氣道上皮細(xì)胞衍生的胰島素樣生長因子觸發(fā)偏移CD8+T細(xì)胞極化
2016-01-12 02:05:54黃邵洪張軍航
中國藥理學(xué)通報 2015年2期
關(guān)鍵詞:凋亡
黃邵洪,李 昀,覃 杰,安 軍,張軍航,榮 健
(中山大學(xué)附屬第三醫(yī)院1.胸心外科、2.放射科,廣東 廣州 510630;3.中山大學(xué)附屬第一醫(yī)院麻醉科,廣東,廣州 510089)
氣道上皮細(xì)胞衍生的胰島素樣生長因子觸發(fā)偏移CD8+T細(xì)胞極化
黃邵洪1,李昀1,覃杰2,安軍1,張軍航1,榮健3
(中山大學(xué)附屬第三醫(yī)院1.胸心外科、2.放射科,廣東 廣州510630;3.中山大學(xué)附屬第一醫(yī)院麻醉科,廣東,廣州510089)
中國圖書分類號:R322.35;R329.25;R392.12;R977.6
摘要:目的探討氣道上皮細(xì)胞衍生的胰島素樣生長因子(IGF1)對CD8+T細(xì)胞極化的影響。方法人氣道上皮細(xì)胞株RPMI2650與鼠過敏原Der p1共培養(yǎng)72 h,用定量PCR及Western免疫印跡檢測其IGF1表達(dá)情況。將前述上皮細(xì)胞、重組IGF1和IGF1抗體分別加入抗體激活的CD8+T細(xì)胞中培養(yǎng),用流式細(xì)胞儀檢測細(xì)胞凋亡。檢測Der p1活化的上皮細(xì)胞及IGF1對CD8+T細(xì)胞p53基因甲基化及表達(dá)的影響。結(jié)果加入Der p1共同孵育后,RPMI2650細(xì)胞IGF1mRNA(23.1%±5.2% vs 5.2%±2.3%,P<0.01)和蛋白表達(dá)(33.4±6.4 vs 9.2±4.6,P<0.01)均明顯增加。將CD3/CD28抗體激活的CD8+T細(xì)胞與Der p1活化的上皮細(xì)胞共培養(yǎng),凋亡細(xì)胞增加的趨勢被抑制(41.7%±8.2% vs 5.2%±1.8%,P<0.01)。直接加入重組IGF1有同樣效果,而IGF1抗體能阻斷該效應(yīng)。Der p1活化的上皮細(xì)胞能抑制活化CD8+T細(xì)胞p53基因mRNA(29.1%±5.9% vs 16.2%±4.3%,P<0.01)和蛋白表達(dá)(63.3±8.9 vs 26.9±5.6,P<0.01),加入IGF1抗體后這種作用消失。重組IGF1使CD8+T細(xì)胞p53基因甲基化增加。結(jié)論Der p1蛋白能夠誘發(fā)RPMI2650細(xì)胞產(chǎn)生IGF1,后者通過誘導(dǎo)p53基因甲基化抑制CD8+T細(xì)胞凋亡。
關(guān)鍵詞:氣道上皮細(xì)胞;胰島素樣生長因子;p53基因;甲基化;CD8+T細(xì)胞;凋亡
偏移的CD8+T細(xì)胞極化在許多免疫異常反應(yīng)中起重要作用,比如風(fēng)濕性關(guān)節(jié)炎[1]、糖尿病[2]和顱內(nèi)感染[3]。在哮喘發(fā)生中,CD8+T細(xì)胞極化同樣起重要作用,但何種原因?qū)е缕錁O化并不清楚[4]。
胰島素樣生長因子(insulin-like growth factor-1,IGF1)又稱為生長調(diào)節(jié)素C(somatomedin C),促進生長和防止細(xì)胞死亡。IGF1在人的一生中都發(fā)揮作用,尤其是在兒童期和青春期。IGF1主要由肝臟產(chǎn)生,通常是生長激素啟動的后續(xù)信號。IGF1也是一些過敏性疾病(如哮喘)的發(fā)病因素之一。嗜堿性粒細(xì)胞表達(dá)IGF1受體,當(dāng)暴露于IGF1后,前者被激活并釋放胞內(nèi)顆粒[5]。小鼠氣道過敏模型中檢出IGF1高表達(dá)[6],還有學(xué)者認(rèn)為IGF1對未成熟的肺成纖維細(xì)胞有促進α平滑肌表達(dá)和膠原蛋白合成的作用,這將導(dǎo)致肺纖維化(由核因子κB轉(zhuǎn)錄介導(dǎo))[7]。除肝臟外,IGF1還可由氣道炎癥細(xì)胞產(chǎn)生,但具體是何種類型細(xì)胞尚不清楚[8]。
