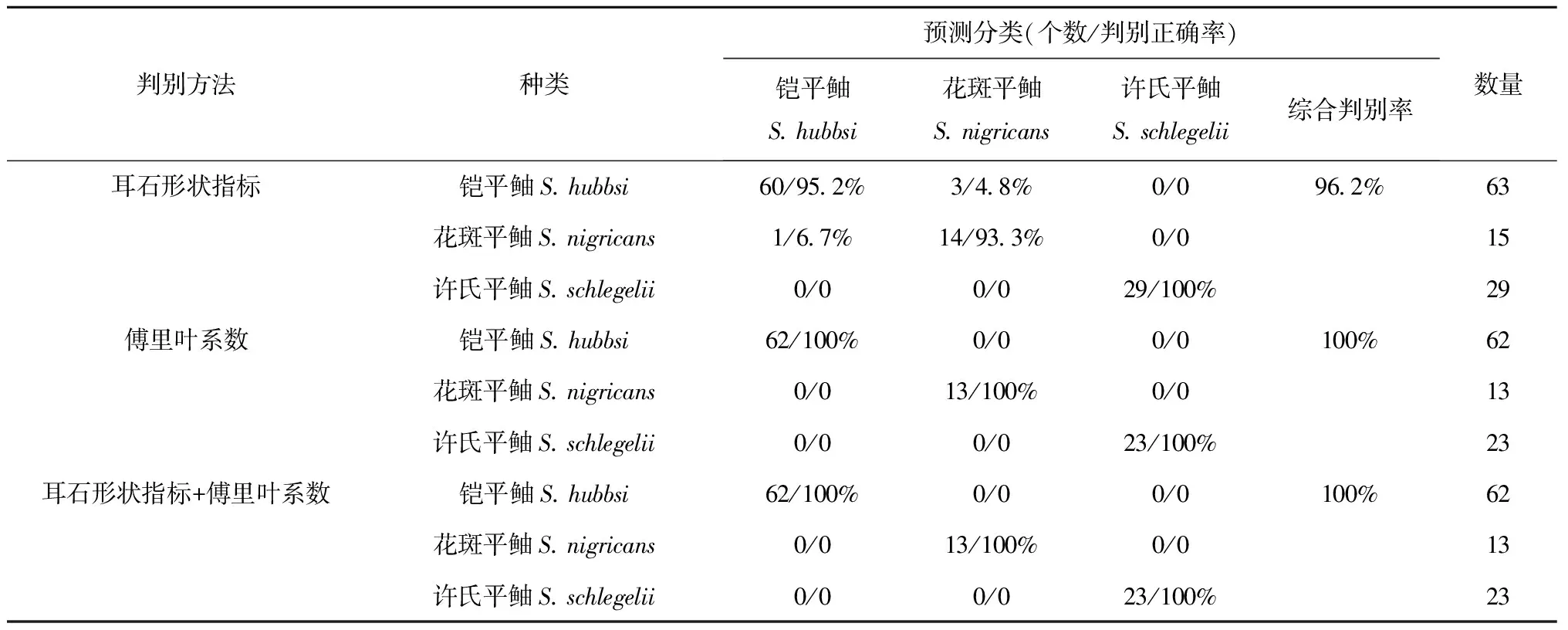
表6 基于平鲉屬3種魚類耳石形態的交叉檢驗結果
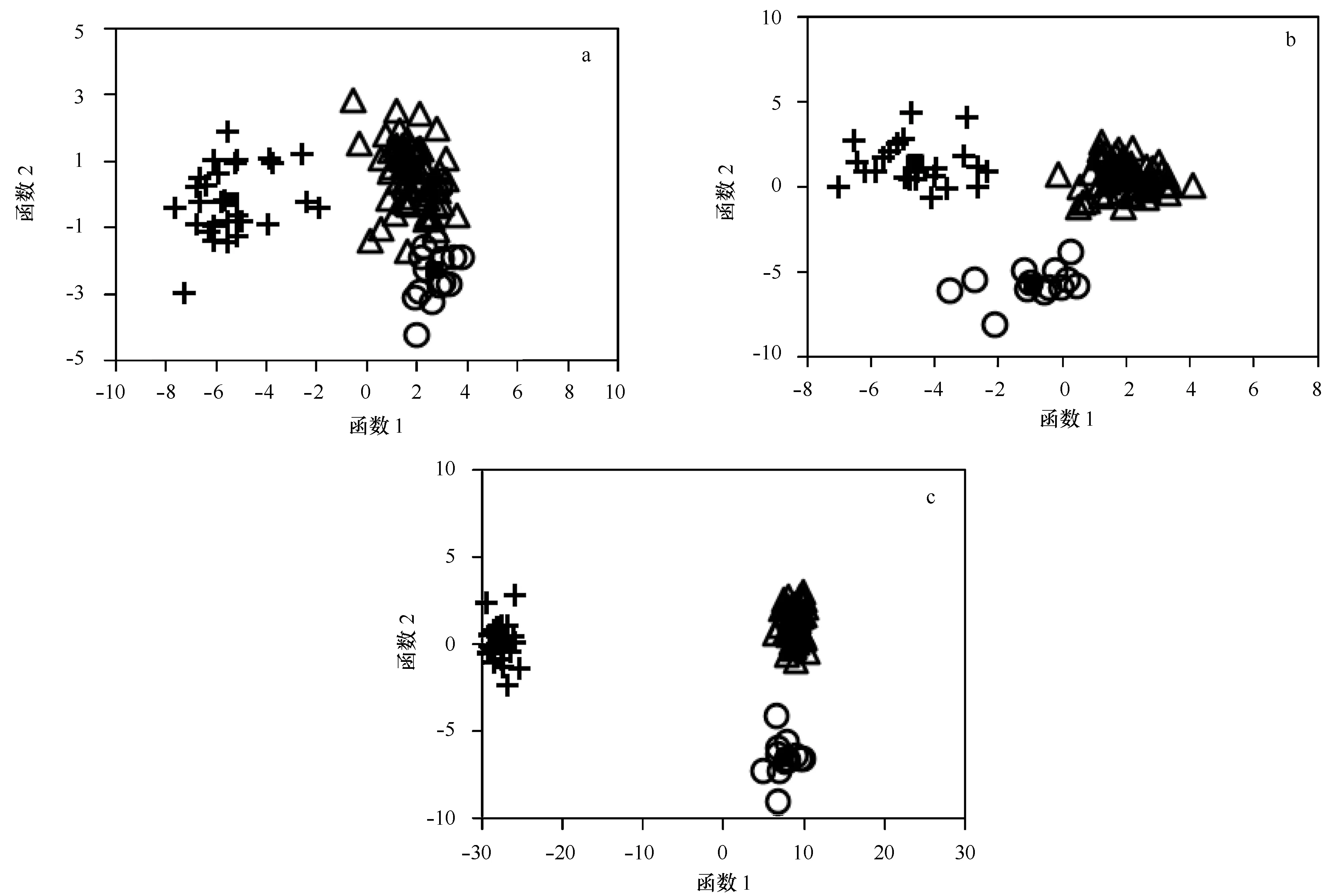
圖2 基于平鲉屬耳石形態的前2個判別函數值的分析結果散點圖Fig.2 Scatter plot of scores based on the first two DFA of Sebastes sagittae
3.4 耳石長介于4~7 mm樣本的對比鑒別結果
對耳石長介于4~7 mm的3種魚類進行Pearson相關分析,校正后的耳石參數進行后續的判別分析。亦采用3種判別方式進行3種魚類的交叉檢驗,結果如下:
僅耳石形狀指標,典則判別函數一能夠解釋65.3%的魚種間差異,可以有效地將鎧平鲉與許氏平鲉(小)區分開;典則判別函數二能夠解釋34.7%的魚種間差異,可以將花斑平鲉與鎧平鲉和許氏平鲉(小)區分開(見圖3a)。對判別函數一貢獻最大的參數為圓度,對判別函數二貢獻最大的參數為橢圓率。由表7可以看出,僅通過耳石形狀指標進行的種類劃分,3種魚類間均出現了誤判。61尾鎧平鲉,有1尾誤判為花斑平鲉,2尾誤判為許氏平鲉(小);14尾花斑平鲉,1尾誤判為許氏平鲉(小);45尾許氏平鲉(小),1尾誤判為花斑平鲉;可見花斑平鲉與許氏平鲉(小)耳石較鎧平鲉更相似。3種魚類的判別正確率為92.9%~97.8%,綜合判別正確率為95.3%。
僅橢圓傅里葉系數,典則判別函數一能夠解釋59.5%的魚種間差異,可以有效地將許氏平鲉(小)與鎧平鲉和花斑平鲉區分開;典則判別函數二能夠解釋40.5%的魚種間差異,可以有效地將鎧平鲉與花斑平鲉區分開(見圖3b)。對判別函數一貢獻最大的參數為第4號傅里葉系數,對判別函數二貢獻最大的參數為第15號傅里葉系數。通過橢圓傅里葉系數進行的判別分析,3種魚類無任何誤判,各魚種判別正確率均為100%(表7)。
