解淀粉芽孢桿菌抑菌機制的研究進展
2015-10-27 01:25:05陳哲黃靜趙佳王長彪梁宏
生物技術通報 2015年6期
關鍵詞:研究
陳哲 黃靜 趙佳 王長彪 梁宏
(山西省農業科學院生物技術研究中心,太原 030031)
解淀粉芽孢桿菌抑菌機制的研究進展
陳哲 黃靜 趙佳 王長彪 梁宏
(山西省農業科學院生物技術研究中心,太原 030031)
解淀粉芽孢桿菌能夠產生種類繁多的抑菌物質,可以有效的抑制真菌和細菌的活性。近幾年來,關于解淀粉芽孢桿菌抑菌機制的研究報道越來越多,大部分研究已經深入到分子水平。綜述了國內外有關解淀粉芽孢桿菌的全基因組信息和解淀粉芽孢桿菌抑菌機制的相關研究。
解淀粉芽孢桿菌;基因組;抑菌機制
目前,在生物防治菌中人們研究較多的是芽孢桿菌。芽孢桿菌是土壤和自然界的優勢微生物種群,廣泛存在于自然界中,大多為非致病性細菌,對人畜無害,并且很多種類都具有抑制植物病害生長的能力。芽胞桿菌大多為內生芽胞,繁殖速度快,抗逆能力強,易定殖在植物根際表面。芽孢細菌種類繁多,廣泛應用于生物防治的種類有枯草芽胞桿菌、蠟狀芽胞桿菌、地衣芽胞桿菌及解淀粉芽孢桿菌[1]。目前關于生防菌的基礎研究集中在抑菌物質的種類、產生條件和分離純化等方面,隨著部分菌株全基因組測序的完成,研究已經逐步深入到分子水平,研究者開始關注抑菌物質產生的相關基因簇以及研究單個基因或者某幾個關聯基因對于抑菌物質分泌的影響等。已經有研究成功的通過Genome shuffling得到能大量分泌抑菌物質的突變菌株,改造后的突變菌株F2-38,surfactin的搖瓶產量比對照提高了3.5倍,在發酵罐中產量提高了10.3倍,而熒光定量(fluorescent quantitation,FQ)RT-PCR分析證實,菌株F2-38的surfactin合成酶基因srfA的表達量時原始菌株的15.7倍[2]。本文主要對國內外解淀粉芽孢桿菌的抑菌基因和抑菌機制的研究進展進行簡要綜述,對于今后工作的開展有重要意義。
1 解淀粉芽孢桿菌的全基因序列
1.1 解淀粉芽孢桿菌的基因組信息預測
從2007年解淀粉芽孢桿菌FZB42基因組序列的公布開始,已經有大約20多株不同的解淀粉芽孢桿菌進行了全基因組測序。
作為首個被測序的菌株,對解淀粉芽孢桿菌FZB42基因組序列的研究最為深入,其主要特征在表1中列出,同時還與枯草芽孢桿菌基因組進行了比較。解淀粉芽孢桿菌FZB42可以編碼3 693個基因,與芽孢桿菌屬的其他成員相比,其中有310個基因是其特有的[3,4]。
2013年,解淀粉芽孢桿菌UCMB5036完成測序,其基因組大小是3 910 324 bp,G+C的比例為46.60%,預測有3 660個編碼序列(CDSs),89個tRNA基因序列,10個 rRNA操作子,預測包含3 842個預測的開放閱讀框(ORF),其中95.39%的序列與菌株FZB42基因組序列具有同源性,這表明這兩個菌株之間有高度的基因同線性(gene synteny)[5]。
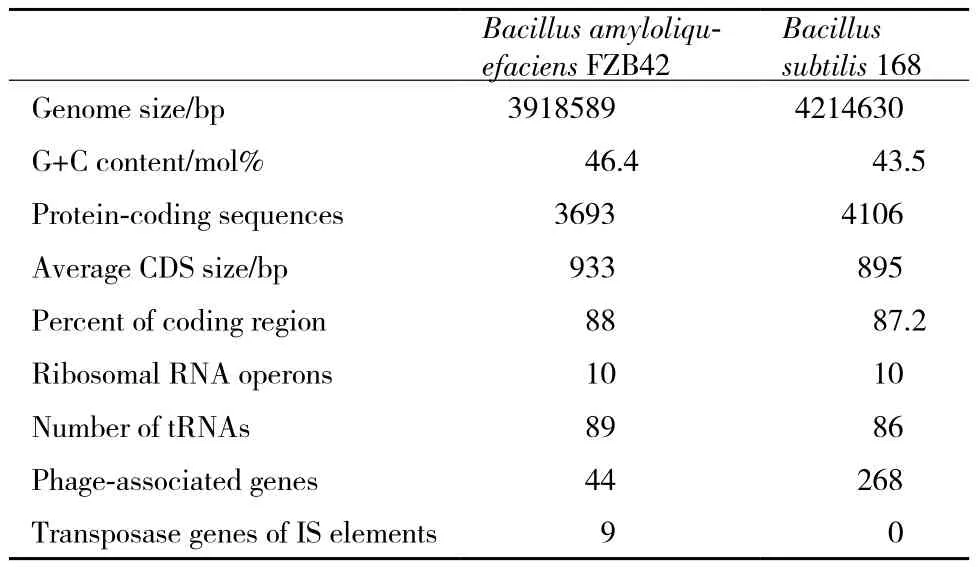