被激活后T細(xì)胞通過凋亡進入激活誘導(dǎo)的細(xì)胞死亡(activation-induced cell death, AICD),維持機體免疫平衡。作為死亡信號受體的Fas基因及其配體FasL,在T細(xì)胞被激活后表達(dá)上調(diào),F(xiàn)as/FasL相互作用下啟動細(xì)胞凋亡[9]。除了Fas/FasL通路外,還有其他分子在凋亡啟動中發(fā)揮作用,如p53蛋白。盡管p53蛋白主要是誘導(dǎo)腫瘤細(xì)胞凋亡,其他細(xì)胞中也有表達(dá),氮芥誘導(dǎo)的小鼠皮膚細(xì)胞凋亡和炎癥反應(yīng),這一過程就是依賴p53基因表達(dá)的[10]。紫外線照射誘導(dǎo)人角質(zhì)細(xì)胞調(diào)節(jié)ERK1/2和血栓調(diào)節(jié)素表達(dá)就是由p53基因介導(dǎo)的[11]。然而,p53基因在CD8+T細(xì)胞AICD過程,尤其是偏移CD8+T細(xì)胞極化中的作用尚不清楚。我們發(fā)現(xiàn),過敏性刺激使小鼠氣道上皮細(xì)胞表達(dá)IGF1,這種上皮細(xì)胞產(chǎn)生的IGF1抑制CD8+T細(xì)胞p53表達(dá),參與抗原特異性的CD8+T細(xì)胞極化過程。
1材料與方法
1.1試劑IGF1及p53抗體購自Santa Cruz Biotech上海分公司;定量qRT-PCR及Western免疫印跡試劑購自Invitrogen上海分公司;Der p1蛋白購自MyBiosource公司(San Diego, CA);顆粒酶B及穿孔素ELISA套件購自R&D Systems上海分公司;膜聯(lián)蛋白Annexin V套件購自Sigma Aldrich上海分公司;免疫細(xì)胞分離提取套件購自Miltenyi Biotech上海分公司;DNA純化套件購自Promega北京分公司;EpiXploreTM甲基化檢測套件購自Clontech北京分公司。
1.2細(xì)胞培養(yǎng)人鼻腔上皮細(xì)胞株RPMI2650購自美國細(xì)胞培養(yǎng)協(xié)會(American Type Culture Collection,ATCC, Manassas, VA, USA),置于DMEM細(xì)胞培養(yǎng)基(Dulbecco′s Modified Eagle Medium)培養(yǎng),加入10%小牛血清、100 kU·L-1青霉素、100 mg·L-1鏈霉素及200 mmol·L-1谷氨酸。培養(yǎng)箱條件為37℃恒溫,含5% CO2。
1.3定量實時PCR(qRT-PCR)細(xì)胞總RNA通過TRIzol reagents試劑盒提取。cDNA由逆轉(zhuǎn)錄試劑盒合成。qRT-PCR由MiniOpticon實時PCR系統(tǒng)(Bio-Rad Life Science公司, 上海)完成。研究所用引物為:IGF1 (NCBI: NM_001111283), 正向:3′-TCTGAATCTTGGCTGCTGGA-5′,反向: 3′-TGTGCTTCTTGACGACTTGC-5′;p53 (NCBI: AB082923), 正向:3′-TGGCCATCTACAAGCAGTCA-5′,反向: 3′-GGTACAGTCAGAGCCAACCT-5′。結(jié)果用內(nèi)參β-actin相對量表達(dá)。
1.4Western免疫印跡細(xì)胞總蛋白提取后由SDS-PAGE凝膠電泳分離,再轉(zhuǎn)印至硝酸纖維膜上。經(jīng)5%脫脂牛奶孵育封閉后加入一抗(100~500 μg·L-1),室溫下孵育1 h后,加入辣根過氧化物酶結(jié)合二抗繼續(xù)孵育1 h。每次孵育完成后,用緩沖液(Triton X-100)沖洗多余抗體。免疫反應(yīng)結(jié)束后生成化學(xué)發(fā)光底物,并由X射線膠片成像。膠片成像后由系統(tǒng)自帶圖像軟件量化免疫印跡密度。
1.5流式細(xì)胞計數(shù)細(xì)胞經(jīng)熒光標(biāo)記的一抗染色后由流式細(xì)胞計數(shù)儀(FACSCanto II,BD Bioscience,上海分公司)計數(shù)。凋亡細(xì)胞通過Annexin V套件染色后檢測。死亡細(xì)胞由碘化丙啶(propidium iodide,PI)5 mg·L-1, 15 min染色后檢測計數(shù)。
1.6CD8+T細(xì)胞純化由健康志愿者獲得外周血標(biāo)本(每人20 mL),實驗及血樣采集經(jīng)我院倫理委員會批準(zhǔn)。經(jīng)梯度離心獲得外周血單核細(xì)胞(peripheral blood mononuclear cells,PBMC)。通過免疫磁珠細(xì)胞分離套件將PBMC中CD8+T細(xì)胞及CD11c+樹突狀細(xì)胞(dendritic cells,DC)分離,并經(jīng)流式細(xì)胞儀檢測,CD8+CD25-T細(xì)胞及DC純度均超過98%。
