HSP70及Ezrin蛋白表達變化對人成骨肉瘤MG63細胞凋亡和增殖的影響
2016-10-24 00:44:33馬國濤孫曉茹
山東醫藥 2016年29期
關鍵詞:檢測
馬國濤,孫曉茹
(1平頂山市第一人民醫院,河南平頂山467000;2平頂山市中醫院)
?
HSP70及Ezrin蛋白表達變化對人成骨肉瘤MG63細胞凋亡和增殖的影響
馬國濤1,孫曉茹2
(1平頂山市第一人民醫院,河南平頂山467000;2平頂山市中醫院)
目的分析人成骨肉瘤MG63細胞中熱休克蛋白70(HSP70)、細胞骨架蛋白(Ezrin)的表達對該細胞凋亡和增殖的影響。方法 首先在有雙啟動子的質粒載體pGFP-V-RS中構建能夠表達HSP70和Ezrin蛋白shRNA的載體并重組質粒,將質粒轉染入MG63細胞,分為聯合表達組、單純shRNA組、空白載體組,篩選穩定表達細胞培養增殖,采用RT-PCR及Western blotting法檢測這兩種蛋白基因表達情況與蛋白水平;Western blotting法檢測凋亡相關蛋白,使用流式細胞術及噻唑藍比色法分別檢測第1、3、5、7天細胞凋亡及增殖率。結果 聯合表達組Ezrin mRNA及Ezrin蛋白合成量低于空白載體組和單純shRNA組,HSP70 mRNA及HSP70蛋白合成量高于空白載體組和單純shRNA組(P均<0.01),HSP70和Ezrin聯合表達的MG63細胞構建成功。低表達Ezrin和高表達HSP70的載體質粒構建成功。聯合表達組MG63細胞凋亡率(23.94%±0.48%)較空白載體組(10.96%±0.25%)高(P<0.01),增殖速度(308.67%±2.91%)較單純shRNA組(394.80%±2.63%)慢(P<0.01);與空白載體組比較,單純shRNA組、聯合表達組第3、5、7天細胞增殖率降低,抑制率升高(P均<0.01)。與單純shRNA組比較,聯合表達組第3、5、7天細胞增殖率低,抑制率低(P均<0.01)。結論 高表達的HSP70和低表達的Ezrin在促進骨肉瘤細胞凋亡的同時減緩其增長速度。
骨肉瘤細胞;熱休克蛋白質70;Ezrin蛋白;shRNA
骨肉瘤是一種多發于年輕人群的具有較高轉移率和復發率的惡性腫瘤,近80%的患者在診斷明確時已發生癌細胞轉移,在較新的放化療治療手段下,其5年存活率仍在一半以下,而治療后約有三分之一的患者3年后再次被檢出癌細胞[1],所以,研究骨肉瘤細胞的凋亡和增殖機制,發現其干擾新方法對于臨床治療非常有意義。Ezrin蛋白存在于細胞表面,作為連接和受體蛋白存在,通過多種信號途徑調節癌細胞凋亡、增殖、浸潤、轉移[2]。系統性分析表明,骨肉瘤細胞中Ezrin蛋白過表達會導致患者5年生存率顯著降低[3]。但Ezrin參與腫瘤細胞凋亡、增殖、局部浸潤、轉移的相關機制尚未清楚,而目前單一靶蛋白很難完全殺死體內所有癌細胞,所以復合的多靶點基因表達治療是較有意義的研究方向[4]。熱休克蛋白(HSP)是一種應激性蛋白,在病理條件下,HSP能夠誘導產生特異性細胞毒性T淋巴細胞(CTL),增加HSP70在癌組織細胞中的含量能夠干預腫瘤細胞生長,并對癌細胞的清除有一定影響[5]。本文探討了HSP70和Ezrin蛋白shRNA聯合對于人成骨肉瘤MG63細胞凋亡和增殖的影響。現報告如下。
1 材料與方法
1.1細胞系及細胞培養MG63系人骨肉瘤細胞,于含10%胎牛血清的RPMI1640培養基中培養,置于空氣中CO2體積為5%、溫度37 ℃的環境中,培養基及胎牛血清均購自Hyclone公司。
1.2表達載體的構建①Ezrin蛋白shRNA表達載體的構建:在Genbank數據庫中查詢Ezrin蛋白的mRNA堿基序列,得到BC013903.2堿基順序,合成一端帶有HindⅢ內切酶酶切位點,另一端帶有BamHⅡ內切酶酶切位點,中部含有5′-TGCTATGTTGGAATACCTTTCAAGAGAAGGTATCCAACATAGCAT-3′發卡序列的DNA片段。將其引入載體pGFP-V-RS,合成pGFP-V-RS-shRNA轉化感受態細胞JM 10,經過增殖篩選并經北京諾禾致源生物科技公司測序后,確認為目標載體。②HSP70與Ezrin蛋白shRNA共表達載體的構建:在Genbank數據庫中查詢HSP70的mRNA堿基序列,得到L12723.1堿基順序并合成HSP70基因片段,通過PCR將載體pGFP-V-RS的GFP段替換為HSP70片段,經過增殖篩選并經測序后,確認為目標載體pGFP-V-RS-HSP70。按上述操作建立目標載體pGFP-V-RS-shRNA-HSP70。
