重組FIP的研究進展
2016-10-24 05:49:14段作文陳麗靜李浩戈范文麗李天來林景衛(wèi)
蔬菜 2016年9期
段作文,韓 笑,陳麗靜,李浩戈,張 麗,張 良,范文麗,李天來,林景衛(wèi)*
(1.沈陽農(nóng)業(yè)大學(xué)生物科學(xué)技術(shù)學(xué)院,遼寧 沈陽 110866;2.沈陽農(nóng)業(yè)大學(xué)園藝學(xué)院,遼寧 沈陽 110866)
真菌免疫調(diào)節(jié)蛋白(Fungal immunomodulatory protein,F(xiàn)IP)是近年在高等真菌中發(fā)現(xiàn)的一類與植物凝集素和免疫球蛋白的結(jié)構(gòu)和免疫功能相似的小分子蛋白質(zhì)。1989年,Kino等[1]在靈芝(Ganoderma lucidum)菌絲體提取物中分離得到第一個真菌免疫調(diào)節(jié)蛋白LZ-8(FIP-glu)后,人們又陸續(xù)在松杉靈芝(G. tsugae)、金針菇(Flammulina veltipes)、草菇(Volvariella volvacea)、紫靈芝(G. japoncium)、小孢子靈芝(G. microsporum)、紫芝(G. sinense)、樟芝(Antrodia camphorate)、茯苓(Poria cocos)、云芝(Trametes versicolor)、樹舌靈芝(Ganoderma applanatum;Gene Bank:AEP68179)、黑靈芝(Ganoderma astum)、血紅叢赤殼(Nectria haematococca)中發(fā)現(xiàn)了真菌免疫調(diào)節(jié)蛋白,分別命名為FIP-fve,F(xiàn)IP-gts,F(xiàn)IP-vvo,F(xiàn)IP-gja,F(xiàn)IP-gmi,F(xiàn)IP-gsi,F(xiàn)IP-aca,F(xiàn)IP-pcp,F(xiàn)IP-tvc,F(xiàn)IP-gap,F(xiàn)IP-gas,F(xiàn)IP-nha[2-10]。
目前已知的FIPs的基因編碼區(qū)大小約為330~360 bp、分子量為13 kDa左右,由110~114個氨基酸殘基組成。富含天門冬氨酸和纈氨酸,缺乏組氨酸、半胱氨酸和甲硫氨酸。其N端氨基酸均為乙酰化阻斷氨基酸[11]。在目前所發(fā)現(xiàn)的FIPs中只有LZ-8中含有糖,含量約為1.3%,其他種類的FIPs都不含有糖。不同F(xiàn)IPs在一級結(jié)構(gòu)上非常相似,進化上比較保守,氨基酸序列之間的同源性比較高,可達到60%~100%(圖1)。
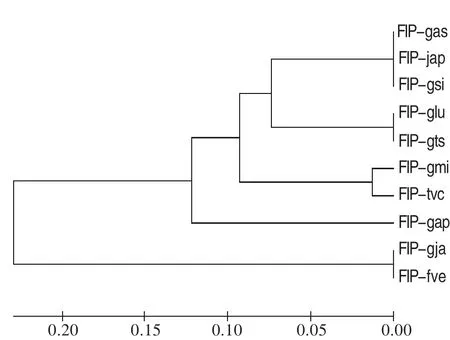
圖1 不同F(xiàn)IP的系統(tǒng)進化樹
臺灣Lin等人利用酵母雙雜交和定點突變的方法,發(fā)現(xiàn)天然有活性的FIPs為同源二聚體,其N端大約10個氨基酸形成α-螺旋,該結(jié)構(gòu)對于FIPs二聚體的形成具有重要作用[12]。還發(fā)現(xiàn)FIPs的二級結(jié)構(gòu)中富含β結(jié)構(gòu),包括2個α-螺旋,7個β-折疊和1個β-轉(zhuǎn)角[13]。該結(jié)構(gòu)特點與免疫球蛋白重鏈可變區(qū)結(jié)構(gòu)非常相似[14]。
