漢豐湖底棲動物與環境因子間相關性初步研究
2015-10-21 18:26:54彭建華鄭志偉鄒曦池仕運張志永潘曉潔萬成炎
環境影響評價 2015年5期
關鍵詞:相關性
彭建華 鄭志偉 鄒曦 池仕運 張志永 潘曉潔 萬成炎

摘要:2013年3、6、9、12月對漢豐湖調節壩未運行前的底棲動物和環境因子進行了4次調查,調查結果表明:漢豐湖底棲動物平均密度為325.17±140.22 ind./m2。各采樣點平均水溫為20.62±5.00℃、電導率為493.84±175.51 μS/cm、溶解性總固體為0.370±0.143 mg/L、溶解氧為8.03±0.49 mg/L、TN為1.584±0.323 mg/L、氨氮為0.392±0.229 mg/L、硝氮為0.912±0.263 mg/L、TP為0.083±0.029 mg/L、磷酸鹽為0.049±0.021 mg/L、高錳酸鉀指數為3.771±1.57 mg/L、葉綠素為19.15±13.03 mg/L、水深均值為5.63±4.59 m、流速為0.048±0.056 m/s、流量為1.25±1.47 m3/s。應用典范對應分析(CCA),環境變量之間存在交互作用,硝氮(F=6.49,P=0.002)、水深(F=4.21,P=0.010)、總磷(F=3.64,P=0.016)是影響漢豐湖底棲動物密度的主要因素;在NO-3N梯度上,軟體動物的最適值比水生昆蟲和寡毛類高,在水深和TP梯度上,寡毛類的最適值高于軟體動物和水生昆蟲。同時對漢豐湖調節壩開始運行,湖體水位穩定后,影響底棲動物的主要環境因子進行了分析預測。
關鍵詞:底棲動物;環境因子;相關性;漢豐湖
DOI: 10.14068/j.ceia.2015.05.015
中圖分類號:X174 文獻標識碼:A 文章編號:2095-6444(2015)05-0063-06
底棲動物是指生活史的全部或大部分時間生活于水體底部的水生動物群,是河流、湖泊、水庫生態系統的重要類群[1]。它們既可利用淤泥、腐殖質中的有機物質,起到凈化水質的作用;也可被其他水生動物利用,提升經濟水生動物的產量;還可作為環境指示生物,評價水域生態環境[1-2]。底棲動物的種類組成和豐度與氣候因素(如降水、霜凍等)、物理因素(如底質、水深等)、化學因素(如溶氧、化學耗氧量、重金屬等)、水文因素(如流速、流量)、生物因素(如水生植物、捕食)以及人類活動(如引水式電站)等密不可分[1-4],研究底棲動物與環境因子之間的關系,對探索水域生態環境有著重要意義。