1.3MG63細胞株的轉染及分組將質粒轉染入MG63細胞,分為聯合表達組(pGFP-V-RS-shRNA-HSP70)、單純shRNA組(pGFP-V-RS-shRNA)、空白載體組(pGFP-V-RS),每組70份。使用DNAfectin 2100高效轉染試劑轉染,200 μg/mL遺傳霉素418篩選培養1個月,擴增后得到穩定表達的細胞。PCR法鑒定轉染細胞。
1.4MG-63細胞中Ezrin、HSP70 mRNA的檢測采用RT-PCR法。收集穩定轉染細胞株,加入1 mL TRIzol溶液進行總RNA提取,行反轉錄反應。在M-MLV反轉錄酶(Promega)作用下反轉錄為cDNA。PCR引物為Ezrin-F:5′-ACTCACCAGAAACCGAAAATG-3′、Ezrin-R:5′-TGGAGGCCAAAGTACCACAC-3′,擴增產物159 bp;HSP70-F: 5′-AAGAGCAACAGCAG-CAGACA-3′、HSP70-R:5′-CGATFGGCAGGTCCA-CAGTA-3′,擴增產物150 bp;內參為18 s RNA-F:5′-CCTGGATACCGCAGCTAGGA-3′、18 s RNA-R:5′-GCGGCGCAATACGAATGCCCC-3′,擴增產物112 bp。參考熒光定量PCR試劑盒操作指導進行相對定量PCR實驗,實驗條件為:單個樣本重復3次,50 ℃、2 min預變性;95 ℃、2 min;95 ℃、15 s,60 ℃、32 s,讀板(40次循環)。
1.5MG-63細胞中Ezrin、HSP70蛋白的檢測采用Western blotting法。將穩定表達細胞中的所有蛋白質提取,經SDS-PAGE凝膠電泳后轉移到PVDF膜上可得清晰蛋白條帶,后封閉1 h,加入一抗GADPH、Ezrin或HSP70,稀釋度1∶1 000,二抗為羊抗兔血清IgG,稀釋度1∶20 000,按順序孵育后,經化學發光反應及X線片處理得到圖像,分光密度儀檢測條帶凈光密度值,計算相對凈光密度值。
1.6MG-63細胞凋亡率、增殖率的檢測①細胞凋亡率的檢測:采用流式細胞術。參考Annexin V-FITC細胞凋亡檢測試劑盒相關指導步驟,將穩定表達的細胞加入緩沖液0.5 mL制作成細胞懸液,每份含有5×105個細胞,加入1.25 μL Annexin V-PE后在25 ℃下避光孵育,15 min后離心洗脫,加入預冷1×結合緩沖液后第1、3、5、7天檢測,檢測3次得到均值。Annexin-V陰性、PI陰性代表正常細胞,Annexin-V陽性、PI陰性代表凋亡早期的細胞,Annexin-V陽性、PI陽性代表凋亡晚期的細胞。② 細胞增殖能力的檢測:采用噻唑藍比色(MTT)法。轉染后穩定表達的細胞稀釋并接種于96孔板,保證每孔細胞數為1~2×103,加入20 μL PMS/MTS混合液,培養4 h后測定吸光度,持續7 d,檢測3次得到均值。

2 結果
2.1轉染情況經轉染的細胞培養1個月后,轉染成功率約70%,選擇狀態較佳的細胞,在熒光顯微鏡下觀測,聯合表達組和單純shRNA組細胞中都表現出HSP70-GFP顯示為綠色熒光。
2.2各組Ezrin、HSP70 mRNA及蛋白表達比較見表1。
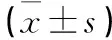
表1 各組Ezrin、HSP70 mRNA及蛋白表達比較
注:與空白載體組相比,*P<0.01;與單純shRNA組相比,#P<0.01。
2.3各組細胞凋亡率比較 見表2。

表2 各組細胞凋亡率比較
注:與空白載體組相比,*P<0.01;與單純shRNA組相比,#P<0.01。
2.4各組不同時間細胞增殖率及抑制率比較見表3。