結合耳石形狀指標和橢圓傅里葉系數,典則判別函數一能夠解釋60.8%的魚種間差異,可以非常有效地將鎧平鲉與花斑平鲉和許氏平鲉(小)區分開;典則判別函數二能夠解釋39.2%的魚種間差異,可以有效地將花斑平鲉與許氏平鲉(小)區分開(見圖3c)。對判別函數一貢獻最大的參數為圓度,對判別函數二貢獻最大的參數為橢圓率。結合耳石形狀指標和橢圓傅里葉系數的判別分析:60尾鎧平鲉有2尾錯判為許氏平鲉(小),鎧平鲉的判別正確率為96.7%;花斑平鲉和許氏平鲉(小)的判別正確率均為100%(表7)。
綜上,僅使用橢圓傅里葉系數的判別分析,綜合判別正確率高達100%;僅采用耳石形狀指標以及耳石形狀指標和傅里葉系數聯合判別分析,綜合判別率雖未達100%,但亦達較高正確率。
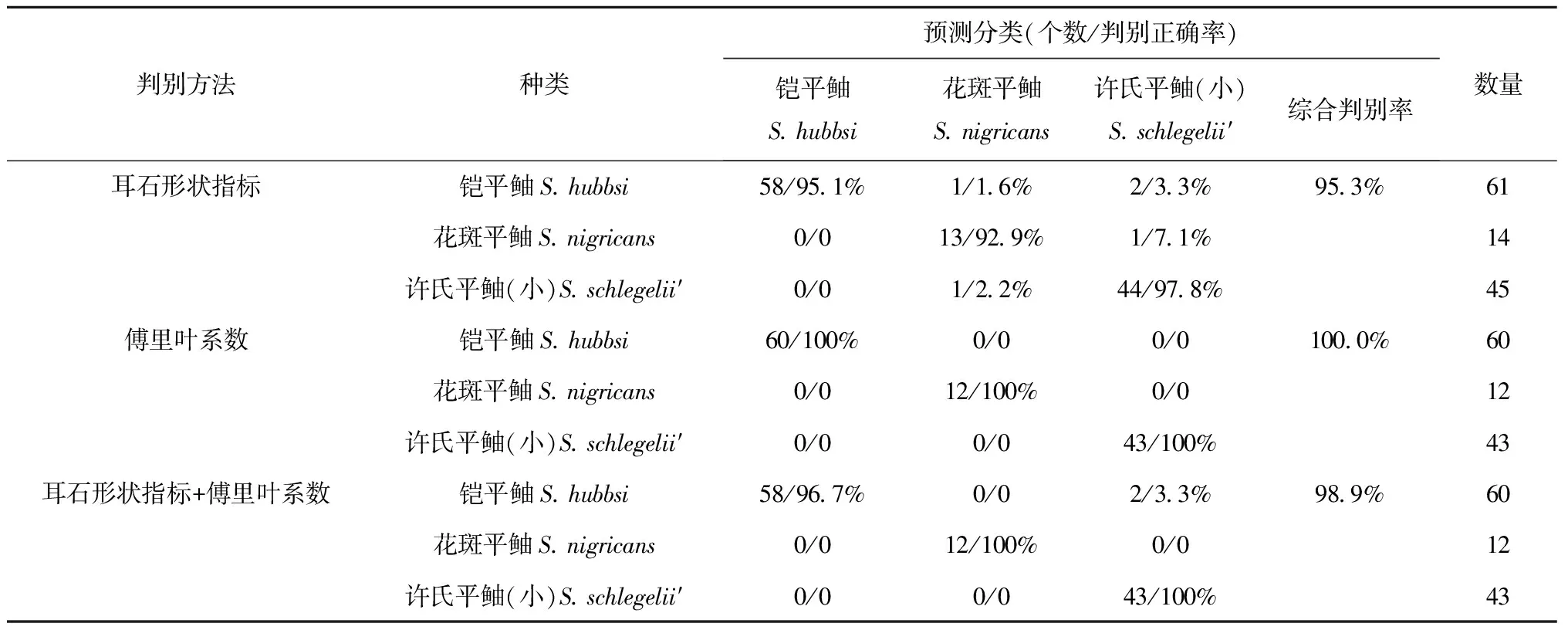
表7 耳石長4~7 mm的3種魚類耳石形態的交叉檢驗結果
注:文中以S.schlegelii′表示許氏平鲉(小)的拉丁名。
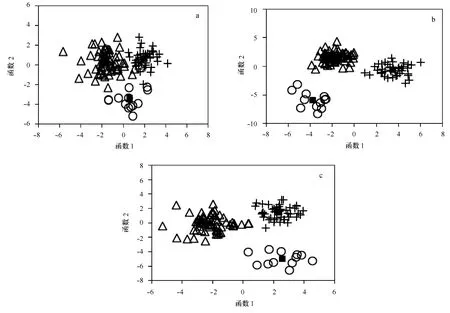
圖3 耳石長4~7 mm的3種魚類判別分析結果散點圖Fig.3 Scatter plot of scores based on the first two DFA of the three Sebastes whose sagittae length were 4 to 7 mm
4 討論
4.1 樣本選擇及耳石形態參數的選擇與校正
研究發現,鎧平鲉在2齡時初次達到性成熟[34]、許氏平鲉在2~3齡時達性成熟[35]。本文首先選取2齡及以上樣本探究基于耳石形態鑒別平鲉屬魚類的可行性,原則上排除了幼魚耳石形狀不穩定對實驗結果的影響,且3種魚類不同體長間耳石形狀指標的單因素方差分析結果顯示,2齡及以上3種魚類的耳石形態已趨于穩定;另外,根據實驗采集的樣本和文獻報道[27]發現,近岸捕撈的許氏平鲉多為體長80~180 mm不足1齡的幼魚,此體長階段的耳石長恰為4~7 mm,因此文中對比加入耳石長介于4~7 mm的小個體許氏平鲉樣本,將耳石長介于4~7 mm的3種平鲉屬魚類單獨進行基于耳石形態的判別分析,充分探討了針對不同生長階段3種魚類種類鑒別的可行性及分析方法的有效性。
