
茄子果實(shí)性狀遺傳及其分子育種研究進(jìn)展
2016-10-19 22:05:06王倩朱士農(nóng)崔群香張愛慧王長義劉衛(wèi)東
湖北農(nóng)業(yè)科學(xué) 2016年5期
關(guān)鍵詞:研究
王倩 朱士農(nóng) 崔群香 張愛慧 王長義 劉衛(wèi)東
摘要:對茄子(Solanum melongena L.)果實(shí)性狀的遺傳規(guī)律等進(jìn)行了綜述,并重點(diǎn)闡述了與果實(shí)性狀相關(guān)的分子標(biāo)記、遺傳圖譜構(gòu)建及基因定位的研究進(jìn)展,對未來該領(lǐng)域的研究方向進(jìn)行了展望。
關(guān)鍵詞:茄子(Solanum melongena L.);果實(shí)性狀;遺傳;分子標(biāo)記;基因定位
中圖分類號:S641.1;S336 文獻(xiàn)標(biāo)識碼:A 文章編號:0439-8114(2016)05-1089-05
DOI:10.14088/j.cnki.issn0439-8114.2016.05.001
Research Progress on Inheritance and Molecular Breeding of Fruit Traits in Eggplant
WANG Qian,ZHU Shi-nong,CUI Qun-xiang,ZHANG Ai-hui,WANG Chang-yi,LIU Wei-dong
(Department of Horticulture, Jinling Institute of Technology, Nanjing 210038,China)
Abstract: In this review, inheritance affecting eggplant(Solanum melongena L.) fruit traits was summarized. Moreover, the paper focused on molecular markers, construction of genetic map and relative gene mapping on fruit traits of fresh eggplant. Lastly, the direction of future research on the field was prospected.
Key words: eggplant(Solanum melongena L.); fruit traits; inheritance; molecular markers; gene mapping
茄子(Solanum melongena L.)屬于茄科茄屬,是世界各地廣泛栽培的一種蔬菜作物,起源于亞洲南部熱帶地區(qū)。一般認(rèn)為中國是茄子的第二起源地及次生演化中心,于1 500年前傳入中國,擁有豐富的品種資源[1]。茄子不僅具有較高的營養(yǎng)價(jià)值,且食用方法也多種多樣,是一種可以鮮食加工結(jié)合、周年供應(yīng)、經(jīng)濟(jì)實(shí)惠的大宗蔬菜,深受廣大消費(fèi)者的歡迎。但與其他茄科蔬菜作物番茄、馬鈴薯、辣椒相比,茄子基礎(chǔ)理論方面的研究比較薄弱,近些年來這方面的研究開始受到重視。
