激素協(xié)同作用對(duì)奶牛乳腺上皮細(xì)胞激素受體基因表達(dá)的影響
2015-12-25 01:52:04王洪榮
中國(guó)飼料 2015年13期
關(guān)鍵詞:胰島素
田 青 , 王洪榮
(1.江蘇食品藥品職業(yè)技術(shù)學(xué)院,江蘇淮安 223003;2.揚(yáng)州大學(xué),江蘇揚(yáng)州 225009)
激素受體是激素發(fā)揮其功能的主要途徑。Lee等(2011)發(fā)現(xiàn)胰島素(INS)要發(fā)揮其功能,必須先與α亞單位結(jié)合后,β亞單位中酪氨酸激酶才被激活,進(jìn)而使受體磷酸化,介導(dǎo)和調(diào)節(jié)細(xì)胞內(nèi)酶系統(tǒng)活性,控制物質(zhì)代謝。李真和李慶章(2010)發(fā)現(xiàn)在奶牛乳腺組織中,胰島素及其受體的變化是有規(guī)律的,INS濃度從妊娠期開(kāi)始上升至分娩前達(dá)到最大,而后開(kāi)始下降;胰島素受體(INSR)的表達(dá)量從青春期到泌乳初期逐漸升高,到泌乳高峰期持續(xù)高水平,隨后開(kāi)始下降;催乳素也必須先與特定組織(如脾臟、淋巴結(jié)、胸腺、乳腺、卵巢等)催乳素(RPLR)基因結(jié)合才能進(jìn)入細(xì)胞內(nèi)發(fā)揮其功能(王亮,2013)。研究表明,在奶牛乳腺內(nèi),催乳素首先與其受體結(jié)合,然后激活JAK2激酶,并通過(guò)JAK-STAT信號(hào)通路刺激乳腺腺泡發(fā)育,并在乳腺組織中刺激乳腺酪蛋白基因的表達(dá)進(jìn)而促進(jìn)乳的生成與分泌、發(fā)動(dòng)和維持泌乳(Akers,2006)。在體內(nèi),糖皮質(zhì)激素與其胞內(nèi)核受體結(jié)合后才能啟動(dòng)并調(diào)控相關(guān)基因的表達(dá)和乳蛋白的合成,并且通過(guò)基因組機(jī)制實(shí)現(xiàn)其生理藥理功能(李敏,2010)。
大量研究表明,激素與激素之間或激素與生長(zhǎng)因子之間均存在一定的協(xié)同作用,如催乳素(PRL)能增加胰島素促進(jìn)乳汁分泌和改善乳品質(zhì)的作用效果,同時(shí)添加蛋白質(zhì)前體物氨基酸尤其是必需氨基酸能顯著提高泌乳奶牛的產(chǎn)奶性能等。也有研究表明,INS、PRL和氫化可的松(HYD)對(duì)奶牛乳腺上皮細(xì)胞生長(zhǎng)、相關(guān)基因表達(dá)及酪蛋白相對(duì)含量的影響均存在著最佳使用劑量,即PRL 50 ng/mL、INS 50 ng/mL 和 HYD 1 ng/mL,且三者協(xié)同作用能促進(jìn)奶牛乳腺上皮細(xì)胞增殖,抑制其凋亡,從而為乳腺的泌乳奠定良好的生理基礎(chǔ)(田青和王洪榮,2014、2013)。