同時,研究樣本均采自青島膠州灣海域,一定程度上克服了地理環境不同對耳石形態的影響,降低了環境因素對實驗結果的干擾。由于花斑平鲉在我國僅分布在黃海海域且資源量較少,故采集到的樣本數量有限,今后應進一步增加花斑平鲉耳石樣本數量,以提高實驗結果的可靠性。
耳石形態特征主要受遺傳因素的影響,魚類的生物學特征(性別、耳石位置、個體大小等)亦對耳石的形態特征產生一定影響,秦巖等[2]報道了鲬魚(Platycephalusindicus)雌雄個體間矢耳石形態特征存在差異,陸化杰等[36]指出不同個體、不同性別間阿根廷滑柔魚(Illexargentinus)耳石形狀參數存在顯著差異。本文逐一檢驗了3種平鲉屬魚類的左、右耳石以及雌、雄個體間耳石的形態差異性。相關研究表明,當影響因素如性別、耳石位置等對耳石形態影響水平不顯著時,若樣本量充足則可去除該因素,若樣本量不足則應匯總樣本進行分析[37]。鑒于本研究樣本數較少,且3種魚類耳石在雌雄個體間均無顯著差異,故將雌雄個體的左耳石一起作為實驗樣本納入后續分析中。
對于耳石參數的選擇,包含了稱量指標耳石質量、由耳石大小變量進行轉換的耳石形狀指標和耳石輪廓變量傅里葉系數,引入耳石質量信息增加了耳石的三維特征,結果表明耳石質量信息的加入可以提高判別率[20]。以耳石形態鑒別3種魚類,為實現無偏分析,鑒于2齡及以上的3種平鲉屬魚類體長差異較大,故對其進行體長效應的校正;而耳石長介于4~7 mm的3種魚類,體長差異較小,故選擇通過耳石長進行校正。校正后的相關指標用于后續的統計分析中,排除了魚體大小、耳石大小對判別結果的影響。
4.2 多元統計結果的分析
8個耳石形狀指標的單因子方差分析結果(見表4)顯示,3種平鲉屬魚類間耳石形狀指標存在顯著差異,尤其是鎧平鲉與許氏平鲉、花斑平鲉與許氏平鲉的耳石,有7個形狀指標存在顯著差異。對3種魚類的37個橢圓傅里葉特征系數進行兩兩比較,傅里葉系數差異顯著的個數占總特征系數的比例均達30%以上,其中鎧平鲉與許氏平鲉之間的差異高達43.2%(見表5)。