漢豐湖位于重慶市開縣境內,三峽水庫小江(又名澎溪河)支流中上游,水域面積14.8 km2,具有美化環境、凈化水質的作用。漢豐湖水位高低受調節壩控制(該閘壩已建成,但還未正式運行),同時還受三峽水庫運行的影響。當三峽水位上升至152.5 m時,漢豐湖區域開始受三峽尾水影響而蓄水;當三峽水庫水位上漲至172.8 m時,水位調節壩閘門全開,漢豐湖水位與三峽水庫水位同步運行;當三峽水庫水位下降至172.8 m時,調節壩下閘,漢豐湖開始蓄水[5]。關于漢豐湖的底棲動物,目前只有群落結構方面的報道[6]。典范對應分析(CCA)是一種非線性多元直接梯度分析方法,其分析結果直觀,包含的信息量大,常用作水生生物群落組成與水環境因子的相關關系研究[7]。本文應用CCA分析研究漢豐湖底棲動物與環境因子之間的關系,以期為漢豐湖的環境保護和污染防治提供理論依據。
1 材料與方法
1.1 斷面設置和采樣時間
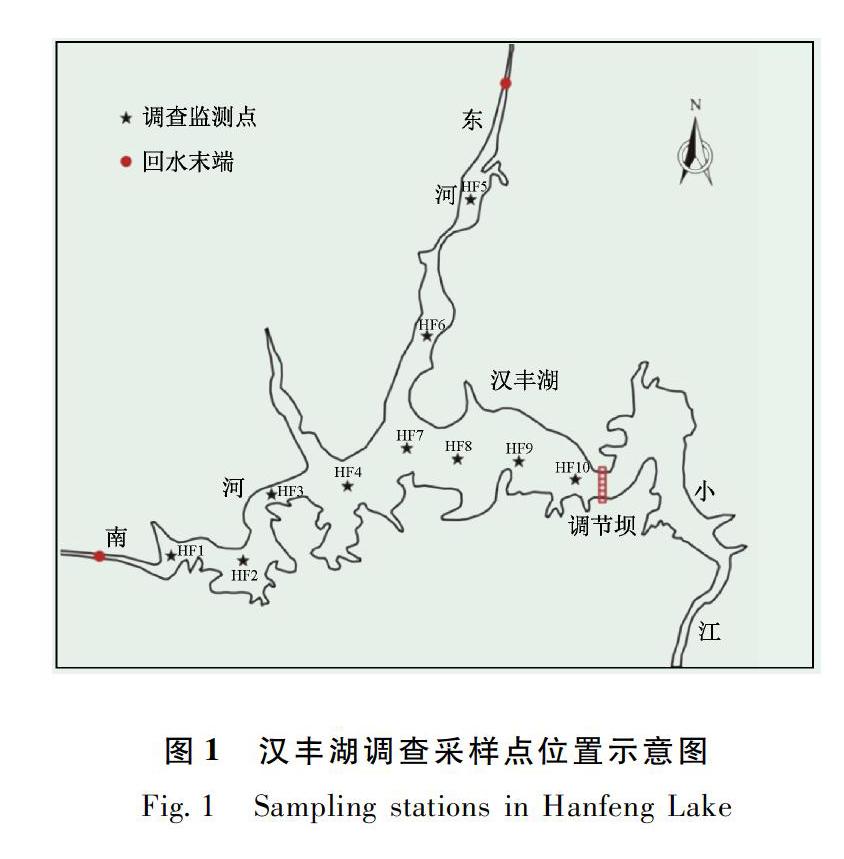
根據漢豐湖形態,共設置10個采樣點,其中HF1和HF2位于南河庫灣,HF5和HF6位于東河庫灣(圖1);采樣分4個季度進行,分別為2013年春季(3月)、夏季(6月)、秋季(9月)和冬季(12月)[6]。
1.2 樣品采集、處理與分析測試
底棲動物的樣品采集與分類鑒定、計數參照文獻[6]的方法。水體理化指標包括水溫、電導率、溶解性總固體(TDS)、溶解氧(DO)、pH、總氮(TN)、氨氮(NH3N)、硝氮(NO-3N)、總磷(TP)、磷酸鹽(PO-4P)、高錳酸鉀指數、葉綠素(Chla)等。其中,水溫、電導率、溶解氧、pH值用YSI水質分析儀現場測定,總氮、氨氮、硝氮、總磷、磷酸鹽、高錳酸鉀指數、葉綠素指標均按照《水和廢水監測分析方法》[8]進行。水文參數水深、流速、流量采用ADP聲學多普勒剖面流速儀現場測量。
1.3 數據分析
利用Canoco for windows 4.5先對物種數據進行除趨勢對應分析(Detrended correspondence analysis,DCA),在Log View內,可以找到每個軸的梯度長度,如果這4個軸中梯度最長超過4,則選擇單峰模型排序,如典范對應分析(CCA)更合適;如果長度小于3,則選擇線性模型,如冗余分析(RDA)比較合理;如果介于3和4之間,單峰模型和線性模型都合適,否則可能誤差較大,會丟失較多信息。由于環境變量的量綱往往不同,需要對環境變量進行平方根轉化。執行RDA過程時,選擇自動前選,進行蒙特卡洛檢驗。通過邊際影響特征值和條件影響特征值判定該環境變量對物種的影響[9-10]。
2 結果與分析
2.1 底棲動物種類和密度
漢豐湖底棲動物種類組成與密度的季節變化見參考文獻[6]。漢豐湖底棲動物平均密度為325.17±140.22 ind./m2,其中水生昆蟲密度為100.4±35.7 ind./m2,寡毛類為85.8±79.2 ind./m2,軟體動物為133.6±60.5 ind./m2,其他類群只有5.3±9.3 ind./m2(表1)。總體來看,各站位底棲動物主要由軟體動物和水生昆蟲組成,呈現自庫尾向壩前遞增趨勢。