茄子主要的形態(tài)性狀為葉、莖、花、果實(shí),其中果實(shí)性狀最為典型[2]。《中國蔬菜品種志》中茄子品種分類也主要依據(jù)果實(shí)形態(tài)將其分為圓茄、卵茄、長茄等類型。由于各地生態(tài)環(huán)境及消費(fèi)習(xí)慣不同,茄子栽培品種形成了不同的生態(tài)類型和市場消費(fèi)區(qū)域[3]。果實(shí)性狀是影響消費(fèi)者對茄子購買需求的主要因素之一,因此育種工作者都在積極利用新資源和新技術(shù)對茄子果實(shí)主要性狀的遺傳規(guī)律進(jìn)行研究與分析[4]。本文對茄子果實(shí)性狀的遺傳規(guī)律等進(jìn)行了綜述,重點(diǎn)闡述了茄子果實(shí)性狀的分子標(biāo)記以及相關(guān)性狀基因的定位研究現(xiàn)狀,并對茄子果實(shí)性狀以后的研究方向進(jìn)行了展望。
1 果實(shí)品質(zhì)性狀遺傳規(guī)律
1.1 果色及果萼色性狀
在茄子的育種改良中,茄子果實(shí)的顏色是一個重要的品質(zhì)性狀。花青素是構(gòu)成茄子果實(shí)的主要色素,屬類黃酮化合物。早在20世紀(jì)初國外便有學(xué)者對茄子花青素進(jìn)行了初步的遺傳研究,Tatebe[5,6]第一次提出控制茄子果實(shí)顏色形成的遺傳因子(基因)包括6個,分別為C、P、D、G、Gv和Puc,其中C、P和D是主效基因,調(diào)控花青素的形成,G和Gv為控制前期果實(shí)中葉綠素的形成因子,Puc調(diào)控位于花萼下部的色素形成。隨后Janick等[7]發(fā)現(xiàn)了一個與D緊密連鎖的互補(bǔ)基因,命名為X。Tigchelaar等[8]研究發(fā)現(xiàn)一個新的可抑制花青素合成的基因(Y),以及調(diào)控花藥形成花青素條紋的基因(Sa),因此Tigchelaar等[8]指出控制茄子花青素形成的基因共計(jì)9個。并通過配制雜交組合進(jìn)一步驗(yàn)證了不同基因的功能,例如D基因有4個等位基因(D、d、dt和dw),P基因有3個等位基因(P、p和pw),而Y基因只有兩個等位基因(Y和y)。這些基因的不同組合使得成熟茄子果實(shí)表現(xiàn)出不同顏色。
李錫香等[1]將茄子果色分為白、白綠、綠、橘紅、淺紫、鮮紫、紫紅和黑紫共計(jì)8類。劉進(jìn)生等[9]按照質(zhì)量性狀遺傳的方法對茄子果色進(jìn)行了研究,結(jié)果表明深紫色對非紫色為不完全顯性。育種實(shí)踐中果色遺傳表現(xiàn)出連續(xù)變異的特點(diǎn),呈現(xiàn)出許多中間型過渡色,應(yīng)為數(shù)量性狀。
近些年來中國學(xué)者通過多世代聯(lián)合的數(shù)量性狀分離分析法,對茄子果色遺傳規(guī)律進(jìn)行了研究。龐文龍等[10]以3個果色不同的茄子栽培種自交系為試驗(yàn)材料,通過P1、P2、F1、B1、B2和F2六世代聯(lián)合分析法,得出茄子果色遺傳為多基因控制的數(shù)量性狀,其遺傳符合2對加性-顯性-上位性主基因+加性-顯性-上位性多基因模型(E模型);主基因遺傳力為35.5%~98.4%,遺傳力較高;多基因遺傳力較低,為0~57.7%。惠云芝等[11]發(fā)現(xiàn)茄子果皮紅色素在各親本間的一般配合力、各組合間的特殊配合力存在顯著差異,顯性基因起主要作用,其狹義遺傳力較低,不利于早代進(jìn)行選擇。