綜合可見,2種方法的方差分析結果基本一致,鎧平鲉與許氏平鲉的耳石形態差異最大,鎧平鲉與花斑平鲉則差異最小。其中,差異不顯著的參數為平鲉屬魚類耳石的共有特征,如3種魚類間耳石圓度差異不顯著,表明該圓度范圍即為3種魚類耳石的共有特征;而對比分析中差異顯著的參數為耳石的種間差異所在,即為基于耳石形態鑒別平鲉屬魚類提供了可能。
本研究采用耳石形狀指標、橢圓傅里葉系數以及耳石形狀指標與橢圓傅里葉系數聯合判別法進行3種平鲉屬魚類的鑒別,結果顯示:(1)3種判別方式對于3種魚類(包括大小規格的許氏平鲉)的綜合判別率均高達95%以上,且判別函數所作散點圖可明顯將3種魚類區分開來(見圖2,圖3),可見基于耳石形態進行平鲉屬魚類的鑒別是可行的,3種判別方式均可作為平鲉屬魚類種類鑒別的輔助方法。(2)僅使用耳石形狀指標進行的判別分析結果顯示,大個體許氏平鲉與鎧平鲉和花斑平鲉間無誤判出現,表明其與鎧平鲉和花斑平鲉間的耳石形態差異較大;而小個體的許氏平鲉與鎧平鲉、花斑平鲉均有一定程度的誤判(最大誤判率為3.3%),說明低于1齡的許氏平鲉(小個體)與鎧平鲉、花斑平鲉的耳石形態差異較之大個體許氏平鲉相對較小。這表明魚體生長過程中許氏平鲉耳石發生一定的形態變化,亦驗證了隨著魚體的生長,耳石亦經歷生長、發育和穩定過程的論點。而圖2、圖3的散點圖顯示,無論許氏平鲉處于哪個生長階段,依據耳石均能將3種平鲉屬魚類區分開來,表明基于耳石形態進行不同生長階段的平鲉屬魚類種類鑒別是可行性的。(3)比較3種判別方式對平鲉屬3種魚類的判別結果,由表6、表7(包含大小規格的許氏平鲉)可知,僅使用橢圓傅里葉系數的判別分析均能實現100%的判別正確率,相較于其他2種判別方法,該研究方法在探討基于耳石形態的3種平鲉屬魚類的鑒別中更具優勢。如李輝華等[38]曾證實對于長江鳳鱭(Coilianasus)和刀鱭(Coiliamystus)的種間和種內鑒別,傅里葉分析法較傳統形態測量法更具優勢。