由此可以推測(cè),三種激素的協(xié)同作用對(duì)其受體有一定的影響作用。因此,本試驗(yàn)以本試驗(yàn)室前期凍存的奶牛乳腺上皮細(xì)胞為對(duì)象,采用L9(34)正交試驗(yàn)設(shè)計(jì),研究了INS、PRL和HYD的協(xié)同作用對(duì)奶牛乳腺上皮細(xì)胞激素受體基因表達(dá)的影響,旨在得到有利于促進(jìn)奶牛乳腺上皮細(xì)胞三種激素受體基因表達(dá)的最佳濃度組合,為后期的研究奠定基礎(chǔ)。
1 材料與方法
1.1 試驗(yàn)材料 細(xì)胞:揚(yáng)州大學(xué)草食動(dòng)物營(yíng)養(yǎng)實(shí)驗(yàn)室凍存的4代奶牛乳腺上皮細(xì)胞。
主要試劑:DMEM/F12 (Gibco)、 胎牛血清(FBS,Gibco)、催乳素(Sigma)、胰島素-轉(zhuǎn)鐵蛋白-硒(Gibco)、表皮生長(zhǎng)因子(Sigma)、氫化可的松(Sigma)、胰島素(Sigma)、胰蛋白酶(Gibco)、DMSO(Gibco)、兩性霉素 B(Amresco)、PBS(北京索萊寶)、熒光定量檢測(cè)試劑盒(TaKaRa,寶生物)。
儀器與設(shè)備:電熱恒溫CO2培養(yǎng)箱 (美國(guó)Thermo)、倒置顯微鏡(CKX41,Olympus)、冷凍離心機(jī) (5415R 型, 德國(guó) Eppendorf)、PCR 儀(ABI 2720型)、NanoDrop ND-1000濃度測(cè)定儀 (美國(guó)Thermo)、 熒光定量 PCR 儀 (ABI 7500 型)、i-MarkTM 酶標(biāo)儀(168-1130,美國(guó) Bio-Rad)、移液器(德國(guó) Eppendorf)。
1.2 試驗(yàn)設(shè)計(jì) 細(xì)胞復(fù)蘇后等密度接種于6孔板,待細(xì)胞長(zhǎng)滿(mǎn)70%~80%時(shí),先用無(wú)血清、無(wú)激素但含雙抗、兩性霉素的DMEM/F12培養(yǎng)基過(guò)渡16 h以上,后采用L9(34)正交試驗(yàn)設(shè)計(jì),即分別用含有不同濃度激素組合的生長(zhǎng)培養(yǎng)基于37℃、5%CO2培養(yǎng)箱培養(yǎng)24 h,每個(gè)處理設(shè)3個(gè)重復(fù),每個(gè)試驗(yàn)獨(dú)立重復(fù)3次,具體方案見(jiàn)表1。