表1 解淀粉芽孢桿菌FZB42基因組與枯草芽孢桿菌的主要特征比較
1.2 抑菌物質的相關基因簇
根據解淀粉芽孢桿菌FZB42的全基因組序列信息,我們發現解淀粉芽孢桿菌可以產生種類繁多的次級代謝產物,這些產物可以抑制植物根際中的有害細菌和真菌。在解淀粉芽孢桿菌的基因序列中有9個巨大基因簇可以指導合成生物活性肽和聚酮化合物,二者分別在非核糖體肽合成酶(NRPS)和聚酮合酶(PKS)的組織下合成(表2)。非核糖體肽合酶(NRPS)是由許多功能模塊組成的復雜酶合成體系[6],模塊的數量、種類和排列順序決定了最終產物具有多樣性的結構、功能和作用機制。聚酮化合物的生物合成途徑遵循和脂肽合成途徑一樣的規則[7]。
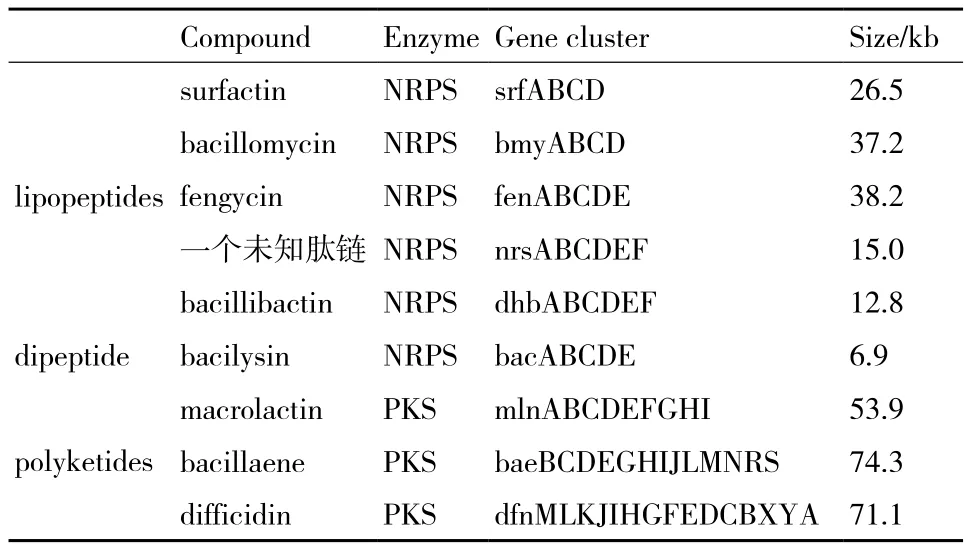
表2 參與非核糖體合成途徑的基因簇
在解淀粉芽孢桿菌FZB42中,能夠合成次級代謝產物的基因總長為340 kb,約占總遺傳力的8.5%,是枯草芽孢桿菌的兩倍[8]。而且在枯草芽孢桿菌中沒有發現負責合成脂肽bacillomycin D,聚酮化合物difficidin 和macrolactin以及一種未知肽鏈的相關基因簇[7]。
同樣的,解淀粉芽孢桿菌UCMB5036的基因組序列中也包含了非核糖體肽合成酶(non-ribosomal peptide synthetase,NRPS)和聚酮合酶(polyketide synthase,PKS)基因簇:surfactin、fengycin、difficidin、bacilysin、macrolactin、bacillaene、bacillomycin D 和bacillibactin,這些基因簇負責菌株次級代謝產物的產生。此外,菌株UCMB5036基因組中還包含其他功能的基因:分解代謝的植物來源的化合物、耐重金屬和藥物、運動和趨化、根定植,以及其他可以使細菌具有植物共生優勢的功能[5]。
1.3 核糖體合成途經的相關基因
與脂肽和聚酮化合物相比,蛋白質類及酶類的抑菌物質合成所涉及的基因就相對簡單許多,并且合成過程也很明確,遵循一般蛋白質的核糖體合成過程。值得注意的是,在枯草芽孢桿菌中羊毛硫抗生素lantibiotics(mersacidin和 subtilin)是常見的由核糖體合成的細菌素,但是解淀粉芽孢桿菌FZB42卻不能合成這種物質,基因組中沒有相應的編碼基因,但是卻有阻礙其合成的基因簇[4]。
2 抑菌機制的研究
目前,關于抑菌物質的抑菌機理有一定的研究,但還是沒有形成完整的體系。一般來說,抑菌物質可能在會在3個方面起作用:對病原菌的生長起到抑制作用;誘導植株的體內基因和免疫系統發生變化;抑菌物質使得拮抗菌具有一定特性,通過菌體的運動定殖在植株根部,發揮抑制作用。
2.1 對病原菌的作用
抑菌物質對病原菌的抑制主要體現在對其菌絲生長的抑制,以及對孢子萌發的影響這兩方面。