然而,傅里葉系數雖可以準確的定量表達耳石外輪廓,但是與魚類種間的實際耳石形狀差異較難聯系起來,難以快速找出不同種間的耳石在統計上差異顯著而實際上微小的形狀差異所在[33,39]。本研究顯示,結合耳石形狀指標法和橢圓傅里葉分析2種方法,對3種平鲉屬魚類的綜合判別率亦高達99%,且由耳石形狀指標所得的單因素方差分析結果可知(見表4),3種魚類耳石在矩形趨近率、形態因子、面密度等耳石形狀指標上存在顯著差異,因此,耳石形狀指標法和橢圓傅里葉分析的聯合使用是基于耳石形態進行平鲉屬魚類種類鑒別的有效手段,該方法既能快速找出3種魚類間耳石形態的差異所在,又能實現較高的種間判別正確率。研究表明,耳石形狀指標結合橢圓傅里葉系數對鮟鱇(Lophiuspiscatorius)[29]、鲯鰍(Coryphaenahippurus)[40]、黃姑魚(Argyrosomusjaponicus)[41]等種群鑒別時效果良好,2種方法聯合使用在魚類耳石的相關研究中可靠性更高。
致謝:本文依托國家自然科學基金“許氏平鲉資源增殖效果評估新方法及其應用基礎研究”。中國海洋大學漁業生態學實驗室為本項目的開展給予了大力支持與幫助,作者在此深表感謝。
[1] 張曉霞,葉振江,王英俊,等. 青島海域小眼綠鰭魚耳石形態的初步研究[J]. 中國海洋大學學報(自然科學版),2009,39(4): 622-626.
Zhang Xiaoxia,Ye Zhenjiang,Wang Yingjun,et al. A preliminary study on otolith morphology ofChelidonichthysspinosusin the Qingdao coastal waters[J]. Periodical of Ocean University of China(Natural Science),2009,39(4): 622-626.
[2] 秦巖,潘曉哲,高天翔. 鲬魚耳石形態多樣性研究[J]. 水生生物學報,2013,37(4): 759-765.