1.7抗原特異性CD8+T細(xì)胞多克隆增殖的評估將CD8+CD25-T細(xì)胞用琥珀酰亞胺酯(carboxy fluorescein succinimidyl ester,CFSE)標(biāo)記,置于2 mg·L-1CD3抗體和5 mg·L-1CD28抗體包被環(huán)境下培養(yǎng)72 h。T細(xì)胞增殖率通過CFSE稀釋法測定。
1.8抗原特異性CD8+T細(xì)胞激活的評估將CD8+CD25-T細(xì)胞置于Transwell底層小室中培養(yǎng),間隔層為RPMI2650細(xì)胞株。Der p1蛋白(鹽水作為對照或加入抗IGF1中和抗體200 μg·L-1)置于頂層小室,濃度為30 μg·L-1,孵育72 h。收獲CD8+T細(xì)胞并用CFSE標(biāo)記,加入DC細(xì)胞(T細(xì)胞 ∶DC=5 ∶1),在30 μg·L-1Der p1蛋白或小牛血清蛋白中培養(yǎng)72 h。CD8+T細(xì)胞增殖率通過CFSE稀釋法測定。培養(yǎng)液中顆粒酶B及穿孔素水平由相應(yīng)ELISA試劑盒檢測。
1.951鉻(51Cr)釋放檢測激活的CD8+T細(xì)胞用51Cr釋放檢測法分析。以結(jié)腸癌細(xì)胞株T84作為靶細(xì)胞,用51Cr標(biāo)記孵育2 h。沖洗后將T84細(xì)胞(108·L-1)和CD8+T細(xì)胞(108·L-1)置于Transwell底層小室中。間隔層為RPMI2650細(xì)胞(109·L-1)。底層小室上清液每4 h收集一次75 μL。51Cr活性由γ射線計數(shù)器(Perkin Elmer公司,美國)測定,最大釋放量和自發(fā)釋放量分別通過加入2% Triton X-100和培養(yǎng)液獲得。CD8+T細(xì)胞激活產(chǎn)生的51Cr特異性釋放量由以下公式計算:(A-B)/(C-B)×100%,A:實際測定量,B:自發(fā)釋放量,C:最大釋放量。
1.10p53基因甲基化測定將純化的DNA用EpiXploreTM甲基化檢測套件進行亞硫酸鹽轉(zhuǎn)化后,進行甲基化測定。p53基因的引物序列分別為:(1)甲基化引物,正向:3′-ATTTTAGTGGTAATTTATTGGGACG-5′,反向: 3′-TAATAAAACTCCCCTTTCTTACGAA-5′;(2)非甲基化引物,正向:3′-TTTTAGTGGTAATTTATTGGGATGG-5′,反向: 3′-TAATAAAACTCCCCTTTCTTACAAA-5′。
2結(jié)果
2.1Der p1蛋白誘導(dǎo)氣道上皮細(xì)胞生成IGF1氣道上皮細(xì)胞能夠產(chǎn)生IGF1[12],但其觸發(fā)生成的誘因尚未明確。我們同時檢測了IGF1在人氣道上皮細(xì)胞(RPMI2650細(xì)胞)基線水平的mRNA(Fig 1A)和蛋白(Fig 1B-C)水平的表達(dá)量,二者在暴露于Der p1后均明顯增加,并且這種增加具有劑量依賴性(Fig 1)。該結(jié)果提示:鼠致敏原能上調(diào)人氣道上皮細(xì)胞IGF1表達(dá)。
Human airway epithelial cell line RPMI2650 cells, was cultured in the presence or absence of a mite allergen, Der p1, for 72 h. A: The bars indicate the levels of IGF1 mRNA; B: The immune blots indicate the IGF1 protein levels. The bars below the blots indicate the summarized integrated density of the blots. BSA is an irrelevant control protein (50 μg·L-1); C: The bars indicate the cell counts from day 0 to day 3.**P<0.01vsBSA group (A, B), or day 0 group (C).