2003年,Seow等使用懸滴法獲得了FIP-fve晶體。同年,Paaventhan等利用NaBr滲透對FIP-fve晶體進行單向不規(guī)則晶體衍射,發(fā)現(xiàn)FIP-fve的N端緊接著α-螺旋有一個纖連蛋白折疊結(jié)構(gòu)——FNⅢ折疊,該結(jié)構(gòu)是7個β-折疊S型和8個β-折疊h形中間過渡構(gòu)型,也稱為假-h-型拓?fù)浣Y(jié)構(gòu)。另外,安敏等利用滴氣象擴散法得到了LZ-8的晶體結(jié)構(gòu),與金針菇的晶體結(jié)構(gòu)非常相似[15]。因此表明,各種FIPs在一級結(jié)構(gòu)、二級結(jié)構(gòu)和高級結(jié)構(gòu)上都有一定的相似性,并且在高級結(jié)構(gòu)上與免疫球蛋白有著相似性。
1 真菌免疫調(diào)節(jié)蛋白的生物學(xué)功能
研究證實FIPs具有重要的生物學(xué)功能。具體包括以下幾個方面。
1.1 血細(xì)胞凝集活性
目前已知的幾種FIPs都被發(fā)現(xiàn)具有血細(xì)胞凝集活性,可以凝集不同哺乳動物的血紅細(xì)胞,對血紅細(xì)胞的凝集無種屬特異性和專一性。
LZ-8和LZ-9能夠凝集兔、鼠和綿羊血紅細(xì)胞,但是它們都不能凝集人類四種類型的血紅細(xì)胞(A、B、AB和O血型)[10]。而FIP-fve可以凝集人血紅細(xì)胞(A、B、AB和O血型)。FIP-vvo能凝集Wistar大鼠和綿羊血紅細(xì)胞[16],但它也不能凝集人血紅細(xì)胞。FIP-tvc可凝集鼠血紅細(xì)胞[8]。另外,研究發(fā)現(xiàn)FIP-nha可以凝集綿羊、家兔、小鼠以及人O型血紅細(xì)胞[17]。
1.2 抗腫瘤活性
FIPs都具有抗腫瘤活性。重組表達的靈芝免疫調(diào)節(jié)蛋白(rLZ-8)可以誘導(dǎo)K562細(xì)胞、HL60細(xì)胞和白血病NB4細(xì)胞凋亡[18-19]。rFIP-fve和rFIP-gts都可以抑制A549腫瘤細(xì)胞的生長[20-23]。又發(fā)現(xiàn)FIP-gts還可抑制腫瘤細(xì)胞的移動,而人23-1型基因可以與其協(xié)同作用增強FIP-gts對癌細(xì)胞轉(zhuǎn)移的抑制作用[24]。
1.3 抗過敏反應(yīng)活性
FIPs具有抗過敏反應(yīng)活性。研究發(fā)現(xiàn),LZ-8,F(xiàn)IP-vvo和FIP-fve都可以明顯地抑制小鼠足墊水腫反應(yīng)(Arthus反應(yīng))和小鼠系統(tǒng)過敏性反應(yīng)。LZ-8還可以預(yù)防自主免疫性糖尿病,并且可以提高兔胰腺移植和小鼠皮膚移植的移植率,而且沒有任何副作用[25]。另外,還發(fā)現(xiàn)FIP-fve可以抑制嗜酸性粒細(xì)胞活性,這表明可能對與嗜酸性粒細(xì)胞有關(guān)的過敏癥如支氣管哮喘、過敏性鼻炎等具有療效[26]。FIP-fve還可以抑制卵清蛋白(OVA)引起的食物過敏反應(yīng)和治療羽刺皮癬螨2型抗原(Dp-2)引起的呼吸道炎癥[27]。
1.4 促進淋巴細(xì)胞增殖活性