在果菜類蔬菜中,茄子的果萼可以食用,并得到許多消費(fèi)者的偏愛,因此果萼作為茄子果實(shí)表型標(biāo)記性狀也具有重要研究價(jià)值。中國傳統(tǒng)的地方品種果萼顏色與果皮顏色較為一致,近些年從國外引進(jìn)的綠萼黑紫色果皮品種豐富了消費(fèi)市場,同時也給育種者提出了更高的品質(zhì)要求。目前關(guān)于果萼色的遺傳研究較少,喬軍等[12]以綠色和紫色兩個果萼色差異顯著的茄子高代自交系為試驗(yàn)材料,研究茄子果萼色性狀遺傳規(guī)律。結(jié)果表明,雜種F1果萼色為中間型偏向于紫色親本,紫色對綠色為部分顯性。F2分離群體的果萼色級值次數(shù)呈單峰偏態(tài)分布,說明茄子果萼色為多基因控制的數(shù)量性狀;茄子果萼色性狀符合E-0遺傳模型,兩對主基因的效應(yīng)都以加性效應(yīng)為主,且第一對主基因占主導(dǎo)地位,且在F2世代中遺傳率高達(dá)96.84%,因此對其改良可在育種早期F2代進(jìn)行選種。
1.2 果形性狀
果形形成與果實(shí)生長發(fā)育密切相關(guān),同時果實(shí)形狀也是茄子品質(zhì)評價(jià)、分類及定級的重要考核標(biāo)準(zhǔn)。果形分類一般根據(jù)相似幾何形狀或象形描述來區(qū)分,李錫香等[1]將茄子果形分為11類,分別為扁圓、圓球、高圓、卵圓、長卵、短筒、長筒、長條、線形、短羊角和長羊角。在實(shí)際果形評價(jià)中常用指標(biāo)為果實(shí)縱徑、果實(shí)橫徑和果形指數(shù)(果實(shí)縱徑與橫徑的比值)。
茄科作物中果形遺傳多數(shù)符合主基因-多基因的復(fù)合遺傳模型,如番茄[13,14]和辣椒[15]。關(guān)于茄子果形性狀的遺傳分析研究報(bào)道較少。黃銳明等[16,17]以兩個茄子自交系為材料對果長和果徑的遺傳效應(yīng)進(jìn)行了研究,結(jié)果顯示其均屬于數(shù)量遺傳,符合加性-多基因模型,且以加性效應(yīng)為主。并指出控制果長的最少基因數(shù)目為3對,控制果徑性狀的最少基因數(shù)目為4對,兩者狹義遺傳力分別為71.01%和61.45%,說明其受環(huán)境因子的影響較小,可以在早世代進(jìn)行選擇。喬軍等[18]指出茄子果形性狀符合D-2遺傳模型,即符合1對加性主基因+加性-顯性多基因模型;主基因遺傳力以F2代最高(組合Ⅰ可達(dá)52.70%),多基因遺傳力B2代最高(組合Ⅱ可達(dá)66.25%)。茄子果形性狀遺傳總體表現(xiàn)出多基因遺傳特征,果形選擇效率在F2代中最高。果形主要受加性基因控制,在育種工作中要重視親本果形的選擇,便于充分利用基因的累加效應(yīng)。
1.3 果臍性狀
茄子果臍是指花脫落后的痕跡,若痕跡太大會影響果實(shí)表面光滑度,對茄子外觀品質(zhì)產(chǎn)生影響,目前關(guān)于該方面研究報(bào)道較少。王利英等[19]考查了果形指數(shù)、果實(shí)橫縱徑、單果質(zhì)量以及果肉顏色與果臍面積的相關(guān)性。結(jié)果表明,果形指數(shù)和果臍面積呈顯著負(fù)相關(guān),果肉顏色與果臍面積呈顯著正相關(guān),相關(guān)系數(shù)分別為-0.463和0.446。并進(jìn)一步通過通徑分析指出,橫徑對果臍面積的正向直接作用最大,縱徑通過果形指數(shù)對果臍面積有著較大的正向間接作用,果肉顏色對果臍面積的直接作用最小。因此,選擇果形指數(shù)高和縱徑大的材料可以提高對小果臍的選擇效率。
2 茄子遺傳圖譜構(gòu)建的研究進(jìn)展