2.2氣道上皮細(xì)胞生成的IGF1抑制CD8+T細(xì)胞AICD盡管偏移CD8+T細(xì)胞極化在氣道過敏性疾病發(fā)生中起作用[13],但上皮細(xì)胞表達(dá)的IGF1是否對CD8+T細(xì)胞極化有作用尚不清楚。我們發(fā)現(xiàn),原始CD8+T細(xì)胞能夠被CD3/CD28抗體激活,表現(xiàn)為CD8+T細(xì)胞增殖(Fig 2A),同時激活的CD8+T細(xì)胞中存在大量凋亡細(xì)胞(Fig 2B-C)。在混合培養(yǎng)液中,鹽水對照組中凋亡CD8+T細(xì)胞處于低水平(Fig 2B、2Ca),加入CD3/CD28抗體激活后凋亡CD8+T細(xì)胞明顯增加(Fig 2B、2Cb)。加入Der p1孵育的上皮細(xì)胞后,凋亡細(xì)胞增加的趨勢被抑制(Fig 2B、2Cc),而加入未經(jīng)處理的上皮細(xì)胞則無此效果(Fig 2B、2Cd)。該結(jié)果提示,氣道上皮細(xì)胞生成的IGF1會抑制CD8+T細(xì)胞AICD過程。為進一步證實該想法,我們在上述實驗中加入IGF1抗體,發(fā)現(xiàn)Der p1孵育上皮細(xì)胞對CD8+T細(xì)胞凋亡抑制作用被阻斷了(Fig 2B、2Ce、2Cf),加入重組IGF1進一步證實了該結(jié)果(Fig 2B、2Cg)。
2.3氣道上皮細(xì)胞生成的IGF1通過MEK-ERK通路誘導(dǎo)CD8+T細(xì)胞p53基因甲基化p53基因產(chǎn)物能夠誘導(dǎo)細(xì)胞凋亡,由此我們進一步確定IGF1是否通過該途徑抑制CD8+T細(xì)胞AICD[14]。我們利用實驗Fig 2中部分細(xì)胞,檢測其生成的IGF1能否調(diào)節(jié)CD8+T細(xì)胞p53蛋白表達(dá)。定量PCR和Western免疫印跡均顯示CD8+T細(xì)胞p53蛋白表達(dá)。以CD3/CD28抗體激活后,CD8+T細(xì)胞p53蛋白表達(dá)明顯增加, Der p1孵育的RPMI2650細(xì)胞能抑制該趨勢,而加入IGF1抗體后這種作用消失(Fig 3A-B)。由此可見,激活CD8+T細(xì)胞將使p53蛋白表達(dá)增加,而上皮細(xì)胞生成的IGF1則可以阻斷該過程。
為闡明IGF1抑制CD8+T細(xì)胞p53表達(dá)的機制,我們進行了甲基化特異性PCR檢測。結(jié)果顯示,在原始CD8+T細(xì)胞中,p53基因存在部分甲基化,而在暴露于重組IGF1后,這一比例明顯增加。在MEK-ERK缺乏CD8+T細(xì)胞中則未觀察到該現(xiàn)象(Fig 3C)。Western免疫印跡顯示,IGF1處理后的CD8+T細(xì)胞Bcl-2和Bcl-xL蛋白水平并無明顯變化(Fig 3D-E)。
2.4Der p1蛋白誘導(dǎo)的氣道上皮細(xì)胞激活的CD8+T細(xì)胞具有Der p1特異性免疫反應(yīng)氣道上皮細(xì)胞表達(dá)MHC I和MHC II[15]及B7分子,并將抗原呈遞給T細(xì)胞[14]。為了明確氣道上皮細(xì)胞生成的IGF1能否作為抗原特異性偏移CD8+T細(xì)胞極化的啟動因子,我們將原始CD8+T細(xì)胞和Der p1蛋白誘導(dǎo)的氣道上皮細(xì)胞共同孵育48 h。將獲得的CD8+T細(xì)胞置于特異性抗原Der p1環(huán)境下,加入新鮮提取的DC共孵育72 h。特異性CD8+T細(xì)胞出現(xiàn)明顯增殖,加入IGF1中和抗體后增殖被抑制(Fig 4A)。