FIPs可促進小鼠脾細(xì)胞和人類外周血淋巴細(xì)胞增殖,并且呈現(xiàn)出與植物凝集素相似的劑量效應(yīng)曲線。LZ-8,F(xiàn)IP-fve和FIP-vvo能夠刺激淋巴細(xì)胞的增殖[28-30]。同時,F(xiàn)IPs在促進淋巴細(xì)胞增殖過程中與植物凝集素具有協(xié)同作用,LZ-8,F(xiàn)IP-fve和FIP-vvo可分別與ConA,PHA和PHA共同促進淋巴細(xì)胞的分裂。
1.5 誘導(dǎo)細(xì)胞因子表達
研究發(fā)現(xiàn)LZ-8可以增強白細(xì)胞介素-2(interleukin-2,IL-2),IFN-γ,腫瘤壞死因子-α(tumor necrosis factor,TNF-α),IL-β等細(xì)胞因子的分泌[31]。FIP-fve可顯著地增強IL-2和IFN-γ的分泌[29]。而FIP-vvo可以顯著增強IL-2,IL-4,IFN-γ,TNF-α,淋巴毒素(LT)和IL-2的分泌。FIP-gsi也可以增強鼠脾細(xì)胞分泌細(xì)胞因子(IL)-2,IL-3,IL-4,IFN-γ,TNF-α和IL-2受體(IL-2R)[32]。
雖然FIPs具有重要的生物學(xué)功能,但是關(guān)于它們臨床試驗應(yīng)用的報道不是很多,主要是因為不能大量穩(wěn)定地獲取這些蛋白。天然提取這些蛋白既耗時又費力,而且獲得的產(chǎn)品率很低,活性也不高,因此,怎樣高效率地表達這些FIPs,對于今后的實踐應(yīng)用至關(guān)重要。因此,目前人們廣泛利用基因工程的手段構(gòu)建一些表達載體使它們高效、穩(wěn)定地表達,以期為將來的臨床應(yīng)用奠定基礎(chǔ)。
2 FIPs的重組表達
目前,人們用于體外蛋白重組表達的系統(tǒng)主要有原核表達系統(tǒng)和真核表達系統(tǒng)兩種,原核表達系統(tǒng)主要是大腸桿菌表達系統(tǒng),而真核表達系統(tǒng)主要包括酵母表達系統(tǒng)、植物表達系統(tǒng)、昆蟲細(xì)胞表達系統(tǒng)(見表1)。
2.1 LZ-8的重組表達
2.1.1 LZ-8在大腸桿菌表達系統(tǒng)中的重組表達
采用CTAB法提取靈芝DNA,通過PCR擴增LZ-8基因,擴增產(chǎn)物可分別與載體pET21a,pET28b,pET30a連接,將連接產(chǎn)物轉(zhuǎn)化到宿主細(xì)胞大腸桿菌BL21(DE3)中,經(jīng)誘導(dǎo)培養(yǎng),檢測出rLZ-8的產(chǎn)量分別是占菌體總蛋白的36.52%,70 mg/L,34.46 mg/L。
生物活性測定表明,rLZ-8蛋白在一定程度上可以延遲小鼠的免疫排斥過程,延長移植體存活時間,具有一定的免疫抑制活性,還可以凝集羊血紅細(xì)胞[32-35]。
2.1.2 LZ-8在酵母表達系統(tǒng)中的重組表達
克隆LZ-8基因,克隆后的LZ-8基因可與載體pPIC9p和PICZαA連接,通過電擊轉(zhuǎn)化法將連接產(chǎn)物轉(zhuǎn)化到畢赤酵母GS115中,經(jīng)甲醇連續(xù)誘導(dǎo)培養(yǎng),rLZ-8的表達效率分別為191.2 mg/L和350 mg/L[36-37]。
生物活性分析表明,rLZ-8可以刺激小鼠脾淋巴細(xì)胞的增殖、增強鼠脾細(xì)胞分泌細(xì)胞因子IL-2,也可以抑制人白血病-NB4細(xì)胞的增殖。
LZ-8基因還可與載體pPICZαA連接、轉(zhuǎn)化到畢赤酵母X-33中,經(jīng)培養(yǎng),表達效率可達34.4 mg/L。