解淀粉芽孢桿菌YN- 1能夠抑制棉花枯萎病菌,用菌株發酵液的粗提物處理病原菌,結果表明,YN -1菌株粗提液能夠造成該病原菌的菌絲消融、變細,菌絲膜發生破裂,原生質體凝集滲漏,菌絲體扭曲變形,形成泡囊結構,孢子的產生受到嚴重抑制[9]。
利用高效液相色譜HPLC分析發現,解淀粉芽孢桿菌SWB16能夠分泌脂肽類抑菌物質芬枯草菌素和伊枯草菌素。試驗證明,這兩種物質可以抑制球孢白僵菌生長:一方面對孢子發芽及生長具有明顯的抑制及破壞作用,絕大多數分生孢子裂解,出現內容物外溢現象;另一方面,菌絲前部生長端出現膨大現象,菌絲體出現多處出現瘤狀畸形,菌絲生長受到明顯抑制[10]。
通過試驗發現,用純脂肽芬枯草菌素fengycins處理串珠鐮刀菌后,菌絲體內出現較大的囊泡,部分菌絲頂端破裂,菌絲膜也會受到破壞。但是,fengycins不容易破壞串珠鐮刀菌孢子的膜透性。此外,還發現卵磷脂很可能是fengycins在膜上的作用靶標,并且,fengycins能夠抑制串珠鐮刀菌分泌的磷脂酶A2的活性,該性質很可能也在fengycins的抑菌活性中起到了一定作用[11]。
2.2 對植株的作用
當菌株在植物根際定殖或大量繁殖時,抑菌物質可能會促使植物體內發生改變。例如,誘導免疫系統中的酶發生變化,或者系統抗病性發生變化。
有研究者對拮抗細菌 C-02 防治棉花黃萎病的機理展開了研究。結果表明,菌株C-02處理棉苗后,棉葉中植株防御性酶系中的苯丙氨酸解氨酶PAL、過氧化物酶POD 和多酚氧化酶PPO的活性提高幅度大,維持時間長,提高了棉苗系統防御能力,降低了病原菌侵染的可能性[12]。
誘導性系統抗性(ISR)是由寄生在植物根部的有益菌,或者稱為植物生長促進菌誘導植物產生的、對逆境和病害的系統抗性。研究表明,當受到溫度或者水分脅迫時,植物的誘導性系統抗病性(ISR)水平會下降;如果事先用解淀粉芽孢菌株S499對植株進行處理,就能夠降低這種負面影響。分析證實,在低溫情況下,解淀粉芽孢桿菌S499菌株的surfactin產量會顯著提高,增加菌株在植株根部的定殖能力,從而使得植物誘導性系統抗病性ISR的水平能維持平衡。許多試驗表明,surfactin與菌株在植物根際的活動,包括定殖能力、叢集運動以及生物膜的形成有關得以實現[13]。
2.3 抑菌物質自身的抑制作用
解淀粉芽孢桿菌分泌的抑菌物質有很多種,常見的種類分為脂肽類物質、聚酮類物質還有具有蛋白活性的抑菌物質。它們可以通過影響菌體自身的叢集運動、定殖能力、生物膜形成能力,促使菌體在植物根際大量繁殖或者定殖,進而起到保護作用。目前研究較多的是脂肽類的抑菌物質,解淀粉芽孢桿菌SQR9可以合成bacillomycin D和fengycin兩種脂肽,對香蕉枯萎病菌有顯著的拮抗作用。根據菌株SQR9分合的兩種物質,研究構建了兩種突變體:bacillomycin D 合成能力受到阻礙的突變體SQR9M1,對尖孢鐮刀菌有輕微的抑制作用;fengycin合成能力受到阻礙的突變體SQR9M2,其抑菌能力等同于野生型菌株SQR9。此外,通過在根部的原位實驗以及定量RT-PCR實驗證實,bacillomycin D有助于生物膜的形成[14]。
