OsMT-1-4c基因原核表達(dá)載體的構(gòu)建、表達(dá)及重金屬抗性研究
2017-03-31 01:10:40王小蘭
廣東農(nóng)業(yè)科學(xué) 2017年1期
關(guān)鍵詞:研究
唐 偉,張 美,王小蘭
(1.廣州大學(xué)生命科學(xué)學(xué)院,廣東 廣州 510006;2.中國(guó)科學(xué)院華南植物園,廣東 廣州 510650)
OsMT-1-4c基因原核表達(dá)載體的構(gòu)建、表達(dá)及重金屬抗性研究
唐 偉1,張 美2,王小蘭1
(1.廣州大學(xué)生命科學(xué)學(xué)院,廣東 廣州 510006;2.中國(guó)科學(xué)院華南植物園,廣東 廣州 510650)
為研究水稻抗重金屬的特性,構(gòu)建了水稻金屬硫蛋白(metallothionein,MT)原核表達(dá)載體PGEX4T-3-OsMT-1-4c,并轉(zhuǎn)化到Rossetta菌株中誘導(dǎo)表達(dá),得到大小為33.643 kDa的GST-OsMT-1-4c重組蛋白。將得到的沉淀用1×PBS溶解并在低溫下超聲破碎,離心并電泳檢測(cè),然后純化得到融合蛋白。并在重金屬Cd2+脅迫下誘導(dǎo)表達(dá)重組載體和空載體菌株,發(fā)現(xiàn)能表達(dá)GST-OsMT-1-4c蛋白菌株比表達(dá)GST蛋白菌株抗重金屬的能力強(qiáng)。
水稻;金屬硫蛋白;原核表達(dá);純化;抗重金屬特性
重金屬在環(huán)境中是普遍存在的,對(duì)植物而言,重金屬分為必需元素和非必需元素,但是非必需重金屬元素和過(guò)量的必需重金屬元素對(duì)植物的生長(zhǎng)和發(fā)育都是有害的,對(duì)植物毒害作用主要表現(xiàn)在損傷細(xì)胞膜[1]、破壞線粒體和葉綠體的超微結(jié)構(gòu)[2]、減少葉綠素和抗壞血酸含量、產(chǎn)生活性氧分子、降低各種的酶活性[3],繼而阻礙植物呼吸代謝、光合作用[4]和細(xì)胞分裂等生命活動(dòng)的正常進(jìn)行。植物生長(zhǎng)在重金屬污染的環(huán)境中[5],受重金屬的脅迫,植物與重金屬接觸的界面首先受到影響,并且這種影響隨著脅迫時(shí)間的延長(zhǎng),隨重金屬濃度的升高,植物的受害亦加劇。植物受重金屬的傷害程度、傷害癥狀與重金屬的種類(lèi)也有密切關(guān)系。重金屬能夠通過(guò)農(nóng)作物直接參與食物鏈循環(huán)[6],最終在人體內(nèi)富集[7],使人體健康受到危害。水稻是我國(guó)種植面積最大的糧食作物,稻米的產(chǎn)量、品質(zhì)及其安全問(wèn)題一直是社會(huì)關(guān)注的焦點(diǎn)。當(dāng)重金屬進(jìn)入水稻體內(nèi)后,大部分會(huì)積累在水稻根部[8],很少一部分向地上組織遷移,積累在主莖稻穗、葉片以及籽實(shí)中[9]。水稻籽實(shí)不同部位中的重金屬含量較不均勻,胚乳中總量最高[10]。據(jù)報(bào)道,我國(guó)華南地區(qū)稻米重金屬含量符合標(biāo)準(zhǔn)的只有64.8%[11]。因此,研究水稻體內(nèi)響應(yīng)金屬脅迫的關(guān)鍵基因,揭示其對(duì)重金屬離子的吸收、轉(zhuǎn)運(yùn)和解毒方面的分子機(jī)制[12],對(duì)水稻的逆境育種、提高稻米的品質(zhì)和解決食品安全問(wèn)題有重大意義。