2.2 環境因子
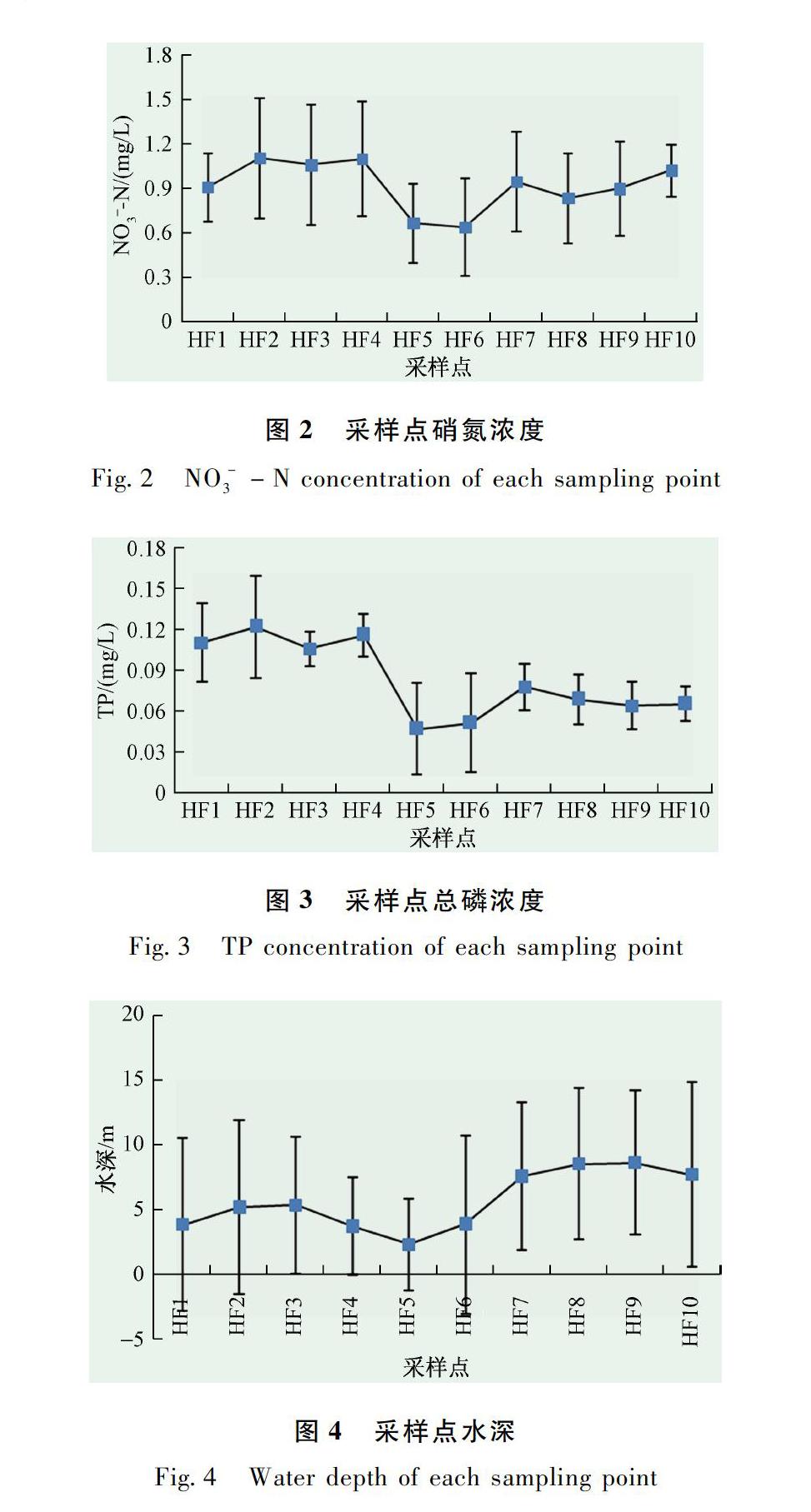
各采樣點環境因子平均值為:水溫20.62±5.00℃、電導率493.84±175.51 μS/cm、溶解性總固體0.370±0.143 mg/L、溶解氧8.03±0.49 mg/L、TN為1.584±0.323 mg/L、氨氮0.392±0.229 mg/L、硝氮0.912±0.263 mg/L(圖2)、TP為0.083±0.029 mg/L(圖3)、磷酸鹽0.049±0.021 mg/L、高錳酸鉀指數3.771±1.57 mg/L、葉綠素19.15 ±13.03 mg/L、水深均值為5.63±4.59 m(圖4)、流速為0.048±0.056 m/s、流量為1.25±1.47 m3/s。根據《地表水環境質量標準》(GB 3838—2002)判斷,漢豐湖水質溶解氧達到Ⅰ類,高錳酸鉀指數為Ⅱ類,氨氮為Ⅱ類,TP為Ⅳ類、TN為Ⅴ類。
2.3 底棲動物密度與環境因子相關性
對底棲動物密度進行的DCA分析表明,最大軸的梯度長度均為3.063,本文選擇CCA分析底棲動物密度與環境因子的關系。
CCA第一和第二排序軸可以解釋底棲動物動物密度的比例分別為34.6%和15.1%,總典范特征值為0.633。NO-3N的邊際影響特征值最大(0.18),也排在條件影響變量中的第一位(0.18),說明漢豐湖底棲動物密度大小受NO-3N的影響最大(表2和圖2)。水溫(0.15)和溶解性總固體(0.13)的邊際影響次之。環境變量中,NO-3N(F=6.49,P=0.002)、水深(F=4.21,P=0.010)、TP(F=3.64,P=0.016)通過了蒙特卡洛檢驗(圖2),說明環境變量之間存在交互作用,影響底棲動物密度的環境因子主要是NO-3N、水深和TP。
在單峰排序圖內,物種點之間的距離為卡方距離,可代表不同物種的空間分布差異,從物種點到數量型環境因子箭頭的投影點的位置次序可以代表物種在該環境因子最適值(Optimum,物種密度達到最大值時的環境因子值)的排序[9-10]。因此,從圖中可以看出,水生昆蟲與軟體動物空間分布差異不明顯。在NO-3N梯度上,軟體動物的最適值比水生昆蟲和寡毛類的高。在水深和TP梯度上,寡毛類的最適值高于軟體動物和水生昆蟲。
特別說明:作圖時,把圖中的英文轉換為漢字,需要轉換的英文及對應關系如下:Temperature,水溫;Cond,電導率;KMnO4,高錳酸鉀指數;Depth,水深;Velocity,流速;Flow,流量;Insecta,水生昆蟲;Oligachata,寡毛類;Mollusca,軟體動物;Others,其他類群。
3 討論