通過遺傳規(guī)律分析可以大致了解影響茄子果實(shí)性狀形成的基因間相互作用,但若需深入研究各基因在染色體上的位置及功能,則需進(jìn)一步利用分子生物學(xué)相關(guān)研究方法才能實(shí)現(xiàn)。遺傳圖譜的構(gòu)建是遺傳學(xué)基礎(chǔ)研究的一個重要組成部分,對基因的定位和克隆具有重要的意義,并可為育種工作提供指導(dǎo)。茄子遺傳圖譜構(gòu)建工作國外較早開始研究。Nunome等[20]構(gòu)建出第一張茄子遺傳圖譜,隨后Nunome等[21]又以育種品系EPL-1×WCGR112-8的168個F2為作圖群體,構(gòu)建了較完整的茄子遺傳圖譜。該圖譜包括181個位點(diǎn)(88個RAPD和93個AFLP標(biāo)記),包含21條連鎖群,圖譜全長為779.2 cM,平均圖距為4.9 cM;Doganlar等[22]利用RFLP探針(來自番茄的cDNA、基因組DNA和EST)構(gòu)建了較為飽和的茄子比較遺傳圖譜,該圖譜包含12條連鎖群,圖譜全長為1 480 cM,共233個標(biāo)記,同番茄遺傳圖譜有很好的共線性,該圖譜是國際上公認(rèn)的茄子參考圖譜。
Sunseri等[23]以抗黃萎病材料和感病材料為親本創(chuàng)建F2作圖群體,構(gòu)建了一張包含有13個連鎖群的遺傳圖譜。該圖譜包括117個RAPD標(biāo)記和156個AFLP標(biāo)記,全長736 cM。曹必好等[24]以F2群體構(gòu)建了一張RAPD連鎖圖譜,該圖譜包括12個連鎖群,總長度651.2 cM,包含77個標(biāo)記,平均距離為8.57 cM。Wu等[25]利用第二代保守的直向同源序列標(biāo)記集(Conserved ortholog set Ⅱ, COSⅡ)對照參考圖譜構(gòu)建了一張包含有110個COSⅡ標(biāo)記的較為飽和的同線性茄子圖譜,連鎖群數(shù)目與茄子染色體數(shù)相等,連鎖群編號與茄子基因組參考圖譜對應(yīng),為E01-E12,這極大地促進(jìn)了茄科作物比較基因組學(xué)的研究。
喬軍[26]以兩個栽培種茄子高代自交系106和113為親本,利用F2群體構(gòu)建了一張包括23個SSR標(biāo)記和85個AFLP 標(biāo)記共15個連鎖群的遺傳圖譜。該圖譜覆蓋基因組長度1 007.9 cM,平均圖距9.3 cM。Barchi等[27]結(jié)合RAD-Seq技術(shù),利用‘305E40ב67/3組合構(gòu)建了一張包含有415個RFLP和SNP標(biāo)記的飽和遺傳圖譜,該圖譜共有13條連鎖群,其中12條主要連鎖群均具有錨定標(biāo)記,從而可以使得圖譜與茄子參照圖譜很好地對應(yīng)起來。Fukuoka等[28]利用兩個F2作圖群體整合出一張高密度遺傳圖譜,該圖譜包含12條連鎖群,圖譜全長1285.5 cM,共包含952個DNA標(biāo)記,其中313個SSR標(biāo)記,623個SNP標(biāo)記和InDel(Insertion/deletion polymorphisms)標(biāo)記。623個SNP和InDel標(biāo)記主要來自于EST和茄科作物直向同源序列,其中326個標(biāo)記具有很好的保守性可以定位到番茄遺傳圖譜上。Frary等[29]報(bào)道指出已構(gòu)建一張飽和遺傳圖譜,包含了850多個AFLP,RFLP和COS Ⅱ標(biāo)記,平均圖距為1.8 cM,對于后續(xù)進(jìn)行數(shù)量性狀位點(diǎn)(Quantitative trait loci, QTL)定位提供了良好的基礎(chǔ)。