表1 各種重組表達系統(tǒng)的比較
生物活性測定表明,rLZ-8可以凝集兔、鼠、綿羊和人血紅細(xì)胞[10]。
2.1.3 LZ-8植物表達系統(tǒng)中的重組表達
擴增LZ-8基因,與載體p3300GB連接,構(gòu)成植物雙元表達載體pCAMLz8,將該載體轉(zhuǎn)化到農(nóng)桿菌LBA4404中,最后通過葉盤轉(zhuǎn)化法轉(zhuǎn)到煙草中。經(jīng)測定,rLZ-8在煙草中的表達量占煙草可溶性蛋白的0.18%,即每克新鮮葉片約含149.82 μg的rLZ-8[38]。
2.1.4 LZ-8在枯草芽孢桿菌和乳酸乳球菌中的重組表達
采用OE-PCR法擴增LZ-8基因,擴增后的LZ-8基因分別與載體pOA和pOAS連接,連接產(chǎn)物轉(zhuǎn)化到芽孢桿菌WB800細(xì)胞中,經(jīng)測定,rLZ-8表達效率分別是17.5 mg/L和13.2 mg/L。而在乳酸菌中的表達,LZ-8基因與載體pNZSLZ連接,轉(zhuǎn)化到宿主細(xì)胞乳酸菌NZ9000細(xì)胞中,表達效率為1.24 mg/L。
生物活性測定表明,不同表達載體表達的rLZ-8具有不同的生物活性。枯草芽孢桿菌WB800(pOA-LZ8)中表達的rLZ-8可以促進hPBMC的增殖,而在乳酸乳球菌中表達的NZ9000(pNZSLZ)雖然可以促進hPBMC的增殖,但是不顯著。枯草芽孢桿菌產(chǎn)生的rLZ-8也可以促進TNF-α的產(chǎn)生,而在NZ9000(pNZSLZ)產(chǎn)生的rLZ-8沒有此活性。三種來源的rLZ-8都可以顯著地增加IL-2/IL-4的比率[39]。
2.2 FIP-fve的重組表達
2.2.1 FIP-fve在大腸桿菌表達系統(tǒng)中的重組表達
構(gòu)建重組表達載體PET30a-FIP-fve,該載體可分別轉(zhuǎn)化到大腸桿菌DE3和M15中,經(jīng)培養(yǎng),表達效率分別為29.1 mg/L[40]和得到的可溶性蛋白占M15總可溶性蛋白的7.4%[30]。同時,F(xiàn)IP-fve基因也可與載體pGEX4t連接,轉(zhuǎn)化到大腸桿菌DE3細(xì)胞中。經(jīng)培養(yǎng),表達效率為5.8 mg/L[41]。FIP-fve基因還可以連接到pET-28(+)載體上,通過熱轉(zhuǎn)化法轉(zhuǎn)化到大腸桿菌BL21細(xì)胞中。經(jīng)培養(yǎng),表達效率可達30 mg/L。
生物活性分析表明,rFIP-fve可以增強鼠血清中細(xì)胞因子IL-2和IFN-γ的分泌[42],也具有刺激細(xì)胞分裂的作用。
2.2.2 FIP-fve在酵母表達系統(tǒng)中的重組表達
金針菇可以在酵母中進行誘導(dǎo)型和組成型表達。擴增金針菇和酵母三磷酸甘油醛脫氫酶啟動子Pgap基因,然后構(gòu)建畢赤酵母誘導(dǎo)型表達載體pPIC9-FIP-fve和組成型表達載體pPIC9-PGAp-FIP-fve,最后,將表達載體轉(zhuǎn)化到畢赤酵母GS115細(xì)胞中。經(jīng)培養(yǎng),誘導(dǎo)型表達rFIP-fve的表達效率是158.2 mg/L,組成型表達rFIP-fve的表達效率為46.3 mg/L[43]。
生物活性分析表明,rFIP-fve可以凝集人血紅細(xì)胞、刺激小鼠脾淋巴細(xì)胞的增殖,還可顯著地刺激小鼠脾淋巴細(xì)胞分泌白介素IL-2。