表3 各組不同時間細胞增殖率及抑制率比較±s)
注:與空白載體組同時間相比,*P<0.01;與單純shRNA組同時間相比,#P<0.01。
3 討論
Ezrin基因和HSP70基因存在于每一個人體細胞中,但正常狀態下,絕大多數細胞的兩種基因處于休眠狀態,而在部分骨髓瘤細胞中發現了Ezrin蛋白或HSP70蛋白同時或單獨存在,兩種蛋白的表達強度并不一致。本研究為避免干擾,保持實驗的可重復性,構建了雙基因穩定表達載體。pGFP-V-RS是一個攜帶CMV和pU6雙啟動位點的載體,在插入Ezrin shRNA片段的同時將HSP70標記GFP綠色熒光蛋白基因后代替質粒上的GFP序列,構建出雙靶點調控pGFP-V-RS質粒載體,插入shRNA并替換GFP序列為HSP70序列后可以在兩個啟動子分別啟動表達。本研究將穩定轉染細胞經過培養1個月后,熒光顯微鏡及熒光定量PCR顯示HSP70-GFP穩定表達,與空白載體組比較,聯合表達組Ezrin mRNA及蛋白質表達均下降,HSP70 mRNA及蛋白表達均升高;單純shRNA組Ezrin mRNA及蛋白下降。單純shRNA與聯合表達組上述各指標比較,P均<0.01。提示聯合表達的MG63細胞構建成功。
使細胞中Ezrin基因低表達的方法有siRNA干擾、shRNA抑制等,也可以使Ezrin基因結構發生變化失去生物作用。考慮到siRNA及改變蛋白結構的方法持續時間短,不能長期觀察。筆者選擇了應用shRNA抑制的方法。而在質粒的選擇上,Zhang等[6]研究發現引入pGenesil-1質粒的Ezrin基因的shRNA雖然穩定轉染MG63,但沉默效率較低;而Gouri-Fard等[7]發現pGFP-V-RS載體質粒能夠穩定利用shRNA抑制RHOXFl基因,沉默效率超過80%,但是轉染的細胞為乳腺癌細胞;另外,周永春等[8]也發現,以pGFP-V-RS為載體的shRNA抑制Sox4基因,沉默效率較高。提示利用pGFP-V-RS載體質粒對于基因的shRNA抑制效果較好,沉默率較高。而本研究將Ezrin基因轉錄的shRNA引入載體轉染細胞后,發現Ezrin蛋白的合成受到抑制,沉默效率超過95%;而Ezrin基因的shRNA在轉染后36 h表達最低,同時延遲12 h出現蛋白表達最低,這可能與調控基因表達的時序性有關。
本研究中利用流式細胞術統計到穩定轉染的MG63腫瘤細胞的早期自發凋亡率為4.2%,晚期自發凋亡率6.6%。Bcl-2常和Bax結合形成特異性同源二聚體,一起介導細胞凋亡,當促進凋亡的Bax生物效果超過抗凋亡蛋白Bcl-2時,表現為促進凋亡,反之抑制凋亡。另外,本研究發現,當shRNA抑制Ezrin蛋白的基因表達時,細胞的各期凋亡率均顯著提高。Lévy等[9]研究發現,Ezrin蛋白通過NF-vB通道使Bcl-2高表達而Bax低表達介導凋亡。Wick等[10]發現,Ezrin通過連接抗原-CD44及HSP70,在其他炎癥因子的介導下,提高Bcl-2水平、抑制膠質細胞瘤凋亡;另外Wick等發現此過程中提高了基質金屬蛋白酶(MMPs)合成,也促進癌細胞的分裂。
通常認為,腫瘤細胞凋亡的同時伴隨著細胞增殖率的降低。有研究認為,抑制Ezrin基因表達或將其蛋白結構改變,可以通過ROCK信號通路傳導,抑制MMPs基因。本研究中與空白載體組比較,單純shRNA組、聯合表達組第3、5、7天增殖率降低、抑制率均升高,聯合表達組第3、5、7天增殖率及抑制率均低于shRNA組。提示高表達的HSP70基因和低表達的Ezrin基因在促進骨肉瘤細胞凋亡的同時減緩其增長速度,但本研究還需要繼續在腫瘤動物模型進行體內研究,才能更準確地評估其對瘤細胞的凋亡和增殖的影響。
[1] Hofbauer LC, Ten RM, Khosla S. The anti-androgen hydroxyflutamide and androgens inhibit interleukin-6 production by an androgen-responsive human osteoblastic cell line[J]. J Bone Miner Res, 1999,14(8):1330-1337.