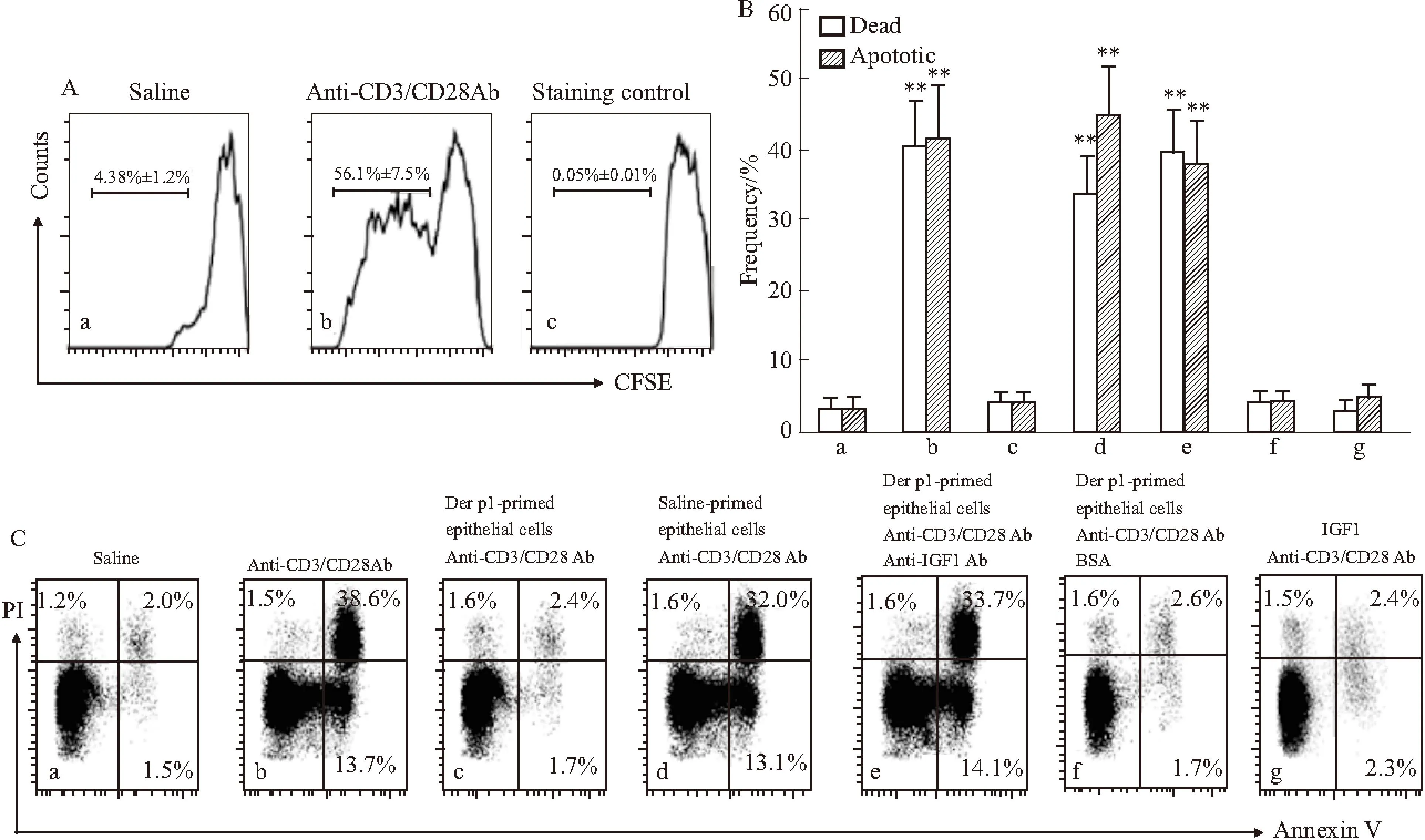
Fig 2 Epithelial cell-derived IGF1 reduces AICD of CD8+ T cells
A: The histograms indicate the proliferation of CD8+T cells; B: The bars indicate the frequency of dead CD8+T cells and apoptotic CD8+T cells in the activated CD8+T cells; C: The dot plots indicate the frequencies of PI+or/and Annexin V+CD8+T cells.**P<0.01vsgroup a.
Na?ve CD8+T cells were treated in the same procedures of Fig 2C, the bars (A) and immune blots (B) indicate the levels of p53 in CD8+T cells.**P<0.01vsgroup a. C:The p53 gene methylation results. M: Methylated; U: Un-methylated; rIGF1: Recombinant IGF1; U0126: The MEK-ERK inhibitor; LY294002:The PIK3-Akt inhibitor. BSA is an irrelevant protein using as a control; Anti-CD3/CD28: CD8+T cells were treated with anti-CD3/CD28 antibodies. D and E: The immune blots indicate the protein contents of Bcl-2 (D) and Bcl-xL (E) in the CD8+T cell extracts.
培養(yǎng)液中細(xì)胞毒因子、顆粒酶B(Fig 4B)及穿孔素(Fig 4C)水平明顯升高。該結(jié)果提示,氣道上皮細(xì)胞生成的IGF1能夠觸發(fā)抗原特異性CD8+T細(xì)胞極化。CD8+T細(xì)胞的激活通過特異性溶解反應(yīng)得到證實(Fig 4D)。
3討論
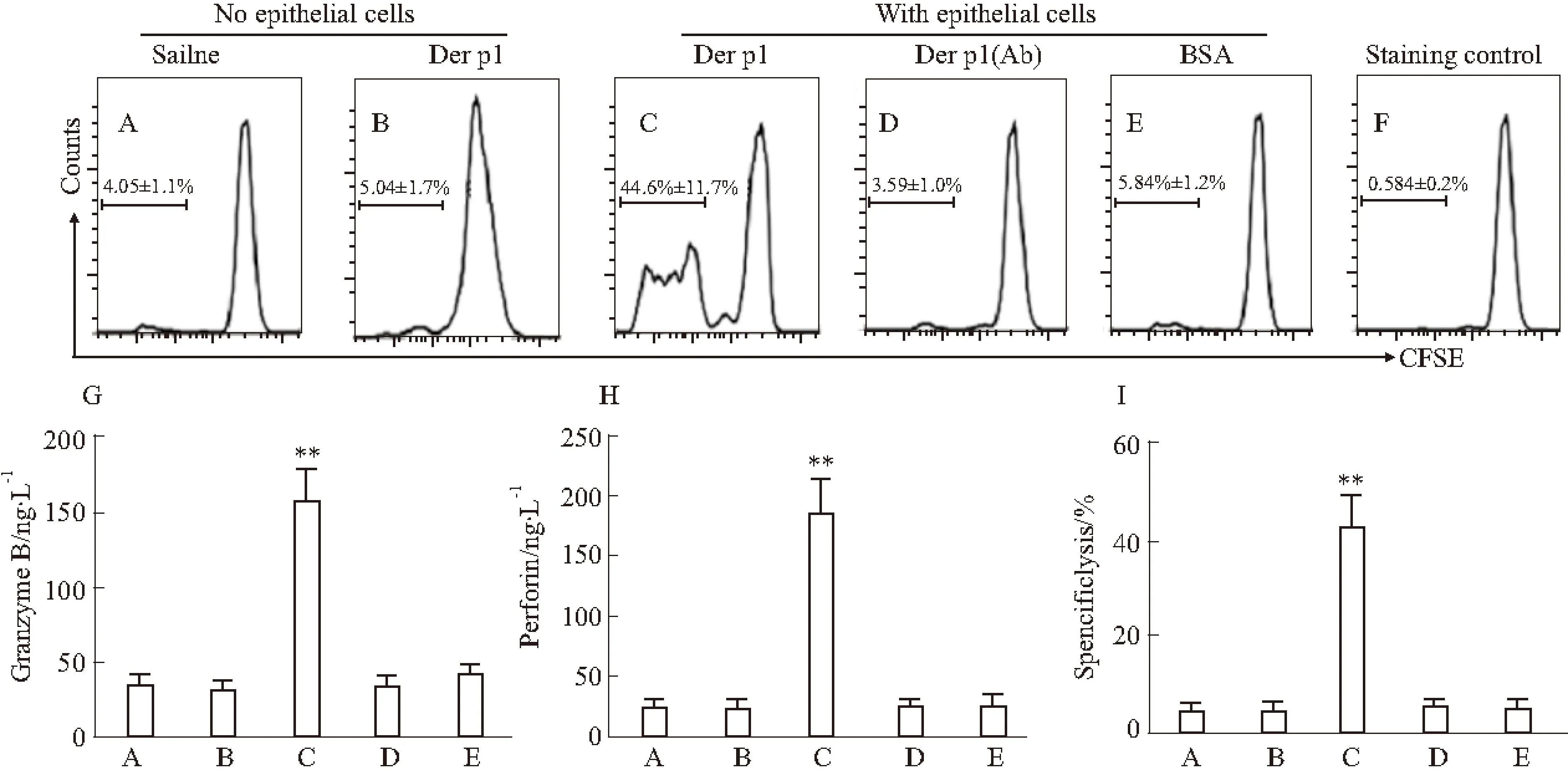
Fig 4 Epithelial cell-derived IGF1 induces CD8+ T cell polarization
A~F:The histograms indicate the frequency of CD8+T cell proliferation; A~B: Without the presence of epithelial cells; C~E: In the presence of epithelial cells; F: Staining controls; G~I: The bars indicate the levels of granzyme B (G), perforin (H) and specific lysis (I). The labels of x axis are the same as the labels of histograms.**P<0.01vsgroup A.