隨著茄科作物基因組學(xué)研究的快速發(fā)展,開發(fā)的新型分子標(biāo)記越來越多,茄子遺傳圖譜也越來越飽和。但目前構(gòu)建圖譜所采用的群體均為F2群體,是非永久性群體。使得已構(gòu)建的圖譜在實(shí)驗(yàn)室間交流和共享較為困難,圖譜整合和利用受到限制,同時很難對茄子數(shù)量性狀進(jìn)行多年的基因定位。
3 茄子果實(shí)性狀相關(guān)QTL定位研究
茄子果實(shí)性狀多為數(shù)量性狀,受QTL和環(huán)境因子的共同作用,因此在茄子果實(shí)品質(zhì)的育種中,可以把控制相應(yīng)性狀的多個QTL先行定位,找出主效QTL,再用基因聚合的方法把多個主效QTL導(dǎo)入到同一個自交系或品種中,就可不受時空限制,提高育種效率。
3.1 與果色相關(guān)的QTL定位
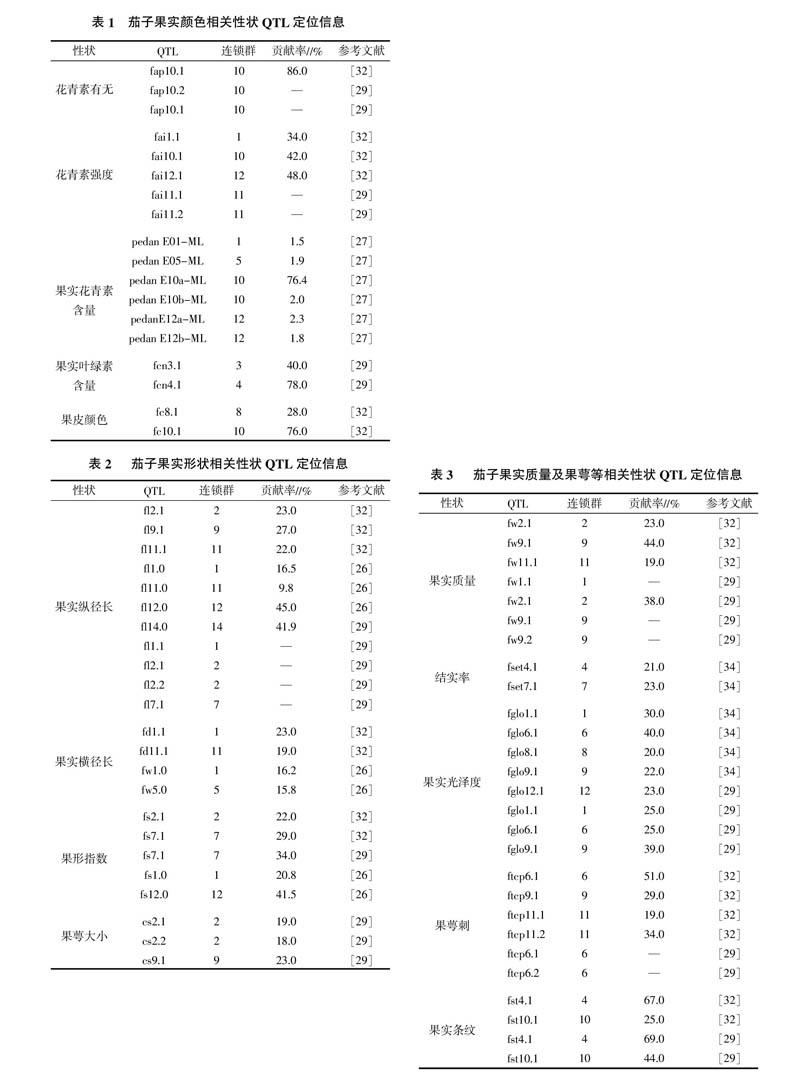
茄子果實(shí)顏色的決定色素主要包括花青素和葉綠素。其中花青素主要分布在茄子表皮的液泡中,隨著其種類和濃度的不同,使得果皮呈現(xiàn)從橘紅過渡至黑紫的不同顏色。葉綠素主要分布在亞表皮細(xì)胞層,決定綠色的產(chǎn)生[30]。因此茄子果色是由兩種色素物質(zhì)的組成、分布、濃度和環(huán)境因子共同作用決定的,是一個復(fù)雜過程。廖毅等[31]以具有不同果皮顏色的茄子及近緣種為材料,采用單株檢測和混合品系法(Bulk segregant analysis,BSA)相結(jié)合篩選到一個與茄子紫紅、紫黑果色相關(guān)的AFLP標(biāo)記,分別命名為EioMi9-1和EioMi9-2,并將該共顯性AFLP標(biāo)記成功轉(zhuǎn)化為SCAR標(biāo)記,可以用于茄子果色分子標(biāo)記輔助育種。利用BSA法篩選分子標(biāo)記效率不高,近些年來學(xué)者們在遺傳圖譜構(gòu)建的基礎(chǔ)上,通過對不同群體F2代果色的表型分析,共定位到18個與果色形成相關(guān)的QTL(表1)。Doganlar等[32]和Frary等[29]分析群體相同,且均采用一年多點(diǎn)的試驗(yàn)方法,獲得結(jié)果較為一致,如fap均位于第10條連鎖群上。但fai和fc等定位結(jié)果不同,其原因可能是Frary等[29]是在Doganlar等[32]已做圖譜基礎(chǔ)上,進(jìn)行進(jìn)一步圖譜飽和,增加了600多個標(biāo)記,提高了作圖和定位效率,因此可以尋找到更多QTL。
Barchi等[27]研究角度不同,主要是通過評價(jià)茄子植株各部位(如莖、葉、花和果實(shí))中花青素含量,并確定相關(guān)QTL位置。其中共定位到6個控制果實(shí)花青素含量的QTL,分別位于第1、第2、第10和第12連鎖群上,其中位于第10連鎖群上的pedanE10a-ML可解釋變異率最高為76.4%。
3.2 與果形相關(guān)QTL定位
茄科作物中番茄的果形遺傳QTL及主效基因定位研究最為深入。Van Der Knaap等[33]分別將控制果形的QTL定位在第2,第3和第11條染色體上(表2),即lifs2、lifs3和lifs11,果長的QTL定位在第7染色體上(lifs7),其中l(wèi)ifs2和lifs7分別是調(diào)控果形ovate基因和sun基因的等位基因。番茄中ovate和sun這兩個主基因控制果形指數(shù),并都已實(shí)現(xiàn)了精細(xì)定位、基因克隆和功能驗(yàn)證。