[2] Riggs BL, Khosla S, Melton LJ. A unitary model for involutional osteoporosis: estrogen deficiency causes both type I and type II osteoporosis in postmenopausal women and contributes to bone loss in aging men[J]. J Bone Miner Res, 1998,13(5):763-773.
[3] 林秾,葉招明,李玢,等.成人骨肉瘤的臨床特點及預后影響因素[J].中華骨科雜志,2009,29(8):717-722.
[4] Lacey DL, Timms E, Tan HL, et al. Osteoprotegerin ligand is a cytokine that regulates osteoclast differentiation and activation[J]. Cell, 1998,93(2):165-176.
[5] Prestwood KM, Kenny AM, Unson C, et al. The effect of low dose micronized 17B-estradiol on bone turnover, sex hormone levels, and side effects in older women: a randomized, double blind, placebo-controlled study[J]. J Clin Endocrinol Metab, 2000,85(12):4462-4469.
[6] Zhang J, Zuo J, Lei M, et al. Ezrin promotes invasion and migration of the MG63 osteosarcoma cell[J]. Chin Med J, 2014,127(10):1954-1959.
[7] Gouri-Fard S, Abdollahi DZ, Omrani M, et al. shRNA mediated RHOXFl silencing influences expression of BCL2 but not CASP8 in MCF-7 and MDA-MB-231 cell lines [J]. Asian Pac J Cancer Prey, 2012,13(11):5865-5869.
[8] 周永春,王熙才,陳艷,等.靶向敲降Sox4基因抑制宣威地區女性肺癌細胞系XWLC-05裸鼠移植瘤的生長[J].中華腫瘤雜志,2014,36(1):11-16.
[9] Lévy P, Robin H, Bertrand F, et al. Butyrate-treated colonic Caco-2 cells exhibit defective integrin-mediated signaling together with increased apoptosis and differentiation[J]. J Cell Physiol, 2003,197(3):336-347.
[10] Wick W, Grimmel C, Wild-Bode C, et al. Ezrin-dependent promotion of glioma cell clonogenicity, motility, and invasion mediated by BCL-2 and transforming growth factor-beta2[J]. J Neurosci, 2001,21(10):3360-3368.
10.3969/j.issn.1002-266X.2016.29.013
R738.3
A
1002-266X(2016)29-0039-03
2015-09-14)
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