1.3 RNA的提取、cDNA的合成與real-time PCR 總RNA的提取:生長(zhǎng)培養(yǎng)基培養(yǎng)24 h后收獲細(xì)胞, 用 RNAiso Plus (TaKaRa,D9108A)總RNA提取試劑盒提取RNA,所有RNA樣品用紫外分光光度計(jì) (NANODROP-1000)測(cè)定其 260/280 nm吸收比率值,用以檢測(cè)RNA濃度、純度和完整性。提取的RNA保存于-70℃,待用。
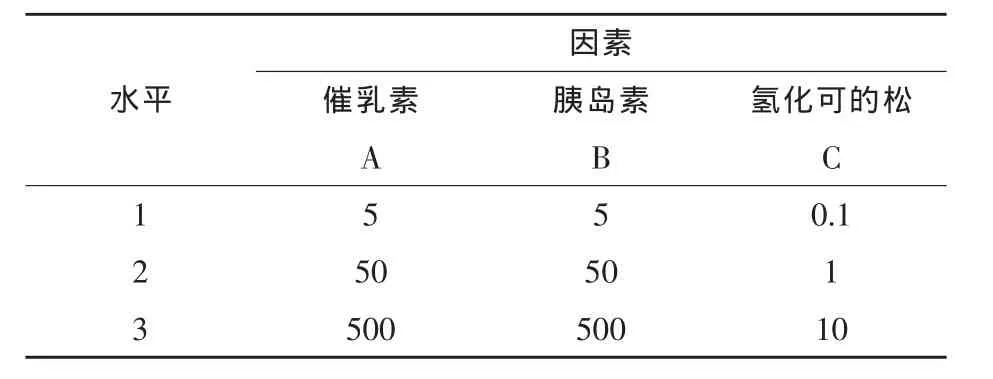
表1 正交試驗(yàn)因素與水平 ng/mL
反轉(zhuǎn)錄:以提取的RNA為模板,用Prime-Script? RT Master Mix(TaKaRa,DRR036A)反轉(zhuǎn)錄試劑盒制備cDNA,其總反應(yīng)體系為80 μL。轉(zhuǎn)錄得到的RT-PCR反應(yīng)液于4℃保存。
real-time PCR:反轉(zhuǎn)錄得到的RT反應(yīng)液用SYBR? Premix Ex TaqTM II(TaKaRa,DRR081A)試劑盒進(jìn)行real-time PCR,采用20 μL體系。反應(yīng)條件為 95 ℃、30 s;95 ℃、5 s,60 ℃、34 s,40 個(gè)循環(huán);溶解曲線(xiàn) 95 ℃、15 s,60 ℃、1 min,95 ℃、15 s,60 ℃、15 s。
各基因的相對(duì)表達(dá)量用2-ΔΔCT值表示。
1.4 奶牛乳腺上皮細(xì)胞中激素受體相關(guān)基因表達(dá)的檢測(cè) 以GAPDH為內(nèi)參,采用Primer3軟件進(jìn)行引物設(shè)計(jì),引物信息見(jiàn)表2。
1.5 數(shù)據(jù)統(tǒng)計(jì)與分析 所有原始數(shù)據(jù)先用Excel進(jìn)行數(shù)據(jù)統(tǒng)計(jì),用SAS 9.1中ANOVA進(jìn)行顯著性分析,Tukey法進(jìn)行多重比較,P<0.05表示差異顯著,結(jié)果用“平均值±標(biāo)準(zhǔn)差”表示。
2 結(jié)果與分析
表3結(jié)果顯示,各試驗(yàn)組三種激素受體基因表達(dá)量與對(duì)照組相比差異均不顯著(P>0.05),各試驗(yàn)組之間三種激素受體基因表達(dá)量差異也不顯著(P>0.05),但各試驗(yàn)組三種激素受體基因表達(dá)量均高于對(duì)照。
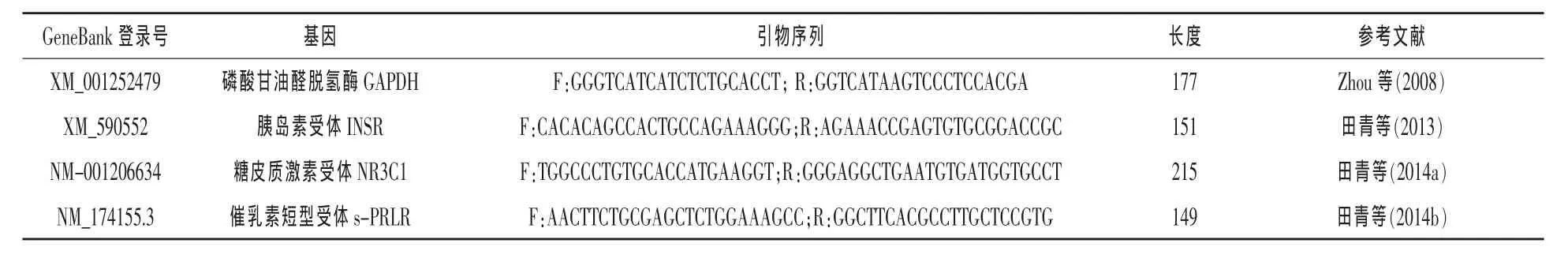
表2 引物信息
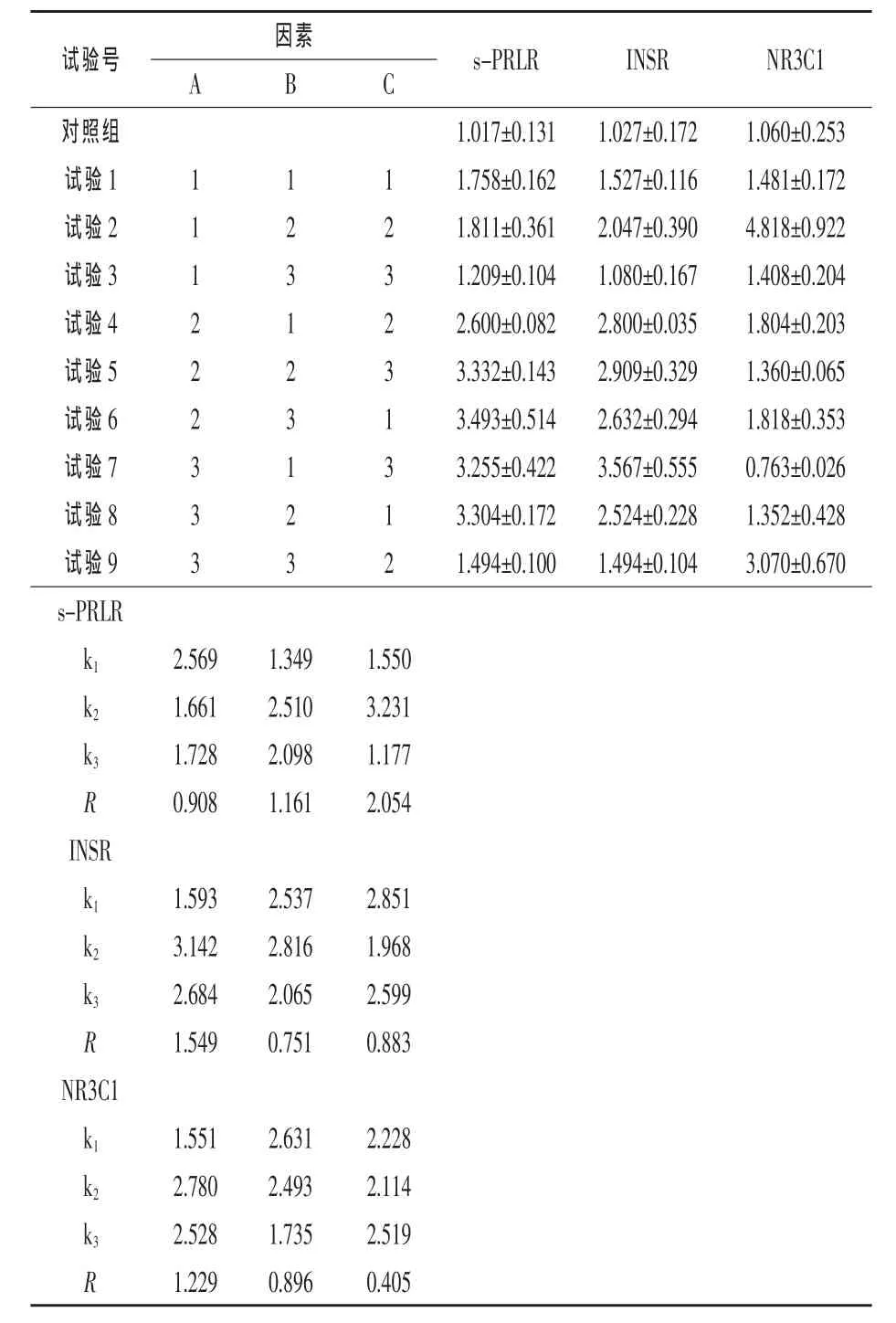
表3 激素協(xié)同作用對(duì)奶牛乳腺上皮細(xì)胞激素受體基因表達(dá)的影響