解淀粉芽孢桿菌FZB42的自發GFP熒光的突變體FB01mut可以用來研究菌株在不同物種中的定殖情況。在玉米中,突變體FB01mut主要定殖在初生根外表細胞的表面以及初生根和側根的交界處;而突變體FB01mut在擬南芥定植定植時,優先定殖于根尖;對于浮萍,突變體FB01mut通過形成生物膜定殖在葉子和根部,從定殖菌的浮萍植株上只提取到了表面活性素surfactin[15]。
還有研究者采用綠色熒光蛋白標記解淀粉芽孢桿菌FZB42發現,該細菌定植在不同物種的植物的根的表面表現不同。為了闡明哪些基因是根定植的關鍵,實驗構建了一系列突變體,即abrB-、sigH-、sigD-、nrfA-、yusV、RBAM017410、sfp、pabB-、pznA-和degU。用共聚焦激光掃描顯微鏡進行掃描分析發現,野生型菌株大量定殖在根尖和側根的表面,而突變體定殖在根尖的能力受到了影響,并且大多數突變體不能在側根定殖。其中,sfp、pabB-、pznA-和 degU基因被敲除的突變體菌株影響了細菌形成生物膜的能力以及菌體的群集運動,從而影響其在根部定殖的能力,這與大多數研究結果一致。有趣的是,菌株在植物根部的定殖能力依賴于生物膜的形成能力及群集運動。例如,試驗中構建的6個突變體,abrB-、sigH-、sigD-、 nrfA-、yusV和RBAM017410,它們在根部的定殖能力顯著下降,但是卻沒有影響菌株生物膜的形成,而且突變株nrfA-和yusV雖然不能在根尖或者或側根定殖,但是卻可以定殖在細胞邊緣及根的表面[16]。
為了研究菌株和根系表面接觸的分子機制,采用cDNA為基礎的消減抑制雜交,在解淀粉芽孢桿菌B55中鑒定出了7個上調的差異表達DNA片段。進一步試驗表明,在解淀粉芽孢桿菌B55和水稻根部共培養1 h之后,ansB(天門冬氨酸氨-裂合酶)和odhA(2-酮戊二酸脫氫酶E1組分)基因的RNA表達上調;隨著菌株和植物互相作用的時間增長,odhA基因呈連續上調表達,而ansB則在2 h后發生下調表達,這表明與水稻根部接觸的早期,ansB的上調表達是瞬間。可以看出,在細菌與植物根際相互作用的初期,會受到植物根系分泌物和根際微生物表面接觸的影響,這一時期是根際細菌定殖根部的關鍵時期,同時也開始促進植物的生長[17]。
需要注意的是,盡管我們能夠將抑制機理大致分為以上3種情況,但是在大自然中,抑菌物質在病原菌、植株和菌體三者之間發揮抑制作用一定依賴一個復雜而精細的調控網絡,這個網絡是我們今后研究的主要任務,尤其是在分子水平上的抑菌原理還需要進行更透徹的研究,包括基因表達和蛋白質組學的研究等,逐漸構建出完整系統的抑菌調控體系,為抑菌物質更好的發揮作用奠定良好的分子基礎。
3 展望