Qin Yan,Pan Xiaozhe,Gao Tianxiang. Study on morphological diversity of sagittae ofPlatycephalusindicus[J]. Aquatic Biology,2013,37(4): 759-765.
[3] Campana S E. Photographic atlas of fish otoliths of the Northwest Atlantic Ocean[M]. Ottawa:NRC Research Press,2004.
[4] Fitch J E,Brownell Jr R L. Fish otoliths in cetacean stomachs and their importance in interpreting feeding habits[J]. Journal of the Fisheries Board of Canada,1968,25(12): 2561-2574.
[5] Lychakov D V,Rebane Y T. Otolith regularities[J]. Hearing Research,2000,143(1): 83-102.
[6] Vignon M. Ontogenetic trajectories of otolith shape during shift in habitat use: Interaction between otolith growth and environment[J]. Journal of Experimental Marine Biology and Ecology,2012,420: 26-32.
[7] Gagliano M,McCormick M I. Feeding history influences otolith shape in tropical fish[J]. Marine Ecology Progress Series,2004,278: 291-296.
[8] Vignon M,Morat F. Environmental and genetic determinant of otolith shape revealed by a non-indigenous tropical fish[J]. Marine Ecology Progress Series,2010,411: 231-241.
[9] Agüera A,Brophy D. Use of saggital otolith shape analysis to discriminate Northeast Atlantic and Western Mediterranean stocks of Atlantic saury,Scomberesoxsaurussaurus(Walbaum)[J]. Fisheries Research,2011,110(3): 465-471.
[10] Morales-Nin B. Review of the growth regulation processes of otolith daily increment formation[J]. Fisheries Research,2000,46(1): 53-67.
[11] Maisey J G. Notes on the structure and phylogeny of vertebrate otoliths[J]. Copeia,1987(2): 495-499.
[12] 葉振江. 中國海洋魚類耳石形態學分析及應用研究[D]. 青島:中國海洋大學,2010.
Ye Zhenjiang. Study on otolith shape,taxonomy and application for fishes in China sea[D]. Qingdao:Ocean University of China,2010.
[13] Begg G A,Overholtz W J,Munroe N J. The use of internal otolith morphometrics for identification of haddock (Melanogrammusaeglefinus) stocks on Georges Bank[J]. Fishery Bulletin,2001,99(1):1-14.
[14] Tuset V M,Rosin P L,Lombarte A. Sagittal otolith shape used in the identification of fishes of the genusSerranus[J]. Fisheries Research,2006,81(2): 316-325.
[15] 張國華,但勝國,苗志國,等. 六種鯉科魚類耳石形態以及在種類和群體識別中的應用[J]. 水生生物學報,1999,23(6): 683-688.
Zhang Guohua,Dan Shengguo,Miao Zhiguo,et al. Otolith morphology of six Cyprinid species with the use in species and stock discriminations[J]. Acta Hydrobiological Sinica,1999,23(6): 683-688.
[16] 郭弘藝,魏凱,唐文喬,等. 基于矢耳石形態特征的中國鱭屬魚類種類識別[J]. 動物分類學報,2010 (1): 127-134.
Guo Hongyi,Wei Kai,Tang Wenqiao,et al. Sibling species discrimination for Chinese genus ofCoiliafishes based on sagittal otolith morphology[J]. Acta Zootaxonomica Sinica,2010 (1): 127-134.
[17] L’Abée-Lund J H. Otolith shape discriminates between juvenile Atlantic salmon,SalmosalarL.,and brown trout,SalmotruttaL.[J]. Journal of fish biology,1988,33(6): 899-903.
[18] 曾祥波,張國華. 基于傅立葉耳石形態分析法對幾種家魚仔稚魚的種類判別分析[J]. 中國水產科學,2012,6: 970-977.
Zeng Xiangbo,Zhang Guohua. Species identification at the larval and juvenile stages for several Chinese domestic fishes by elliptical Fourier analysis of otolith form[J]. Journal of Fishery Sciences of China,2012,6: 970-977.
[19] Ponton D. Is geometric morphometrics efficient for comparing otolith shape of different fish species?[J]. Journal of Morphology,2006,267(6): 750-757.
[20] 潘曉哲,高天翔. 基于耳石形態的鱚屬魚類鑒別[J]. 動物分類學報,2010,35(4): 799-805.
Pan Xiaozhe,Gao Tianxiang. Sagittal otolith shape used in the discrimination of fishes of the GenusSillagoin China[J]. Acta Zootaxonomica Sinica,2010,35(4): 799-805.