金屬硫蛋白(metallothionein,MT)的研究是當(dāng)今生物化學(xué)和分子生物學(xué)的熱點(diǎn)之一,研究表明金屬硫蛋白是一類(lèi)富含半胱氨酸,能結(jié)合重金屬的低分子量的蛋白。其主要功能是與重金屬離子結(jié)合,一般通過(guò)半胱氨酸殘基與金屬離子結(jié)合成螯合物從而減輕或消減重金屬的毒害作用[13]。此外MT 具有清除活性氧(reactive oxygen species,ROS)的功能在抗衰老、抗氧化壓力及細(xì)胞凋亡等過(guò)程中發(fā)揮著重要作用,且MT是一種潛在的細(xì)胞凋亡負(fù)調(diào)控因子,在化療中對(duì)正常機(jī)體細(xì)胞有一定的保護(hù)作用[14],但其作用機(jī)理仍不清楚。由于植物體內(nèi)MT的含量很低、分子量極小、富含的半胱氨酸殘基極易被氧化,導(dǎo)致對(duì)植物MT的研究主要停留在轉(zhuǎn)錄水平上,而對(duì)其功能的研究相對(duì)較少。有研究報(bào)道MT具有耐受重金屬Cd的特性[15],MT的大量表達(dá)能顯著提高植物耐受性。當(dāng)今對(duì)水稻金屬硫蛋白基因家族的研究較多,但是對(duì)OsMT-1-4c基因研究相對(duì)較少,現(xiàn)有的文獻(xiàn)還沒(méi)有純化得到其蛋白,也沒(méi)有對(duì)其重金屬特性做初步研究,因此本實(shí)驗(yàn)室從水稻cDNA酵母表達(dá)文庫(kù)[16]中分離得到了OsMT-1-4c基因(http://rice.plantbiology.msu.edu/, LOC_ Os01g10400),并將其構(gòu)建到PYES260載體上保存,其基因大小為237 bp,編碼78個(gè)氨基酸,其半胱氨酸含量達(dá)到16.7 %,從而推測(cè)其具有較強(qiáng)的抗重金屬能力。本研究構(gòu)建了水稻金屬硫蛋白重組表達(dá)質(zhì)粒PGEX4T-3-OsMT-1-4c,轉(zhuǎn)入Rossetta菌株中誘導(dǎo)表達(dá)重組蛋白,優(yōu)化誘導(dǎo)條件后,產(chǎn)生大量可溶的谷胱甘肽巰基轉(zhuǎn)移酶融合蛋白(GST-OsMT-1-4c),并純化得到了該重組蛋白。通過(guò)在重金屬脅迫下產(chǎn)生重組蛋白的菌株與GST的菌株其生長(zhǎng)曲線相比較,發(fā)現(xiàn)產(chǎn)生重組蛋白的菌株比GST蛋白菌株生長(zhǎng)狀況好很多,表明OsMT-1-4c蛋白在Rossetta菌株中的表達(dá)提高了其抗重金屬的能力,從而初步驗(yàn)證了OsMT-1-4c蛋白具有抗重金屬能力。
1 材料與方法
1.1 試驗(yàn)材料
原核表達(dá)載體PGEX4T-3、重組質(zhì)粒PYES260和大腸桿菌DH5α由本實(shí)驗(yàn)室保存;限制性內(nèi)切酶BamHⅠ、XhoI購(gòu)自TaKaRa公司; IPTG購(gòu)自艾基生物技術(shù)有限公司、膠回收試劑盒和質(zhì)粒提取試劑盒購(gòu)自Magen公司、GST Bind Resin和填料柱購(gòu)自Novagen公司、引物合成及序列測(cè)定由上海英駿生物技術(shù)有限公司提供。