到目前為止,解淀粉芽孢桿菌的抑菌作用研究已經廣泛開展,但是生物體是復雜多變的,仍然有很多東西是未知的,并且利用解淀粉芽孢桿菌進行生物防治的前景十分廣闊,所以要加強對抑菌物質的分子研究。未來,抑菌物質的分子研究應該集中在以下幾個方面:研究抑菌調控體系,明確抑菌物質在病原菌、植株和菌體三者之間發揮作用的機制;改良合成抑菌物質的相關基因,使菌株產生更多的抑菌物質,從而有更強的抑菌能力;找到抑制病原菌生長的關鍵基因,將其轉入到植物中,使得植物本身具有抵抗病原菌的能力;使用高表達的宿主菌進行抗菌物質的高效表達,開展抑菌物質生物制劑的研究,包括制劑的規模化生產、儲存條件、存活期限、施用方法以及施入土壤后的存活條件和繁衍系數等方面;還可以通改造已知抑菌物質的結構,人工合成抗菌物質,從而在植物真菌的生物防治中發揮不亞于天然抑菌物質的作用[18]。
[1] Benitez LB, Velho RV, Lisboa MP, et al. Isolation and characterization of antifungal peptides produced by Bacillus amyloliquefaciens LBM5006[J]. The Journal of Microbiology, 2010, 48(6):791-797.
[2] Zhao J, Li Y, Zhang C, et al. Genome shuffling of Bacillus amyloliquefaciens for improving antimicrobial lipopeptide production and an analysis of relative gene expression using FQ RT-PCR[J]. Journal of Industrial Microbiology & Biotechnology, 2012, 39(6):889-896.
[3] Chen X, Koumoutsi A, Scholz R, et al. Genome analysis of Bacillus amyloliquefaciens FZB42 reveals its potential for biocontrol of plant pathogens[J]. Journal of Biotechnology, 2009, 140(1):27-37.
[4] Chen XH, Koumoutsi A, Scholz R, et al. Comparative analysis of the complete genome sequence of the plant growth-promoting bacterium Bacillus amyloliquefaciens FZB42[J]. Nature Biotechnology,2007, 25(9):1007-1014.
[5] Manzoor S, Niazi A, Bejai S, et al. Genome sequence of a plant-Associated bacterium, Bacillus amyloliquefaciens strain UCMB5036[J]. Genome Announcements, 2013, 1(2):e0011113.