此外,F(xiàn)IP-fve還可與載體pPICZαA連接,連接產(chǎn)物轉(zhuǎn)化到畢赤酵母X-33細(xì)胞中,經(jīng)甲醇連續(xù)誘導(dǎo)培養(yǎng),表達效率為18.9 mg/L。
生物活性分析表明rFIP-fve可以凝集兔的血紅細(xì)胞[10]。
2.2.3 FIP-fve在昆蟲表達系統(tǒng)中的重組表達
采用PCR法擴增FIP-fve基因,PCR產(chǎn)物與轉(zhuǎn)移載體質(zhì)粒pAcUW21結(jié)合形成質(zhì)粒pAcP10Fve,然后,該質(zhì)粒pAcP10Fve與事先合成的SPbbx(單肽)片段結(jié)合,形成重組質(zhì)粒pAcP10SPbbxFve,再與AcMNPV一起合成重組病毒vAcP10SPbbxFve,最后感染昆蟲Sf21細(xì)胞。經(jīng)培養(yǎng),F(xiàn)IP-fve表達效率為6.25 mg/L。
生物活性測定表明rFIP-fve可以顯著地刺激鼠脾細(xì)胞分泌IL-2[44]。
2.3 FIP-gts的重組表達
2.3.1 FIP-gts在大腸桿菌表達系統(tǒng)中的重組表達
擴增FIP-gts基因,擴增產(chǎn)物與載體pET-30a(+)連接,連接產(chǎn)物轉(zhuǎn)化到大腸桿菌TG1細(xì)胞中,經(jīng)培養(yǎng),測定其表達效率為20 mg/L[2]。
2.3.2 FIP-gts在昆蟲表達系統(tǒng)中的重組表達
擴增FIP-gts基因,從轉(zhuǎn)移載體質(zhì)粒pAcUW21中構(gòu)建重組質(zhì)粒pAcP10Fve,然后該重組質(zhì)粒與AcRP23.LacZ DNA共轉(zhuǎn)染Sf21細(xì)胞,產(chǎn)生重組桿狀病毒vAcP10Fve。最后將vAcP10Fve轉(zhuǎn)化到Sf21細(xì)胞中,經(jīng)培養(yǎng),測定3×106受感染的細(xì)胞rFIP-gts表達效率是47.2 μg。
生物活性測定表明,rFIP-gts可以增強鼠脾細(xì)胞分泌細(xì)胞因子IL-2[45]。
2.4 FIP-vvo的重組表達
擴增FIP-vvo基因,然后與載體pPICZaA連接。通過電轉(zhuǎn)化法將連接產(chǎn)物轉(zhuǎn)化到畢赤酵母X-33細(xì)胞中。經(jīng)甲醇連續(xù)誘導(dǎo)培養(yǎng),表達效率達到了410 mg/L。
生物活性測定表明rFIP-vvo可以凝集鼠和綿羊的血紅細(xì)胞、刺激鼠脾細(xì)胞的增殖、增強鼠脾細(xì)胞中細(xì)胞因子IFN-γ的分泌[16]。
2.5 FIP-gja的重組表達
2.5.1 FIP-gja在大腸桿菌表達系統(tǒng)中的重組表達
采用PCR法擴增FIP-gja基因。擴增產(chǎn)物與載體pET-30a(+)連接,轉(zhuǎn)化到大腸桿菌BL21細(xì)胞中,經(jīng)培養(yǎng),表達效率為36.46 mg/L[46]。
2.5.2 FIP-gja在植物表達系統(tǒng)中的重組表達
采用PCR法擴增FIP-gja基因。PCR產(chǎn)物先后與載體pBluecsriPt和p3300GB連接,得到FIP-gja植物雙元表達載體pcAMGJA。并將該載體轉(zhuǎn)化到農(nóng)桿菌LBA4404中,通過葉盤轉(zhuǎn)化法轉(zhuǎn)入煙草。經(jīng)鑒定,其表達效率為可溶性總蛋白的0.31%。