多數底棲動物具有區域性強、遷移能力弱等特點,外界環境的許多方面影響底棲動物的種類分布、群落結構[11-12]。氣候變化引起流域水文特征、水體理化等環境特征變化,這些環境因素的變化又將影響底棲動物的生長、繁殖和種群演替,直至群落結構的變化[3]。物理因素方面,在多類型和高復雜性的生境中,底棲動物的類群組成復雜性也高[13];隨著河流級別的升高,底棲動物攝食功能群從以撕食者和集食者為主變為以收集者為主[14];同一河流不同河段的底棲動物群落組成及多樣性有所差別[2]。水體理化因子方面,因溫度因素,大型底棲動物有暖水種、溫水種、冷水種的區分[15];熱帶河流中大型底棲動物的物種數要高于溫帶河流,而溫帶河流又高于寒溫帶和寒帶河流[16];偏堿性水體利于多數大型底棲動物生存,酸性水體則反之[17];營養物質(尤其是氮和磷的濃度)的豐富度,明顯影響河流大型底棲動物群落結構[18]。水文因素方面,生活在激流中的大型底棲動物形體上具有激流適應性,可確保不被水流沖走[19]。生物因素方面,水生植物的豐富度與底棲動物現存量成正比[20],魚類捕食會減少底棲動物的生物量[21]。人類活動中,引水式水電站在某種程度上影響了河道中的物質輸送和能量流動,進而影響到大型底棲動物攝食類群的組成[3]。
本研究CCA分析結果顯示,環境變量NO-3N、TP是影響漢豐湖底棲動物密度的主要因素。水體氮、磷營養濃度的增加,有助于水域生態系統中生產者的生長、繁殖,進而為水域生態系統中的消費者——底棲動物各類功能攝食類群(刮食者、撕食者、收集者、捕食者)提供更豐富的食物來源,促進底棲動物的生長、繁殖,最終影響底棲動物的群落結構、物種多樣性。相關研究表明,底棲動物對水環境變化敏感,當水體受到污染時,其群落結構、生物多樣性都會發生改變[1-4]。隨著水體從中營養型向超富營養型過渡,大型底棲動物的物種多樣性下降[22];在水體富營養化日趨嚴重的情況下,底棲動物耐污染的優勢種類相當穩定,個體數量明顯增加[23]。漢豐湖位于重慶開縣縣城旁,有東河、南河兩條入湖支流,東河流域面積達1 469 km2,南河流域面積達1 710 km2,流域內人口眾多、鄉鎮密布,面源污水和城鎮生活廢水大量流入,豐富了漢豐湖水體營養物質,2013年湖水TN為1.584 mg/L、TP為0.083 mg/L、硝氮0.912 mg/L,TN達到地表水Ⅴ類、TP達到Ⅳ類標準,豐富的營養物質影響了漢豐湖底棲動物的群落結構。類似情況在其他水體也存在,如漢江中下游[24]、香溪河[3]、洪澤湖[25]水體中的NO-3N,洞庭湖[12]、松花江下游[26]水體中的TP,是影響其底棲動物群落結構的主要因子。