許多免疫系統(tǒng)異常或多或少都與偏移CD8+T細(xì)胞極化相關(guān)。本研究顯示氣道上皮細(xì)胞(RPMI2650細(xì)胞),能夠在鼠過敏原Der p1蛋白作用下產(chǎn)生IGF1。而這種氣道上皮細(xì)胞生成的IGF1將使p53基因發(fā)生甲基化,進而抑制CD8+T細(xì)胞的凋亡進程,促成CD8+T細(xì)胞極化。
鼠過敏原在許多過敏性疾病發(fā)生過程中扮演重要角色。除了作為通常意義下的過敏原外,它們中的一些亞型(如Der p1)具有蛋白酶樣作用,能夠影響上皮細(xì)胞緊密連接處的完整性,并且能夠激活蛋白酶活化受體2,從而破壞上皮屏障功能[16]。我們的研究提供了進一步證據(jù):Der p1能夠誘導(dǎo)RPMI2650細(xì)胞生成IGF1。IGF1通常是由肝細(xì)胞生成的,但其他細(xì)胞(如氣道組織中的炎癥細(xì)胞)也可以產(chǎn)生[8]。
AICD是一種T細(xì)胞的正常激活反應(yīng)。這種過程在T細(xì)胞被激活后就啟動并誘導(dǎo)凋亡,AICD在維持機體免疫系統(tǒng)穩(wěn)定中意義重大。AICD是由Fas受體(Fas,CD95)和Fas配體(FasL,CD95 ligand)相互作用下誘導(dǎo)的細(xì)胞凋亡。機體許多(免疫耐受)重要器官,如睪丸、大腦、角膜等,其細(xì)胞均有FasL高表達(dá),用以抑制免疫反應(yīng)對這些器官可能造成的損傷。這就不難理解如果AICD機制出現(xiàn)異常,會發(fā)生多種自身免疫疾病。本研究顯示,被CD3/CD28抗體激活后凋亡CD8+T細(xì)胞明顯增加,而氣道上皮細(xì)胞生成的IGF1使凋亡和死亡細(xì)胞數(shù)減少。加入IGF1中和抗體后,減少CD8+T細(xì)胞凋亡的作用被抑制,再加入重組IGF1又可恢復(fù)這種抑制作用。其他學(xué)者也發(fā)現(xiàn)IGF1能夠抑制細(xì)胞凋亡[17-18]。
p53基因作用缺失會造成許多疾病。p53蛋白是一種腫瘤抑制蛋白,在許多腫瘤中存在變異[19]。p53蛋白還在其他疾病中起作用,諸如缺血性細(xì)胞損傷[20]、吸煙相關(guān)性疾病[21]。我們發(fā)現(xiàn)p53蛋白在氣道上皮細(xì)胞生成的IGF1誘導(dǎo)抑制CD8+T細(xì)胞AICD過程中有作用。IGF1能夠通過MEK-ERK通路誘發(fā)p53基因甲基化。其他學(xué)者證實在間質(zhì)干細(xì)胞向成骨細(xì)胞轉(zhuǎn)化時,IGF1同樣通過MEK-ERK通路促進這一進程[22]。
總之,鼠過敏原Der p1蛋白能夠誘發(fā)氣道上皮細(xì)胞——RPMI2650細(xì)胞產(chǎn)生IGF1,后者通過誘導(dǎo)p53基因甲基化抑制CD8+T細(xì)胞AICD。
參考文獻:
[1]Carvalheiro H, da Silva J A,Souto-Carneiro M M. Potential roles for CD8(+) T cells in rheumatoid arthritis[J].AutoimmunRev, 2013, 12(3): 401-9.