生物學(xué)功能分析表明,rFIP-gja可增強TNF-α的表達水平[46]。
2.6 FIP-gsi的重組表達
2.6.1 FIP-gsi在大腸桿菌表達系統(tǒng)中的重組表達
FIP-gsi基因與載體pET-30a(+)連接,連接產(chǎn)物可分別轉(zhuǎn)化到大腸桿菌BL21和M15細(xì)胞中,通過IPTG誘導(dǎo)培養(yǎng),表達的rFIP-gsi分別約占大腸桿菌總蛋白的46.1%和得到的可溶性蛋白占M15總可溶性蛋白的25%[47-48]。
生物活性測定表明,rFIP-gsi能夠誘導(dǎo)細(xì)胞因子IL-2,IL-3,IL-4,IFN-γ,TNF-α,LT及IL-2R(IL-2 receptor)表達。
2.6.2 FIP-gsi在擔(dān)子菌類灰蓋鬼傘中的重組表達
構(gòu)建重組質(zhì)粒pBFIP-gsi,然后將重組質(zhì)粒轉(zhuǎn)化到灰蓋鬼傘菌株LT2細(xì)胞中,經(jīng)培養(yǎng),表達效率為每千克新鮮的菌絲體中可以提取314 mg的rFIP-gsi。
凝血實驗表明,rFIP-gsi可以凝集小鼠血紅細(xì)胞,但不能凝集人的血紅細(xì)胞[49]。
2.7 FIP-tvc重組表達
2.7.1 FIP-tvc在大腸桿菌表達系統(tǒng)中的重組表達
將擴增后的FIP-tvc基因與載體pET21a(+)連接,轉(zhuǎn)化到宿主細(xì)胞大腸桿菌BL21細(xì)胞中。經(jīng)IPTG誘導(dǎo)培養(yǎng),重組表達的蛋白有20%是以可溶性蛋白的形式存在[8]。
生物學(xué)活性分析表明,rFIP-tvc可以凝集大鼠和小鼠血紅細(xì)胞,可以有選擇性地增強小鼠脾細(xì)胞分泌白細(xì)胞介素(IL)-1α,IL-2,IL-5,IL-6,TNF-α,LT[8]。
2.7.2 FIP-tvc在酵母表達系統(tǒng)中的重組表達
PCR法擴增FIP-tvc基因,然后與載體pPIC9連接,最后將連接產(chǎn)物轉(zhuǎn)化到畢赤酵母GS115細(xì)胞中,經(jīng)甲醇連續(xù)誘導(dǎo)培養(yǎng),其表達效率可達41.06 mg/mL[50]。
2.8 LZ-9的重組表達
擴增LZ-9基因,與載體pPICZα-A連接,轉(zhuǎn)化到畢赤酵母菌株X-33細(xì)胞中,經(jīng)培養(yǎng),表達效率為6.7 mg/L。
血凝試驗分析表明,rLZ-9可以凝集兔、鼠和綿羊血紅細(xì)胞[10]。

表2 已知的FIPs重組表達現(xiàn)狀
2.9 FIP-nha的重組表達
2.9.1 FIP-nha在大腸桿菌表達系統(tǒng)中的重組表達
擴增FIP-nha基因,擴增產(chǎn)物與GST融合載體pGEX-4T-1連接,連接產(chǎn)物轉(zhuǎn)化到大腸桿菌Rosetta細(xì)胞中。經(jīng)培養(yǎng),表達效率為42.7 mg/L[17]。
生物活性分析表明,rFIP-nha可以凝集人類和兔血紅細(xì)胞、顯著地刺激小鼠脾淋巴細(xì)胞增殖,且不具有協(xié)同效應(yīng),有增強鼠脾細(xì)胞分泌IL-2的能力,還可以誘導(dǎo)HL60,HepG2,MGC823癌癥細(xì)胞的凋亡,抑制MGC-823和HepG2細(xì)胞的增殖[17]。