茄子果形QTL目前為止已經(jīng)定位到20個,其中果實(shí)縱徑長fl定位到11個QTL,果實(shí)橫徑長fd(喬軍[26]命名為fw)定位到4個QTL,果形指數(shù)fs定位到5個QTL(表2)。位于第2和第7連鎖群上的fl2.1、fl2.2、fl7.1以及fs2.1、fs7.1是否與sun和ovate存在共線關(guān)系尚不清楚,在以后的研究中可以通過其側(cè)翼標(biāo)記進(jìn)行分子標(biāo)記輔助選擇和分子標(biāo)記加密工作,實(shí)現(xiàn)果形QTL精細(xì)定位和基因克隆功能驗(yàn)證才可確定。此外,喬軍[26]指出控制果形的許多QTL都在同一條染色體上聚集,可能是連鎖的原因,也有可能是一因多效的結(jié)果,因?yàn)榭刂乒L和果徑的基因也會影響果形,這與Doganlar等[32]和Frary等[29]研究結(jié)果較為一致。
3.3 與果重、果萼等相關(guān)QTL定位
Doganlar等[32]和Frary等[29,34]對果實(shí)質(zhì)量、坐果率、果實(shí)光澤度以及果萼刺等性狀進(jìn)行了研究,共定位到27個相關(guān)QTL(表3)。其中8個是控制果實(shí)光澤度(fglo)的QTL,7個是控制果重(fw)的QTL,6個是控制果萼刺(ftcp)的QTL,4個是控制果實(shí)有無條紋(fst)的QTL,2個是控制果實(shí)結(jié)實(shí)率(fset)的QTL。如上所述,兩者所用分析群體相同,定位到的位點(diǎn)也較為一致,如fw2.1、fw9.1、ftcp6.1、fst4.1和fst10.1。但圖譜中標(biāo)記類型不同,尚不能確定是否為同一個數(shù)量基因座位點(diǎn)。隨著茄科作物基因組學(xué)的發(fā)展,番茄、馬鈴薯測序工作的完成,可以通過借助與番茄等作物在基因組同線性關(guān)系,開發(fā)茄子QTLs上與果實(shí)性狀相關(guān)的候選基因。
4 茄子分子育種存在的問題及展望
茄子是僅次于番茄和馬鈴薯的重要茄科作物,在世界蔬菜供給中占有重要的作用。但同番茄和馬鈴薯研究相比,茄子的分子生物學(xué)的研究較為落后,因此加強(qiáng)茄子分子育種的研究將有助于茄子遺傳資源的利用、創(chuàng)新及新品種選育。目前茄子分子育種研究主要集中在單性結(jié)實(shí)、抗病和抗逆性等方面,其基因克隆、功能驗(yàn)證及定位易于研究。而對于數(shù)量基因控制的果實(shí)品質(zhì)性狀研究,國內(nèi)分子水平研究還較為薄弱,國外也局限于幾個研究組,尚未進(jìn)行大規(guī)模研究。同時茄子的基因序列信息相對較少,但隨著番茄全基因組測序的完成,將為茄子的研究提供一個全新的平臺。此外,除番茄外其他茄科植物,如馬鈴薯、辣椒、煙草的測序,都將對茄子分子生物學(xué)研究有重大促進(jìn)作用。借助其他茄科植物的序列信息,可以開發(fā)新型標(biāo)記如SNP、InDel和COSⅡ等,用于茄子重要經(jīng)濟(jì)性狀的分子標(biāo)記、遺傳資源遺傳多樣性分析、指紋圖譜構(gòu)建等。另外深入開展茄子與其他茄科植物間比較基因組學(xué)研究,分離茄子的重要功能基因,將不但有助于了解茄子的遺傳機(jī)制,也有利于茄子的分子標(biāo)記輔助育種實(shí)踐。
參考文獻(xiàn):
[1] 李錫香,朱德蔚. 茄子種質(zhì)資源描述規(guī)范和數(shù)據(jù)標(biāo)準(zhǔn)[M]. 北京:中國農(nóng)業(yè)出版社, 2006.17-18.