LB培養(yǎng)基配方:胰蛋白胨(Tryptone)10 g/L,酵母提取物(Yeast extract)5 g/L,氯化鈉(NaCl)10 g/L,用2.5 mol/LNaOH調(diào)節(jié)培養(yǎng)基的pH,使其達(dá)到7.4,去離子水定容至1 L,即配成液體LB培養(yǎng)基。固體培養(yǎng)基100 mL添加1.5 g瓊脂粉,然后120℃、20 min滅菌,待LB培養(yǎng)基不燙手后加入氨芐青霉素,至終濃度100 μg/mL,倒平板,凝固后放入4℃ 冰箱保存?zhèn)溆谩?/p>
2×YT培養(yǎng)基配方:胰蛋白胨(Tryptone)10 g/L,酵母提取物(Yeast extract)5 g/L,氯化鈉(NaCl)10 g/L,用2.5 mol/L NaOH調(diào)節(jié)培養(yǎng)基的pH,使其達(dá)到7.2,去離子水定容至1 L,即配成液體2×YT培養(yǎng)基。固體培養(yǎng)基100 mL添加1.5 g瓊脂粉,然后120℃ 、20 min滅菌,冷卻后室溫保存?zhèn)溆谩?/p>
SDS-PAGE所用試劑:30%丙烯酰胺混合液(Acr∶Bis為29∶1):將60 mL水預(yù)熱至37 ℃,加入丙烯酰胺(Ars)29 g及甲叉丙烯酰胺(Bis)1.0 g,溶劑定容至100 mL,并用0.45微米孔徑濾紙過(guò)濾,裝入棕色瓶中4℃ 保存;1.5 mol/L Tris-HCl緩沖液:取1 mol/L HCl溶液48 mL、三羥基甲烷(Tris)36.6 g,加雙蒸至90 mL,調(diào)pH至8.8,然后定容至100 mL,至于玻璃瓶中保存,室溫貯存; 5×電泳緩沖液:取15.1 g Tris 和94 g甘氨酸溶于800 mL去離子水中,再加入5 g SDS,調(diào)節(jié)pH至8.3,定容至1 L;10%過(guò)硫酸銨(AP):過(guò)硫酸銨0.2 g于2.0 mL EP管中,定容至2 mL,現(xiàn)用現(xiàn)配;10%SDS(十二烷基磺酸鈉):取10 g SDS加去離子水80 mL,68℃水浴加熱溶解定容至100 mL,室溫保存。
GST-OsMT-1-4c純化所需試劑:結(jié)合液:Na2HPO4·12H2O 2.9 g,KH2PO40.2 g,NaCl 8.0 g,KCl 0.2 g,加 0.8 L 去離子水溶解,調(diào)pH 7.3,去離子水定容至 1 L,4℃保存;洗脫液:Tris 堿0.605 g,還原型谷胱甘肽(reduced glutathione)0.307 g,加 80 mL去離子水溶解,調(diào) pH 8.0,去離子水定容 100 mL,-20℃保存。
1.2 重組質(zhì)粒PGEX4T-3-OsMT-1-4c的構(gòu)建
將含有OsMT-1-4c基因序列的質(zhì)粒PYES260作為PCR的模板,通過(guò)網(wǎng)站 Convert Primers Into In-Fusion? Primers(http://bioinfo. clontech.com/infusion/convertPcrPrimersInit.do)設(shè)計(jì)引物如下:
OsMT-1-4c-PEF:CTGGTTCCGCGTGGATCCAT GTCTTGCTGCGGAGGAAG