本研究CCA分析結果顯示,水深也是影響漢豐湖底棲動物密度的主要因素。一般來說,湖泊、水庫底棲動物密度和生物多樣性隨水深增加而不斷遞減[1-2]。因為隨著水深的增加,底棲動物所需的食物被微生物礦化的程度越大,底棲動物可利用的部分越少,因此底棲動物的現存量越低[27]。漢豐湖雖然不屬于深水湖泊,但是漢豐湖調節壩還未運行,水位受三峽水庫運行調度的影響,波動明顯,夏季水位最低,冬季水位最高,相差十多米,水位的波動對漢豐湖底棲動物物種豐富度和生物多樣性均有顯著影響[6]。類似情況在其他水體也存在,如天津于橋水庫[28]、武漢東湖[1]的水深是影響其底棲動物群落結構的主要因子。
按照國家防總批復的《三峽工程2014年試驗性蓄水實施計劃》,2015年5月25日漢豐湖水位調節壩開始蓄水試運行。隨著漢豐湖下閘運行,漢豐湖水位受三峽水庫運行的影響降低,水位將趨于穩定,同時漢豐湖水域生態系統隨著時間的推移,亦會慢慢發育至穩定,底棲動物群落結構組成將以寡毛類和搖蚊類為主,寡毛類為主要優勢類群[6]。此時,水深可能不會成為影響漢豐湖底棲動物密度的主要因素,但水體中的營養元素氮、磷等可能仍會像其他水體一樣,如漢江中下游[24]、香溪河[3]、洪澤湖[25]、洞庭湖[12],是影響漢豐湖底棲動物密度的主要因素。至漢豐湖水位穩定時,南河的HF1和東河的HF5兩個采樣點的生境可能仍會處于河流、湖泊相互交替狀態,底棲動物的群落結構受河流生態和湖泊生態的影響,河流態類型、湖泊態類型和河湖過渡態類型的底棲動物群落共存,此時上游徑流的流速、流量以及水深變化等因素可能成為影響HF1、HF5兩個采樣點底棲動物密度的主要因素。漢豐湖水位穩定后,沿岸沉水植物、挺水植物將會越來越豐富,豐富的水生植物可為底棲動物的繁殖、攝食、捕食、生長以及避難提供優良生境[27],不同生活習性的底棲動物亦會越來越豐富,此時沿岸帶水生植物將成為影響沿岸帶底棲動物分布的主要因素。
參考文獻(References):
[1] 劉健康. 高級水生物學[M]. 北京: 科學出版社, 1999.
[2] 段學花, 王兆印, 徐夢珍. 底棲動物與河流生態評價[M]. 北京: 清華大學出版社, 2010.
[3] 渠曉東. 香溪河大型底棲動物時空動態、生物完整性及小水電站的影響研究[D]. 武漢: 中國科學院水生生物研究所, 2006.
[4] 王強. 山地河流生境對河流生物多樣性的影響研究[D]. 重慶: 重慶大學, 2011.
[5] 李波, 袁興中, 熊森, 等. 城市消落帶景觀基塘系統設計初探——以重慶開縣漢豐湖為例[J]. 重慶師范大學學報(自然科學版), 2013, 31(6): 51-54.
[6] 彭建華, 遲仕運, 鄭志偉, 等. 漢豐湖水位調節壩運行前水位變動對底棲動物群落結構的影響分析[J]. 環境影響評價, 2014(6): 44-49, 58.
[7] TerBraak C J F, Prentic I C. A theory of gradient analysis[J]. ADVANCES IN ECOLOGICAL RESEARCH, 1986, 18:271-317.