[2] 蘇曉梅,劉衛(wèi)東,柳李旺,等. 茄子種質(zhì)資源分類研究進(jìn)展[J]. 中國蔬菜, 2012(10): 1-6.
[3] 吳雪霞,查丁石.我國茄子育種遺傳研究進(jìn)展[J].現(xiàn)代農(nóng)業(yè)科技, 2010(3):143-146.
[4] 李 寧,姚明華,焦春海,等. 亞洲及非洲茄子種質(zhì)資源主要農(nóng)藝性狀的遺傳多樣性分析[J]. 湖北農(nóng)業(yè)科學(xué),2014,53(23): 5769-5774.
[5] TATEBE T. On inheritance of color in Solanum melongena L. [J]. Japanese Journal of Genetics, 1939, 15(5): 261-271.
[6] TATEBE T. Further studies on inheritance of color in Solanum melongena Linn[J]. Japanese Journal of Genetics,1944,20(1): 1-7.
[7] JANICK J, TOPOLESKI L D. Inheritance of fruit color in eggplant (Solanum melongena) [J]. Process of Ameica Society Horticultural Science, 1963, 83: 547-558.
[8] TIGCHELAAR E C, JANICK J, ERICKSON H T. The genetics of anthocyanin coloration in eggplant (Solanum melongena L.) [J]. Genetics, 1968, 60(3): 475-491.
[9] 劉進(jìn)生,PHATAK S C. 茄子功能性雄性不育的遺傳及其與果紫色基因連鎖關(guān)系的研究[J]. 遺傳學(xué)報(bào),1992,19(4):349-354.
[10] 龐文龍,劉富中,陳鈺輝,等. 茄子果色性狀的遺傳研究[J]. 園藝學(xué)報(bào), 2008, 35(7): 979-986.
[11] 惠云芝,孫賀新,林 巖,等. 10個茄子自交系果皮紅色素的配合力遺傳分析[J]. 吉林蔬菜, 2010(4): 32-34.
[12] 喬 軍,劉富中,陳鈺輝,等. 茄子果萼色遺傳研究[J]. 植物遺傳資源學(xué)報(bào), 2011, 12(5): 801-810.
[13] 王 雷,王 鳴,石 英,等. 加工番茄主要數(shù)量性狀遺傳相關(guān)的研究[J]. 西北農(nóng)業(yè)學(xué)報(bào), 1998, 7(1): 32-37.
[14] 成 穎,李海濤,呂書文. 櫻桃番茄果形主基因-多基因混合遺傳分析[J]. 華北農(nóng)學(xué)報(bào), 2008, 23(6): 164-167.
[15] 張樹根,蔣鐘仁,邢永萍,等. 一個辣椒雜交種的加倍單倍體(DH)群體果實(shí)性狀的遺傳分析[J]. 園藝學(xué)報(bào),2008,35(4): 515-520.
[16] 黃銳明,陳國良,謝曉凱,等. 茄子果徑遺傳效應(yīng)初探[J]. 長江蔬菜, 2006(9): 45-46.
[17] 黃銳明,謝曉凱,盧永奮,等.茄子果長遺傳效應(yīng)的初步研究[J]. 廣東農(nóng)業(yè)科學(xué),2006(7):25-26.
[18] 喬 軍,劉富中,陳鈺輝,等. 茄子果形遺傳研究[J]. 園藝學(xué)報(bào), 2011, 38(11): 2121-2130.