OsMT-1-4c-PER:ACGATGCGGCCGCTCGAGTT AGCAGTTGCAGGGATTGC
PCR反應(yīng)體系(50μL):10×Buffer 5.0μL,dNTP4.0μL,Taq DNA聚合酶0.5 μL,模板1μL,引物4μL,H2O 35.5μL。膠回收純化的目的片段連接到PGEX4T-3的載體(已由NcoI和BamHI雙酶切)上,轉(zhuǎn)入大腸桿菌DH5α,菌落PCR鑒定,并將鑒定正確的PGEX4T-3-OsMT-1-4c保存菌種。
1.3 GST-OsMT-1-4c誘導(dǎo)表達(dá)
將測(cè)序正確的PGEX4T-3-OsMT-1-4c質(zhì)粒轉(zhuǎn)化到Rossetta感受態(tài)細(xì)胞,挑取單菌落進(jìn)行PCR鑒定陽(yáng)性克隆。陽(yáng)性單克隆接種于2 mL含100μg/mL 氨芐青霉素 (Amp)、50μg/mL卡那霉素(Kam)、34μg/mL氯霉素(Chl)的 LB培養(yǎng)基上,37℃ 200 r/min過(guò)夜培養(yǎng),按體積比1∶100轉(zhuǎn)接到100 mL LB(含AmpKanChl)新鮮2×YT培養(yǎng)基,相同條件下培養(yǎng),當(dāng)OD600值到0.4~0.6,加異丙基硫代半乳糖苷(IPTG)至終濃度0.2 mmol/L,誘導(dǎo)表達(dá),收集菌液離心,并用SDSPAGE凝膠電泳分析誘導(dǎo)表達(dá)結(jié)果。
1.4 GST-OsMT-1-4c蛋白純化
通過(guò)SDS-PAGE電泳檢測(cè)融合蛋白已經(jīng)大量表達(dá),收集菌體,沉淀用1×PBS懸浮,超聲破粹30 min(9 s開(kāi),9 s關(guān))離心50 min(4℃,8 000 r/min),收集上清液和沉淀,SDSPAGE檢測(cè)融合蛋白在上清液還是沉淀中,如果在上清液中,則利用有GST標(biāo)簽的層析柱進(jìn)行蛋白純化。
1.5 重金屬抗性分析
1.5.1 適合的金屬離子濃度確定 通過(guò)分光光度計(jì)測(cè)定重金屬脅迫下重組大腸桿菌的生長(zhǎng)狀況,并將PGEX4T-3的空載體作為對(duì)照,比較表達(dá)OsMT蛋白的大腸桿菌Rossetta耐受重金屬離子鎘的能力[17]。前期實(shí)驗(yàn)確定了Rossetta菌對(duì)Cd2+耐受范圍,因此對(duì)Cd2+通過(guò)0、0.8、1.0、1.2 mmol/L濃度梯度試驗(yàn),重復(fù)3次,確定金屬離子合適的濃度。誘導(dǎo)方法如前所述,將加入IPTG和重金屬離子定為0 h,每隔1 h測(cè)定1次OD600值,以O(shè)D600為縱軸、時(shí)間為橫軸作圖,確定對(duì)細(xì)菌生長(zhǎng)和增殖有一定抑制作用,且未使細(xì)菌全部死亡的濃度范圍。
1.5.2 表達(dá)GST-OsMT-1-4c與GST蛋白的Rossetta菌對(duì)重金屬抗性比較 用吸光度法研究表達(dá)OsMT的大腸桿菌Rossetta細(xì)胞對(duì)金屬離子的耐受性,并將空載體PGEXT4T-3菌株作對(duì)照,重復(fù)3次。在Rossetta菌液中加入IPTG和合適的金屬離子濃度誘導(dǎo),繼續(xù)培養(yǎng)并每隔1 h測(cè)定一次OD600值,以O(shè)D600為縱軸作圖。