[8] 國家環境保護總局, 《水和廢水監測分析方法》編委會. 水和廢水監測分析方法[M]. 4版. 北京: 中國環境科學出版社, 2002.
[9] LEPS J, SMILAUER P. Multivariate analysis of ecological data using CANOCO[M]. New York: Cambridge University Press, 2003: 13-15, 50-51, 70-217.
[10] Gazer M H. Vegetation composition and floristical diversity in date palm orchards of Central Saudi Arabia[J]. Acta Botanica Hungarica, 2011, 53(12): 111-126.
[11] Bilkovic D M, Roggero M, Hershner C H, et al. Influence of land use on macrobenthic communities in nearshore estuarine habitats[J]. Estuarines and coasts, 2006, 29(6): 1185-1195.
[12] 汪星, 鄭丙輝, 劉錄三, 等. 洞庭湖典型斷面底棲動物組成及其與環境因子的相關分析[J]. 中國環境科學, 2012, 32(12): 2237-2244.
[13] Thienemann, A. Ein drittes biozonotisches Grundprinzip[J]. Arch. Hydrobiol., 1954, 49(3): 421-422.
[14] Vannote R L, Minshall G W, Cummins KW, et al. The river continuum concept[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1980, 37: 130-137.
[15] Tait C K, Li J L, Lamberti G A, et al. Relationships between riparian cover and the community structure of high desert stream[J]. Journal of the North American Benthological Society, 1994, 13(1): 45-56.
[16] Stout J, Vandermeer J. Comparison of species richness for streaminhabiting insects in tropical and midlatitude streams[J]. American Naturalist, 1975, 109(967): 263-280.
[17] Scheuhammer A M, Mcnicol D K, Mallory M L, et al. Relationships between lake chemistry and calcium and trace metal concentrations of aquatic invertebrates eaten by breeding insectivorous waterfowl[J]. Environmental Pollution, 1997, 96(2): 235-247.
[18] Mason C F. Biology of freshwater pollution[M]. 4th ed. Harlow: PrenticeHall, 2002.
[19] Hershey A E, Lamberti G A. Stream macroinvertebrate communities[M]//Naiman R J, Bilby R E. River ecology and management. New York: Springer New York Inc. 2001.
[20] Soszka G J. Ecological relations between invertebrates and submerged macrophytes in the lake littoral[J]. Ekol Polska, 1975, 23: 393-415.
[21] Kajak Z. Considerations on benthos abundance in freshwaters, its factors and mechanisms[J]. International Revue der Gesmten Hydrobiologie Und Hydrographie, 1988, 73(1): 5-19.
[22] Gong Zhijun, Xie Ping. Impact of Eutrophication on Biodiversity of the macrozoobenthos community in a Chinese shallow lake[J]. J. of Freshwater Ecology, 2001, 16(2): 171-178.
[23] 王士達. 武漢東湖底棲動物的多樣性及其與富營養化的關系[J]. 水生生物學報, 1996, 20(增刊): 75-89.
[24] 潘曉潔, 朱愛民, 鄭志偉, 等. 漢江中下游春季浮游植物群落結構特征及其影響因素. 生態學雜志, 2014, 33(1): 33-40.
[25] 張超文, 張堂林, 朱挺兵, 等. 洪澤湖大型底棲動物群落結構及其與環境因子的關系[J].水生態學雜志, 2012, 33(3): 27-33.
[26] 李中宇, 胡顯安, 劉錄三. 松花江下游底棲動物組成及其環境指示作用[J]. 中國環境監測, 2014, 30(1): 86-92.
[27] 王銀東, 熊邦喜, 陳才保, 等. 環境因子對底棲動物生命活動的影響[J]. 浙江海洋學院學報(自然科學版), 2005, 24(3): 253-257.
[28] 馬秀娟. 天津于橋水庫大型底棲動物群落結構研究[D]. 武漢: 華中農業大學, 2012.
猜你喜歡
商情(2016年42期)2016-12-23 14:25:52
商情(2016年42期)2016-12-23 13:35:35
東方教育(2016年4期)2016-12-14 22:15:13
財經界·學術版(2016年19期)2016-11-16 16:28:33
科技視界(2016年21期)2016-10-17 17:37:34
中國實用醫藥(2016年24期)2016-10-17 04:31:12
中國實用醫藥(2016年24期)2016-10-17 03:37:40
中國實用醫藥(2016年24期)2016-10-17 03:35:06
科學與財富(2016年28期)2016-10-14 21:58:50