[21] Lord C,Morat F,Lecomte-Finiger R,et al. Otolith shape analysis for threeSicyopterus(Teleostei: Gobioidei: Sicydiinae) species from New Caledonia and Vanuatu[J]. Environmental Biology of Fishes,2012,93(2): 209-222.
[22] Tuset V M,Lozano I J,Gonzalez J A,et al. Shape indices to identify regional differences in otolith morphology of comber,Serranuscabrilla(L.,1758)[J]. Journal of Applied Ichthyology,2003,19(2): 88-93.
[23] Stransky C,Murta A G,Schlickeisen J,et al. Otolith shape analysis as a tool for stock separation of horse mackerel (Trachurustrachurus) in the Northeast Atlantic and Mediterranean[J]. Fisheries Research,2008,89(2): 159-166.
[24] Tracey S R,Lyle J M,Duhamel G. Application of elliptical Fourier analysis of otolith form as a tool for stock identification[J]. Fisheries Research,2006,77(2): 138-147.
[25] Begg G A,Brown R W. Stock identification of haddockMelanogrammusaeglefinuson Georges Bank based on otolith shape analysis[J]. Transactions of the American Fisheries Society,2000,129(4): 935-945.
[26] Galley E A,Wright P J,Gibb F M. Combined methods of otolith shape analysis improve identification of spawning areas of Atlantic cod[J]. ICES Journal of Marine Science: Journal du Conseil,2006,63(9): 1710-1717.
[27] 徐勝勇,張輝,潘曉哲,等. 許氏平鲉群體耳石形態學比較[J]. 中國海洋大學學報 (自然科學版),2012,42(11):54-61.
Xu Shengyong,Zhang Hui,Pan Xiaozhe,et al. Morphological comparison of the otolith ofSebastesschlegeliipopulations[J]. Periodical of Ocean University of China(Natural Science),2012,42(11):54-61.
[28] Tuset V M,Lombarte A,Gonzalez J A,et al. Comparative morphology of the sagittal otolith inSerranusspp.[J]. Journal of fish biology,2003,63(6): 1491-1504.
[30] 王英俊. 傅里葉分析在魚類耳石形態學中的應用研究[D]. 青島:中國海洋大學,2010.
Wang Yingjun. The application of Fourier Analysis in the research of otolith morphology[D]. Qingdao:Ocean University of China,2010.
[31] Kuhl F P,Giardina C R. Elliptic Fourier features of a closed contour[J]. Computer Graphics and Image Processing,1982,18(3): 236-258.
[32] 王英俊,葉振江,劉群,等. 細條天竺魚 (Apogonichthyslineatus) 與黑鰓天竺魚 (Apogonichthysarafurae) 耳石形態識別的初步研究[J]. 海洋與湖沼,2010,41(2): 281-284.
Wang Yingjun,Ye Zhenjiang,Liu Qun,et al. Using otolith to identify two fish species:ApogonichthyslineatusandApogonichthysarafurae[J]. Oceanologica et Limnological Sinica,2010,41(2): 281-284.
[33] 竇碩增,于鑫,曹亮. 魚類矢耳石形態分析及其在群體識別中的應用實例研究[J]. 海洋與湖沼,2012,43(4): 702-712.
Dou Shuozeng,Yu Xin,Cao Liang. Otolith shape analysis and its application in fish stock discrimination: a case study[J]. Oceanologia et Limnologia Sinica,2012,43(4): 702-712.
[34] 王英俊,葉振江,張弛,等. 車牛山島鎧平鲉繁殖群體生物學研究[J]. 中國海洋大學學報(自然科學版),2011,41(12): 46-52.
Wang Yingjun,Ye Zhenjiang,Zhang Chi,et al. The fisheries biology ofSebasteshubbsiin Che-Niushan island[J]. Periodical of Ocean University of China(Natural Science),2011,41(12): 46-52.
[35] 張輝. 西北太平洋兩種卵胎生魚類 (許氏平鲉和褐菖鲉) 的分子系統地理學研究[D]. 青島:中國海洋大學,2013.
Zhang Hui. Molecular phylogeography of two marine ovoviviparous fishes in northwestern Pacific[D]. Qingdao:Ocean University of China,2013.
[36] 陸化杰,陳新軍,劉必林. 個體差異對西南大西洋阿根廷滑柔魚耳石形態的影響[J]. 水產學報,2011,35(2): 247-253.
Lu Huajie,Chen Xinjun,Liu Bilin. Effects of individual difference on statolith morphology ofIllexargentinusin the southwest Atlantic Ocean[J]. Journal of Fisheries of China,2011,35(2): 247-253.