[19] 王利英,石 瑤,劉文明,等. 茄子果實(shí)主要性狀與果臍因素的相關(guān)和通徑分析[J]. 天津農(nóng)業(yè)科學(xué), 2008, 14(5): 11-13.
[20] NUNOME T, YOSHIDA T, HIRAI M. Genetic linkage map of eggplant[A]. PALLOIX A, DAUNAY M C. Proceedings of the 10th Eucarpia meeting on genetics and breeding of capsicum and eggplant[C]. Avignon: INRA Paris,1998,239-242.
[21] NUNOME T, ISHIGURO K, YOSHIDA T, et al. Mapping of fruit shape and color development traits in eggplant (Solanum melongena L.) based on RAPD and AFLP markers[J]. Breeding Science,2001,51(1):19-26.
[22] DOGANLAR S, FRARY A, DAUNAY M C, et al. A comparative genetic linkage map of eggplant (Solanum melongena) and its implications for genome evolution in the Solanaceae[J]. Genetics, 2002,161(4):1697-1711.
[23] SUNSERI F, SCIANCALEPORE A, MARTELLI G, et al. Development of RAPD-AFLP map of eggplant and improvement of torance to Verticillium wilt[J]. Acta Horticulturae, 2003, 625: 107-110.
[24] 曹必好,雷建軍,孫秀東,等. 茄子RAPD分子標(biāo)記圖譜的構(gòu)建[J]. 園藝學(xué)報(bào),2006,33(5):1092.
[25] WU F, EANNETTA N T, XU Y, et al. A detailed synteny map of the eggplant genome based on conserved ortholog set II (COS II) markers[J]. Theoretical and Applied Genetics, 2009, 118(5): 927-935.
[26] 喬 軍. 茄子果實(shí)性狀遺傳研究及果形QTL定位[D]. 北京:中國農(nóng)業(yè)科學(xué)院,2011.
[27] BARCHI L, LANTERI S, PORTIS E, et al.A RAD tag derived marker based eggplant linkage map and location of QTLs determining anthocyan in pigmentation[J]. Ploss One, 2012, 7(8): 1-11.
[28] FUKUOKA H,YAMAGUCHI H, NUNOME T, et al. Accumulation, functional annotation, and comparative analysis of expressed sequence tags in eggplant (Solanum melongena L.), the third pole of the genus Solanum species after tomato and potato [J]. Gene,2010,450(1-2):76-84.
[29] FRARY A, FRARY A, DAUNAY M C, et al. QTL hotspots in eggplant (Solanum melongena) detected with a high resolution map and CIM analysis[J]. Euphytica, 2014,197(2): 211-228.
[30] DAUNAY M C, AUBERT S, FRARY A, et al. Eggplant (Solanum melongena) fruit color: pigments, measurements and genetics[A]. VAN HINTUM J, LEBEDA A, PINK D, et al. Proceeding of the 12th Eucarpia meeting on genetics and breeding of capsicum and eggplant[C]. Noordwijkerhout: Centre for Genetic Resources of Wageningen, 2004.108-116.
[31] 廖 毅,孫保娟,孫光聞,等. 與茄子果皮顏色相關(guān)聯(lián)的AFLP及SCAR標(biāo)記[J]. 中國農(nóng)業(yè)科學(xué),2009,42(11):3996-4003.
[32] DOGANLAR S, FRARY A, DAUNAY M C, et al. Conservation of gene function in the Solanaceae as revealed by comparative mapping of domestication traits in eggplant[J]. Genetics,2002,161(4):1713-1726.
[33] VAN DER KNAAP E, TANKSLEY S D. The making of a bell pepper-shaped tomato fruit: Identification of loci controlling fruit morphology in Yellow Stuffer tomato[J]. Theoretical and Applied Genetics,2003,107(1):139-147.
[34] FRARY A, DOGANLAR S, DAUNAY M C, et al. QTL analysis of morphological traits in eggplant and implications for conservation of gene function during evolution of Solanaceous species[J]. Theoretical and Applied Genetics,2003, 107(2):359-370.
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國語大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學(xué)報(bào)(2017年2期)2017-07-05 08:13:02
國際商務(wù)財(cái)會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