從三種激素協(xié)同作用對(duì)奶牛乳腺上皮細(xì)胞影響的最優(yōu)組合和影響程度來(lái)看,對(duì)s-PRLR基因,三種激素的最優(yōu)組合為A1B2C2(PRL 5 ng/mL、INS 50 ng/mL、HYD 1 ng/mL),三種激素對(duì)試驗(yàn)結(jié)果的影響程度為C>B>A;對(duì)NR3C1基因,三種激素的最優(yōu)組合為 A2B1C3(PRL 50 ng/mL、INS 5 ng/mL、HYD 10 ng/mL),三種激素對(duì)試驗(yàn)結(jié)果的影響程度為A>B>C;對(duì)INSR基因而言,三種激素的最優(yōu)組合為 A2B2C1(PRL 50 ng/mL、INS 50 ng/mL、HYD 0.1 ng/mL),三種激素對(duì)試驗(yàn)結(jié)果的影響程度為A>C>B。
3 討論
通過(guò)對(duì)試驗(yàn)結(jié)果的分析可以看出,受體基因不同,三種激素的最優(yōu)組合和影響程度也各不相同,試驗(yàn)結(jié)果并未表現(xiàn)出明顯的規(guī)律,也沒(méi)有對(duì)三種激素受體均一致的最優(yōu)組合。但可發(fā)現(xiàn),三種激素的組合中某種激素含量高,對(duì)應(yīng)的該種激素受體的表達(dá)量就高 (氫化可的松除外,NR3C1的表達(dá)量隨著氫化可的松的增加而降低),由此可知,對(duì)于泌乳期的奶牛,催乳素的需要量適當(dāng)大一些,胰島素次之,氫化可的松的用量最小。
研究認(rèn)為胰島素通過(guò)與受體結(jié)合,主要通過(guò)mTOR信號(hào)通路發(fā)揮作用,即胰島素激活I(lǐng)RS-1,IRS-1的磷酸化可活化信號(hào)分子PI3K,活化后的PI3K催化其下游靶蛋白Akt磷酸化,磷酸化的Akt抑制TSC2的活性,在TSC1的參與下,mTOR被激活,進(jìn)而發(fā)揮調(diào)節(jié)糖代謝的功能 (Biona和Loor,2011;Sophie 等,2007);對(duì)于催乳素,比較公認(rèn)的則是催乳素與受體結(jié)合后激活JAK2/STAT5信號(hào)通路,也可激活Ras/Raf/MAPK及ERK和PI3K/AKT信號(hào)通路,進(jìn)而促進(jìn)奶牛乳腺上皮細(xì)胞的增殖,維持泌乳和促進(jìn)乳蛋白的合成(Swaminathan 等,2008;Wagner等,2004;);關(guān)于氫化可的松參與奶牛乳蛋白合成調(diào)控的研究并不多見(jiàn),但有研究表明糖皮質(zhì)激素通過(guò)調(diào)控E6-P53-miR145信號(hào)通路誘導(dǎo)宮頸癌細(xì)胞產(chǎn)生耐藥表型 (杜利彬,2011),也有研究表明氫化可的松發(fā)揮作用主要是在機(jī)體處于能量負(fù)平衡等特殊狀態(tài)下促進(jìn)乳腺周?chē)鸂I(yíng)養(yǎng)物質(zhì)的重分配而維持產(chǎn)奶量和乳蛋白含量的,由此可以推測(cè),動(dòng)物處于不同生理?xiàng)l件下,氫化可的松的作用信號(hào)通路不同,糖皮質(zhì)激素發(fā)揮的作用也不同。在奶牛的乳腺中,前期研究發(fā)現(xiàn)單獨(dú)添加氫化可的松的確具有增加奶牛乳腺上皮細(xì)胞合成乳蛋白能力的作用(田青和王洪榮,2014)。
從功能上來(lái)看,催乳素的主要功能是啟動(dòng)和維持泌乳,胰島素則主要與能量代謝有關(guān),而氫化可的松主要與炎癥反應(yīng)(免疫)有關(guān)。催乳素可以通過(guò)與其受體結(jié)合先為泌乳的啟動(dòng)和維持奠定生理基礎(chǔ)(充分發(fā)育的乳腺),胰島素可以通過(guò)與其受體結(jié)合為乳腺上皮細(xì)胞的生長(zhǎng)、泌乳的啟動(dòng)和維持提供能量保證(充分的能量),而氫化可的松則可以通過(guò)與其受體結(jié)合而維持乳腺細(xì)胞組織健康。在體外培養(yǎng)的條件下,奶牛乳腺上皮細(xì)胞要增殖和生長(zhǎng)就需要較多的催乳素和胰島素參與,所以催乳素和胰島素受體的表達(dá)量也相對(duì)較高;而體外培養(yǎng)主要是在無(wú)菌條件下培養(yǎng)得到的具有正常形態(tài)和生物學(xué)功能的健康乳腺上皮細(xì)胞,所以不需要更多的氫化可的松發(fā)揮其抗炎癥功能,因此對(duì)應(yīng)的氫化可的松受體基因表達(dá)量就較低。
4 結(jié)論
4.1 催乳素、胰島素和氫化可的松協(xié)同作用有利于促進(jìn)PRLR、NR3C1、INSR基因受體的表達(dá)。
4.2 催乳素、胰島素和氫化可的松協(xié)同促進(jìn)s-PRLR、NR3C1和INSR基因表達(dá)的最優(yōu)組合分別為催乳素5 ng/mL、胰島素50 ng/mL、氫化可的松1 ng/mL,催乳素50 ng/mL、胰島素5 ng/mL、氫化可的松10 ng/mL和催乳素50 ng/mL、胰島素50 ng/mL、氫化可的松0.1 ng/mL;對(duì)s-PRLR、NR3C1和INSR基因表達(dá)影響的大小次序分別為:氫化可的松>胰島素>催乳素;催乳素>胰島素>氫化可的松;催乳素>氫化可的松>胰島素。
[1]杜利彬.糖皮質(zhì)激素通過(guò)調(diào)控E6-P53-miR145信號(hào)通路影響宮頸癌耐藥:[碩士學(xué)位論文][D].開(kāi)封:河南大學(xué),2011.