2.9.2 FIP-nha在酵母表達系統(tǒng)中的重組表達
從血紅叢赤殼中獲得FIP-nha基因,然后與載體pPICZα-A連接,連接產(chǎn)物轉(zhuǎn)化到畢赤酵母菌株X-33細(xì)胞中,經(jīng)培養(yǎng),表達效率為18.9 mg/L。
生物活性測定表明rFIP-nha可以凝集鼠、兔、綿羊和胰蛋白酶處理過的人血紅細(xì)胞[10]。
2.1 0 FIP-SN15的重組表達
FIP-SN15是通過基因改組技術(shù),從LZ-8和FIP-gsi兩種真菌免疫調(diào)節(jié)蛋白的基因中合成的重組DNA序列[51]。
擴增FIP-SN15基因,然后與載體pET-30a連接,連接產(chǎn)物轉(zhuǎn)化到大腸桿菌BL21細(xì)胞中,經(jīng)IPTG誘導(dǎo)培養(yǎng),表達效率為35.95 mg/L[40]。
生物活性分析表明,rFIP-SN15可以凝集鼠血紅細(xì)胞、顯著地增強細(xì)胞因子IL-2,IL-4,IFN-γ,IL-2R和LT分泌[52],也可以誘導(dǎo)人類膠質(zhì)母細(xì)胞瘤細(xì)胞系U-251 MG的凋亡。
以上關(guān)于已知的FIPs重組表達現(xiàn)狀的總結(jié)見表2。
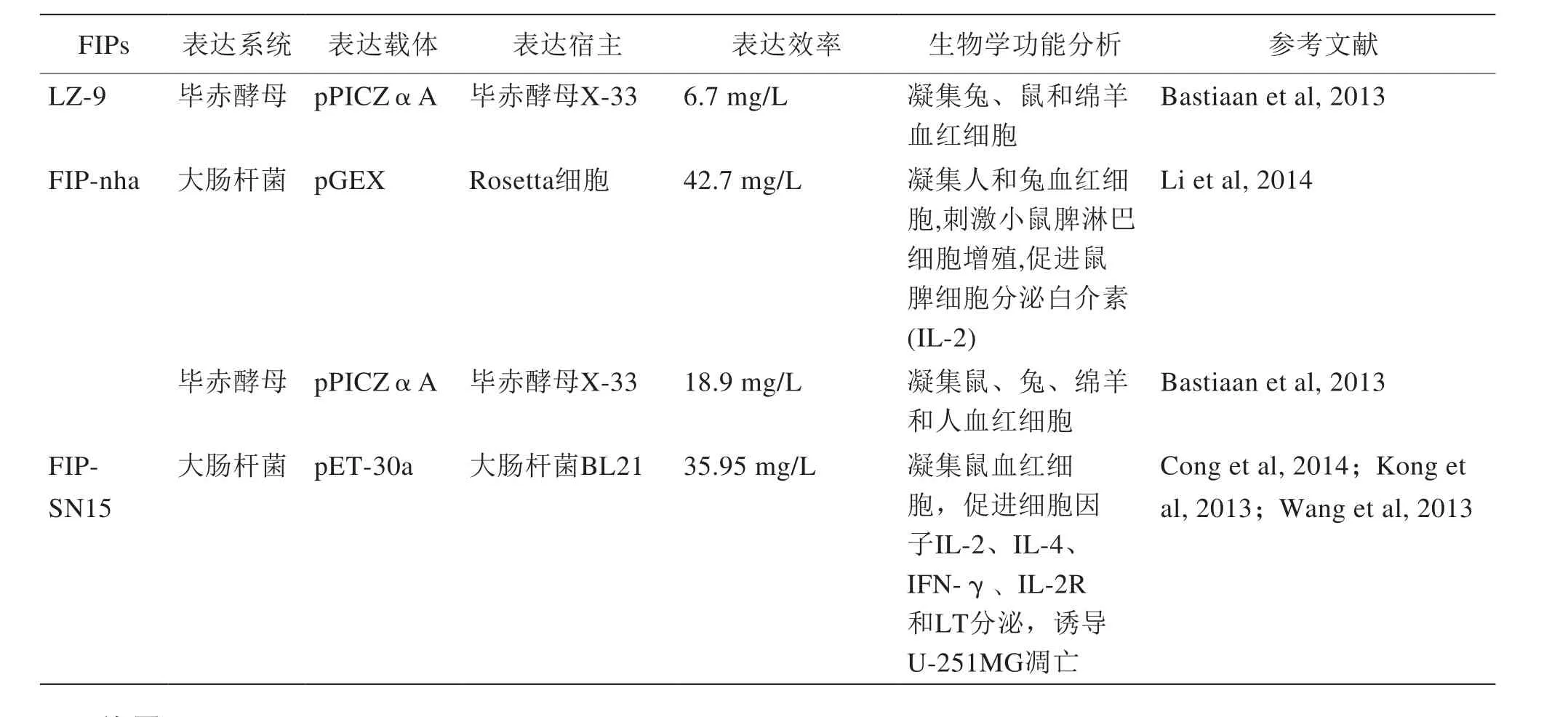
續(xù)表2
3 前景
真菌免疫調(diào)節(jié)蛋白是一種與植物凝集素和免疫球蛋白的結(jié)構(gòu)和免疫功能相似的小分子蛋白質(zhì),具有抗腫瘤、抗過敏、刺激淋巴細(xì)胞增殖等免疫活性,同時研究結(jié)果未發(fā)現(xiàn)它們具有副作用,因此,F(xiàn)IPs的研究具有廣泛的應(yīng)用發(fā)展空間。但是,天然提取的FIPs的產(chǎn)出率很低,費時耗力且成本較高,所以,尋求一個可以高效重組表達這些FIPs的技術(shù)是今后研究的主要方向。近年來,隨著分子生物學(xué)和生物工程技術(shù)的發(fā)展,人們可以利用基因工程菌株工業(yè)生產(chǎn)藥用蛋白質(zhì),這就為人們后續(xù)進一步研究FIPs應(yīng)用于實踐提供了重要的手段。總之,關(guān)于FIPs的研究將會越來越受到重視,尤其作為新型免疫調(diào)節(jié)劑,或者食品添加劑方面具有重大應(yīng)用潛力。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
甘肅教育(2020年14期)2020-09-11 07:57:42
中學(xué)生數(shù)理化(高中版.高考數(shù)學(xué))(2020年5期)2020-06-02 09:19:08
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級)(2017年10期)2017-10-10 01:00:05
商周刊(2017年9期)2017-08-22 02:57:49
時代英語·高二(2015年1期)2015-03-16 00:08:11
中國衛(wèi)生(2014年11期)2014-11-12 13:11:32
云南中醫(yī)學(xué)院學(xué)報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