[2]Coppieters K T,von Herrath M G. Viruses and cytotoxic T lymphocytes in type 1 diabetes[J].ClinRevAllergyImmunol, 2011, 41(2): 169-78.
[3]Willing A,Friese M A. CD8-mediated inflammatory central nervous system disorders[J].CurrOpinNeurol, 2012, 25(3): 316-21.
[4]Robinson D S. The role of the T cell in asthma[J].JAllergyClinImmunol, 2010, 126(6): 1081-91.
[5]Koketsu R, Suzukawa M, Kawakami A, et al. Activation of basophils by stem cell factor: comparison with insulin-like growth factor-I[J].JInvestigAllergolClinImmunol, 2008, 18(4): 293-9.
[6]Jaradat M, Stapleton C, Tilley S L, et al. Modulatory role for retinoid-related orphan receptor alpha in allergen-induced lung inflammation[J].AmJRespirCritCareMed, 2006, 174(12): 1299-309.
[7]Chetty A, Cao G J,Nielsen H C. Insulin-like growth factor-I signaling mechanisms, type I collagen and alpha smooth muscle actin in human fetal lung fibroblasts[J].PediatrRes, 2006, 60(4): 389-94.
[8]Vieira R P, Duarte A C, Claudino R C, et al. Creatine supplementation exacerbates allergic lung inflammation and airway remodeling in mice[J].AmJRespirCellMolBiol, 2007, 37(6): 660-7.
[9]Lettau M, Paulsen M, Schmidt H, et al. Insights into the molecular regulation of FasL (CD178) biology[J].EurJCellBiol, 2011, 90(6-7): 456-66.
[10]Inturi S, Tewari-Singh N, Jain A K, et al. Absence of a p53 allele delays nitrogen mustard-induced early apoptosis and inflammation of murine skin[J].Toxicology, 2013, 311(3): 184-90.
[11]Huang H C, Chang T M, Chang Y J, et al. UVB irradiation regulates ERK1/2- and p53-dependent thrombomodulin expression in human keratinocytes[J].PLoSOne, 2013, 8(7): e67632.
[12]Kawaguchi M, Fujita J, Kokubu F, et al. Induction of insulin-like growth factor-I by interleukin-17F in bronchial epithelial cells[J].ClinExpAllergy, 2010, 40(7): 1036-43.
[13]Dakhama A, Collins M L, Ohnishi H, et al. IL-13-producing BLT1-positive CD8 cells are increased in asthma and are associated with airway obstruction[J].Allergy, 2013, 68(5): 666-73.
[14]Pant V, Quintas-Cardama A,Lozano G. The p53 pathway in hematopoiesis: lessons from mouse models, implications for humans[J].Blood, 2012, 120(26): 5118-27.
[15]Kreisel D, Lai J, Richardson S B, et al. Polarized alloantigen presentation by airway epithelial cells contributes to direct CD8+T cell activation in the airway[J].AmJRespirCellMolBiol, 2011, 44(6): 749-54.
[16]Roelandt T, Heughebaert C,Hachem J P. Proteolytically active allergens cause barrier breakdown[J].JInvestDermatol, 2008, 128(8): 1878-80.
[17]Chung H, Seo S, Moon M, et al. IGF-I inhibition of apoptosis is associated with decreased expression of prostate apoptosis response-4[J].JEndocrinol, 2007, 194(1): 77-85.
[18]Garrouste F, Remacle-Bonnet M, Fauriat C, et al. Prevention of cytokine-induced apoptosis by insulin-like growth factor-I is independent of cell adhesion molecules in HT29-D4 colon carcinoma cells-evidence for a NF-kappaB-dependent survival mechanism[J].CellDeathDiffer, 2002, 9(7): 768-79.
[19]Muller P A,Vousden K H. p53 mutations in cancer[J].NatCellBiol, 2013, 15(1): 2-8.