[6] Strieker M, Tanovi? A, Marahiel MA. Nonribosomal peptidesynthetases:structures and dynamics[J]. Current Opinion in Structural Biology, 2010, 20(2):234-240.
[7] Walsh CT. Polyketide and nonribosomal peptide antibiotics:modularity and versatility[J]. Science, 2004, 303(5665):1805-1810.
[8] Stein T. Bacillus subtilis antibiotics:structures, syntheses and specific functions[J]. Molecular Microbiology, 2005, 56(4):845-857.
[9] 鄧建良, 劉紅彥, 劉玉霞, 等. 解淀粉芽孢桿菌YN-1抑制植物病原真菌活性物質鑒定[J]. 植物病理學報, 2010, 40(2):202-209.
[10] 汪靜杰, 趙東洋, 劉永貴, 等. 解淀粉芽孢桿菌SWB16菌株脂肽類代謝產物對球孢白僵菌的拮抗作用[J]. 微生物學報,2014, 54(7):778-785.
[11] 胡梁斌, 周威, 章挺, 等. 抗菌肽 Fengycins 抑制串珠鐮刀菌的初步機制[J]. 微生物學通報, 2010, 37(2):251-255.
[12] 田紹仁, 聶太禮, 王夢亮, 等. 拮抗細菌C-02防治棉花黃萎病的機理研究[J]. 棉花學報, 2012, 24(5):420-426.
[13] Pertot I, Puopolo G, Hosni T, et al. Limited impact of abiotic stress on surfactin production in planta and on disease resistance induced by Bacillus amyloliquefaciens S499 in tomato and bean[J]. FEMS Microbiology Ecology, 2013, 86(3):505-519.
[14] Xu Z, Shao J, Li B, et al. Contribution of bacillomycin D in Bacillus amyloliquefaciens SQR9 to antifungal activity and biofilm formation[J]. Applied and Environmental Microbiology, 2013,79(3):808-815.
[15] Fan B, Chen XH, Budiharjo A, et al. Efficient colonization of plant roots by the plant growth promoting bacterium Bacillus amyloliquefaciens FZB42, engineered to express green fluorescent protein[J]. J Biotechnol, 2011, 151(4):303-311.
[16] Dietel K, Beator B, Budiharjo A, et al. Bacterial traits involved in colonization of Arabidopsis thaliana roots by Bacillus amyloliquefaciens FZB42[J]. The Plant Pathology Journal, 2013,29(1):59-66.
[17] Liu J, He D, Ma X, et al. Identification of up-regulated genes of Bacillus amyloliquefaciens B55 during the early stage of direct surface contact with rice R109 root[J]. Current Microbiology,2011, 62(1):267-272.
[18] 趙亦靜, 倪密, 諾林, 等. 人工合成抗菌肽對棉花黃萎病菌的抑菌效果[J]. 浙江大學學報:農業與生命科學版, 2013(1):11-17.
(責任編輯 狄艷紅)
Research Advances on Antibacterial Mechanism of Bacillus amyloliquefaciens
Chen Zhe Huang Jing Zhao Jia Wang Changbiao Liang Hong
(Biotechnology Research Center,Shanxi Academy of Agriculture Science,Taiyuan 030031)
Bacillus amyloliquefaciens produces a wide range of antibacterial substances which effectively inhibit fungal and bacterial activities. In recent years, more and more studies have focused on antibacterial mechanism of B. amyloliquefaciens, and most of them already at the molecular level. Hence, we review the research progress on the genome sequences and antibacterial mechanism of B. amyloliquefaciens in this study.
Bacillus amyloliquefaciens;genome;antibacterial mechanism
10.13560/j.cnki.biotech.bull.1985.2015.06.004
2014-09-16
山西省農業科學院產品項目(2012ycp18),山西省基礎研究計劃項目(2009021032-1),山西省農業科學院科技攻關項目(2013GG22)
陳哲,女,研究實習員,碩士,研究方向:拮抗菌抑菌物質;E-mail:changer0006@163.com
梁宏,助理研究員,研究方向:微生物;E-mail:lh1964@126.com
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