[37] 于鑫,曹亮,南鷗,等. 基于矢耳石形態分析的鳳鱭 (Coiliamystus) 群體識別研究[J]. 海洋與湖沼,2013,44(3):768-774.
Yu Xin,Cao Liang,Nan Ou,et al. Stock identification ofCoiliamystususing otolith shape analysis[J]. Oceanologia et Limnologia Sinica,2013,44(3):768-774.
[38] 李輝華,郭弘藝,唐文喬,等. 兩種耳石分析法在鱭屬種間和種群間識別效果的比較研究[J]. 淡水漁業,2013,43(1): 14-18.
Li Huihua,Guo Hongyi,Tang Wenqiao,et al. Comparative study of two otolith shape analysis for genusCoillaspecies and stocks identification[J]. Freshwater Fisheries,2013,43(1): 14-18.
[39] DeVries D A,Grimes C B,Prager M H. Using otolith shape analysis to distinguish eastern Gulf of Mexico and Atlantic Ocean stocks of king mackerel[J]. Fisheries Research,2002,57(1): 51-62.
[40] Duarte-Neto P,Lessa R,Stosic B,et al. The use of sagittal otoliths in discriminating stocks of common dolphinfish (Coryphaenahippurus) off northeastern Brazil using multishape descriptors[J]. ICES Journal of Marine Science: Journal du Conseil,2008,65(7): 1144-1152.
[41] Ferguson G J,Ward T M,Gillanders B M. Otolith shape and elemental composition: Complementary tools for stock discrimination of mulloway (Argyrosomusjaponicus) in southern Australia[J]. Fisheries Research,2011,110(1): 75-83.
Sagittae morphology used in the discrimination of the fish ofSebastesand the comparison of sagittal research method
Zhang Fengxia1,Zhang Xiumei1,Qin Lezheng1,Li Wentao1
(1.TheKeyLaboratoryofMariculture,MinistryofEducation,OceanUniversityofChina,Qingdao266003,China)
The sagittae morphological characteristics of three speciesSebasteshubbsi,S.nigricansandS.schlegeliifrom Jiaozhou Bay,Qingdao,were analyzed to test the hypothesis that the threeSebastesspecies could be identified by their sagittae morphology and to find out which method was more appropriate. Eight otolith shape indices and 37 elliptical Fourier coefficients (eFcs) were analyzed,and results showed that the difference in the sagittae morphology betweenS.hubbsiandS.schlegeliiwere the largest,and those betweenS.hubbsiandS.nigricanswere the least. In the discriminant analyses of the three species,discriminating rates of more than 95% were obtained and they were clearly sorted in scatter plots when using the otolith shape indices,the eFcs,as well as a combination of the two. When applied to the three species of which otolith was 4 to 7 mm and those were more than two years old,the species identification based on the otolith morphology was proved to be feasible: when only using the eFcs,a 100% of discriminating rate was obtained but this method couldn’t quickly locate the shape which was tiny in fact but significant difference among the sagittae of the threeSebastesin statistics. Combing otolith shape indices and the eFcs could not only obtain a high classification rate of about 99%,but also identify the differences in the rectangularity,form-factor and surface density among the threeSebastessagittae. The results showed that combing shape indices analysis and elliptical Fourier analysis was the effective method for species discrimination of genusSebastesand it can find out the differences of the sagittae of the threeSebastes.
Sebastes; sagittae morphology; otolith shape indices; elliptical Fourier coefficients; species discrimination
10.3969/j.issn.0253-4193.2015.04.003
2014-04-20;
2014-12-28。
國家自然科學基金(31172447,41176117);國家海洋公益性行業科研專項(201305043)。
張鳳俠(1990—),女,安徽省阜陽市人,從事資源生物學研究。 Email: fxzhang1003@163.com
*通信作者:張秀梅,教授。E-mail: xiumei1227@163.com
S917.4
A
0253-4193(2015)04-0028-12
張鳳俠, 張秀梅, 覃樂政,等.基于矢耳石形態的平鲉屬(Sebastes)魚類鑒別及研究方法比較[J].海洋學報,2015,37(4):28—39,
Zhang Fengxia, Zhang Xiumei, Qin Lezheng, et al. Sagittae morphology used in the discrimination of the fish ofSebastesand the comparison of sagittal research method[J]. Haiyang Xuebao,2015,37(4):28—39, doi: 10.3969/j.issn.0253-4193.2015.04.003
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