[20]Hong L Z, Zhao X Y,Zhang H L. p53-mediated neuronal cell death in ischemic brain injury[J].NeurosciBull, 2010, 26(3): 232-40.
[21]Yu M W, Yang S Y, Chiu Y H, et al. A p53 genetic polymorphism as a modulator of hepatocellular carcinoma risk in relation to chronic liver disease, familial tendency, and cigarette smoking in hepatitis B carriers[J].Hepatology, 1999, 29(3): 697-702.
[22]Xue P, Wu X, Zhou L, et al. IGF1 promotes osteogenic differentiation of mesenchymal stem cells derived from rat bone marrow by increasing TAZ expression[J].BiochemBiophysResCommun, 2013, 433(2): 226-31.
網(wǎng)絡(luò)出版時間:2015-1-9 13:37網(wǎng)絡(luò)出版地址:http://www.cnki.net/kcms/doi/10.3969/j.issn.1001-1978.2015.02.012.html
Airway epithelial cell-derived insulin-like growth factor-1
triggers skewed CD8+T cell polarization
HUANG Shao-hong1, LI Yun1,QIN Jie2, AN Jun1, ZHANG Jun-hang1, RONG Jian3
(1.DeptofThoracocardiacSurgery, 2.DeptofRadiology,theThirdAffiliatedHospital,SunYet-senUniversity,Guangzhou
510630,China; 3.DeptofAnesthesiology,theFirstAffiliatedHospital,SunYet-senUniversity,Guangzhou510089,China)
Abstract:AimTo investigate the effects of airway epithelial cell-derived insulin-like growth factor-1(IGF1) on CD8+T cell polarization. MethodsHuman airway epithelial cell line, RPMI2650 cells, was cultured in the presence of a mice allergen, Der p1, for 72 h. IGF1 expression was checked with quantitative RT-PCR and Western blot. Der p1-primed RPMI2650 cells, recombinant IGF1 and anti-IGF1 antibody was cocultured respectively with CD8+T cells, which were activated by anti-CD3/CD8 Ab. Apoptotic cells frequency was calculated with flow cytometry. The alteration of p53 gene hypermethylation in CD8+T cells elicited by Der p1-primed airway epithelial cell and IGF1 was plotted. ResultsBoth mRNA(23.1%±5.2% vs 5.2%±2.3%,P<0.01)and protein(33.4±6.4 vs 9.2±4.6,P<0.01)expression of IGF1 in RPMI2650 cells markedly increased after exposure to Der p1. The increase of apoptotic CD3/CD28 Ab-activated CD8+T cells was abolished by the presence of Derp1-primed epithelial cells(41.7%±8.2% vs 5.2%±1.8%,P<0.01). The results were con-firmedbytheadditionofrecombinantIGF1.Anti-IGF1 antibody abolished the effect of the epithelial cells. Derp1-primed epithelial cells inhibited p53 gene mRNA(29.1%±5.9% vs 16.2%±4.3%,P<0.01)and protein(63.3±8.9 vs 26.9±5.6,P<0.01)expression. Anti-IGF1 antibody abolished the effect. Recombinant IGF1 promoted CD8+T cells′ p53 gene hypermethylation. ConclusionDer p1 induces RPMI2650 cells to produce IGF1, and this factor prevents CD8+T cell apoptosis by inducing p53 gene hypermethylation.
Key words:airway epithelial cell; insulin-like growth factor-1; p53 gene; hypermethylation; CD8+T cell; apoptosis
通訊作者榮健(1971-),女,博士,副教授,副主任醫(yī)師,研究方向:體外循環(huán)及惡性腫瘤臨床與基礎(chǔ),,E-mail:jiji126@126.com
作者簡介:黃邵洪(1976-),男,博士,主治醫(yī)師,研究方向:肺癌及食管腫瘤臨床與基礎(chǔ),E-mail:legendhuang@126.com;
基金項目:國家自然科學(xué)基金資助項目(No 81371714);廣東省自然科學(xué)基金資助項目(No S2012010008151)
收稿日期:2014-11-11,修回日期:2014-12-12
文獻標(biāo)志碼:A
文章編號:1001-1978(2015)02-0204-07
doi:10.3969/j.issn.1001-1978.2015.02.012
猜你喜歡
中國當(dāng)代醫(yī)藥(2016年31期)2017-02-23 17:40:52
中外醫(yī)學(xué)研究(2017年1期)2017-02-23 14:40:35
中國當(dāng)代醫(yī)藥(2016年31期)2017-02-23 13:22:56
中國當(dāng)代醫(yī)藥(2016年32期)2017-02-23 13:22:29
中國現(xiàn)代醫(yī)生(2016年27期)2016-12-21 16:05:30
中國醫(yī)藥導(dǎo)報(2016年25期)2016-11-30 07:20:06
中國實用醫(yī)藥(2016年14期)2016-05-20 10:25:43
中國實用醫(yī)藥(2016年7期)2016-03-17 14:04:02
科技視界(2016年5期)2016-02-22 19:03:28
中國現(xiàn)代醫(yī)生(2015年21期)2015-08-07 12:33:18