2 結(jié)果與分析
2.1 PGEX4T-3-OsMT-1-4c載體的構(gòu)建
根據(jù)引物所在位置,PCR的大小應(yīng)為 237 bp,電泳檢測(cè)結(jié)果顯示在200~400 bp位置有條帶(圖1),說(shuō)明OsMT-1-4c基因已經(jīng)克隆成功,然后In-fusion連接并轉(zhuǎn)化DH5α菌株,37℃培養(yǎng),挑取單菌落進(jìn)行菌落PCR鑒定,結(jié)果顯示在200~400 bp位置有條帶(圖2)。再對(duì)該單菌落提取質(zhì)粒測(cè)序,確定OsMT已經(jīng)正確插入載體PGEX4T-3中,表明已經(jīng)成功構(gòu)建了PGEX4T-3-OsMT-1-4c的表達(dá)載體。

圖1 PCR擴(kuò)增 OsMT cDNA電泳結(jié)果

圖2 PGEX4T-3-OsMT-1-4c菌液 PCR電泳結(jié)果
2.2 GST-0sM-1-4c蛋白誘導(dǎo)表達(dá)
將轉(zhuǎn)入鑒定正確的載體菌株誘導(dǎo)表達(dá),設(shè)收集菌體,SDS-PAGE凝膠電泳結(jié)果顯示在37℃、3 h和0.2 mmol/L IPTG時(shí)蛋白誘導(dǎo)表達(dá)最大(圖3)。將誘導(dǎo)表達(dá)最大的條件大量誘導(dǎo),收集菌體,超聲破碎收集上清和沉淀,發(fā)現(xiàn)融合蛋白基本在上清液中,將上清液低溫保存?zhèn)溆谩?/p>
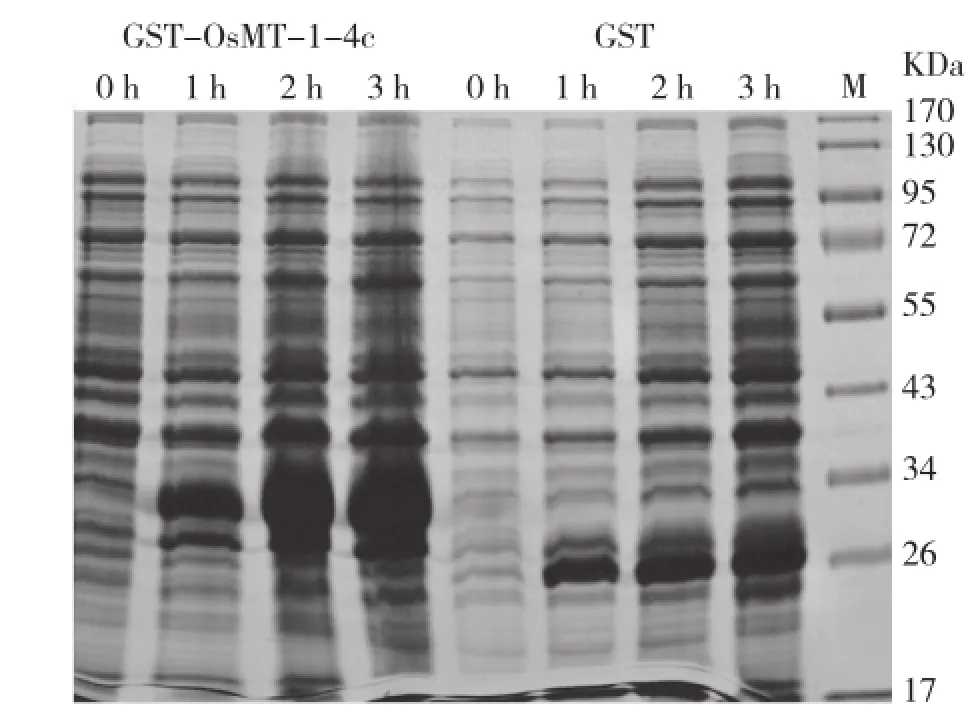
圖3 GST-OsMT-1-4C和GST蛋白誘導(dǎo)表達(dá)
2.3 GST-0sM-1-4c蛋白純化
將大量表達(dá)蛋白的上清液進(jìn)行GST親和層析柱純化,得到蛋白洗脫液,再取少量洗脫液進(jìn)行SDS-PAGE電泳分析,在26~34 kDa位置只有一條很粗的條帶,GST大小為26 kDa,OsMT-1-4c蛋白大小為8 kDa,表明得到了純化的融合蛋白GST-0sM-1-4c(圖4)。

圖4 GST-OsMT-1-4c蛋白純化
2.4 重金屬抗性分析
2.4.1 適合的金屬離子濃度確定 選用Cd2+作為研究的金屬離子[18],方法如前所述,每小時(shí)測(cè)定一次OD600值,重復(fù)3次,以時(shí)間為橫軸作圖(圖5),確定對(duì)Rossetta菌的生長(zhǎng)和增殖有一定抑制作用,又未使細(xì)菌全部死亡的濃度范圍。通過(guò)比較4個(gè)濃度梯度細(xì)菌的生長(zhǎng)狀況發(fā)現(xiàn),1.0 mmol/L濃度梯度下細(xì)菌的生長(zhǎng)曲線斜率接近1,即在此濃度下對(duì)Rossetta菌的生長(zhǎng)和增殖有一定抑制作用,又不至于死亡。從而確定實(shí)驗(yàn)合適濃度是1.0 mmol/L CdCl2。為了確定重組菌與對(duì)照菌(轉(zhuǎn)入空載體PGEX4T-3的Rossetta菌)之間,在沒(méi)有任何處理的情況下其生長(zhǎng)狀況是否一致,按上述方法培養(yǎng)大腸桿菌,結(jié)果顯示當(dāng)培養(yǎng)液不添加金屬離子時(shí),對(duì)照菌與重組菌生長(zhǎng)狀況基本相同,為進(jìn)一步的實(shí)驗(yàn)排除干擾。