[2]李敏.新型糖皮質(zhì)激素對(duì)肥大細(xì)胞脫顆粒的快速作用及其機(jī)制研究[博士學(xué)位論文][D].上海:上海第二軍醫(yī)大學(xué),2010.
[3]李真,李慶章.奶山羊乳腺發(fā)育過(guò)程中生長(zhǎng)激素、胰島素及其受體的變化規(guī)律研究[J].中國(guó)農(nóng)業(yè)科學(xué),2010,43(8):1730 ~ 1737.
[4]田青,王洪榮.氫化可的松對(duì)奶牛乳腺上皮細(xì)胞生長(zhǎng)及酪蛋白合成的影響[J].畜牧獸醫(yī)學(xué)報(bào),2014a,45(10):1663 ~ 1670.
[5]田青,王洪榮.胰島素、催乳素和氫化可的松對(duì)奶牛乳腺上皮細(xì)胞增殖和凋亡的影響[J].中國(guó)飼料,2013,2:8 ~ 12.
[6]田青,王洪榮.催乳素對(duì)奶牛乳腺上皮細(xì)胞生長(zhǎng)及酪蛋白合成的影響[J].中國(guó)畜牧雜志,2014b,50(19):52~57.
[7]王海燕.子宮內(nèi)膜催乳素功能及信號(hào)轉(zhuǎn)導(dǎo)機(jī)制[J].國(guó)外醫(yī)學(xué):婦產(chǎn)科學(xué)分冊(cè),2003,30(6):340 ~ 343.
[8]王亮.葛根素對(duì)小鼠生長(zhǎng)激素和催乳素含量及其受體表達(dá)量的影響:[碩士學(xué)位論文][D].呼和浩特:內(nèi)蒙古農(nóng)業(yè)大學(xué),2013.
[9]Akers R M.Major advances associated with hormone and growth factor regulation of mammary growth and lactation in dairy cows[J].Journal of Dairy Science,2006,89:1222 ~ 1234.
[10]Biona M,Loor J J.Gene networks driving bovine mammary protein systhesis during the lactation cycle[J].Bioininformatics and Biology Insights,2011,5:83 ~ 98.
[11]Lee J Y,Park D W,Kim Y H,et al.Incidence,predictors,treatment,and longterm prognosis of patients with restenosis after drug-eluting stent implantation for unprotected left main coronary artery disease[J].J Am Coll Cardio,2011,57(12):1349 ~ 1358.
[12]Sophie T,Sonia M,Sophie D,et al.Regulation of protein metabolism by insulin:Value of different approaches and animal models[J].Domestic Animal Endocrinology,2007,33:123 ~ 142.
[13]Swaminathan G,Varghese B,F(xiàn)uchs S Y.Regulation of prolactin receptor levers and activity in breast cancer[J].The journal of Mammary Gland Biology Neoplasia,2008,13(1):81 ~ 91.
[14]Wagner K U,Krempler A,Triplett A A,et al.Impaired alveologenesis and maintenance of secretory mammary epithelial cells in JAK2 conditional knockout mice[J].Molecular and Cellular Biology,2004, 24:5510 ~ 5520.
[15]Zhou Y,Akers R M,Jiang H.Growth hormone can induce expression of four major milk protein genes in transfected MAC-T cells[J].Journal of Dairy Science,2008,91:100 ~ 108.
猜你喜歡
人人健康(2023年26期)2023-12-07 03:55:46
家庭醫(yī)藥(2019年9期)2019-09-23 18:54:32
中國(guó)生殖健康(2019年2期)2019-08-23 08:12:10
家庭科學(xué)·新健康(2018年8期)2018-10-30 10:23:20
人生與伴侶·共同關(guān)注(2018年5期)2018-08-15 10:00:00
科學(xué)生活(2016年9期)2016-10-20 13:12:45
中國(guó)衛(wèi)生標(biāo)準(zhǔn)管理(2015年1期)2016-01-14 03:41:27
人人健康(2015年17期)2015-09-09 16:25:20
藥學(xué)與臨床研究(2015年4期)2015-06-05 11:35:51
中國(guó)醫(yī)藥科學(xué)(2015年15期)2015-02-27 12:32:27