圖5 Cd2+對(duì)含GST基因的Rossetta菌生長(zhǎng)狀況的影響
2.4.2 表達(dá)GST-OsMT-1-4c 與GST蛋白的Rossetta菌對(duì)重金屬抗性比較 如圖6所示,1.0 mmol/L CdCl2處理3 h內(nèi),對(duì)照菌和重組菌增長(zhǎng)沒(méi)有明顯差異;處理3 h后,表達(dá)OsMT蛋白的重組菌增長(zhǎng)率明顯高于對(duì)照菌。說(shuō)明表達(dá)OsMT融合蛋白確實(shí)增強(qiáng)了Rossetta菌對(duì)Cd離子的耐受性,提高了細(xì)菌的存活率,從而使相應(yīng)的重組菌表現(xiàn)出耐受重金屬離子的能力。

圖6 Cd2+對(duì)表達(dá)GST和GST-MT-1-4c蛋白的Rossetta菌生長(zhǎng)狀況的影響
3 結(jié)論與討論
金屬硫蛋白是一類(lèi)富含半胱氨酸殘基的低分子量蛋白,能通過(guò)半胱氨酸殘基上的巰基基團(tuán)與金屬離子結(jié)合,從而減輕重金屬離子對(duì)植物體的危害。目前,對(duì)植物MT蛋白的研究很多,如在蠶豆中對(duì)MT基因的表達(dá)分析認(rèn)為[19],MT能與金屬離子直接結(jié)合[20],穩(wěn)定植物內(nèi)環(huán)境中金屬離子的含量,并能有效的轉(zhuǎn)運(yùn)金屬離子到有需要的組織,從而說(shuō)明了植物MT基因參與了金屬離子的運(yùn)輸?shù)墓δ堋T跀M南芥中利用轉(zhuǎn)MT-Ⅱ基因的超表達(dá)植株,分析表明,轉(zhuǎn)基因擬南芥能有效維持體內(nèi)氧自由基的含量,減少M(fèi)DA的產(chǎn)生,從而緩解干旱脅迫引起的傷害,提高抗旱性[21]等。金屬硫蛋白基因豐富的巰基含量及重金屬結(jié)合的能力決定了功能的多樣性。目前雖然對(duì)抗重金屬方面研究不斷深入,但是對(duì)其他功能尚不清楚,加上已知的植物MT蛋白較少,進(jìn)一步阻礙了對(duì)MT功能的研究。因此,水稻金屬硫蛋白表達(dá)載體的構(gòu)建及抗重金屬特性初步研究,將有助于進(jìn)一步研究植物MT的功能。
由于金屬硫蛋白在植物體內(nèi)含量較少,分子量極低,且半胱氨酸極易被氧化,因此很難檢測(cè)其在生物體內(nèi)的表達(dá)活性。本研究的OsMT-1-4c蛋白僅為7.643 kDa左右,在大腸桿菌中表達(dá)很難直接檢測(cè)。因此,本研究選用含有GST標(biāo)簽的蛋白表達(dá)載體PGEX4T-3,其GST大小為26 kDa,是根據(jù)唐青藍(lán)等[22]用PGEX4T-3成功構(gòu)建了原核表達(dá)載體,并與王娟等[23]選擇PGEX系列構(gòu)建海州香薷金屬硫蛋白重組表達(dá)質(zhì)粒PGEX-2T-Eh MT1相類(lèi)似,為大量表達(dá)和純化重組蛋白GST-OsMT-1-4c提供依據(jù),為在蛋白水平上研究MT蛋白打下堅(jiān)實(shí)的基礎(chǔ)。表達(dá)載體的宿主菌為 Rossetta,是一種廣泛使用的表達(dá)宿主菌,適合于 GST 表達(dá)體系,OsMT-1-4c蛋白在此宿主菌中活性穩(wěn)定且產(chǎn)量高[24]。在IPTG誘導(dǎo)3 h后,目的蛋白得到大量表達(dá),然后純化得到GST-OsMT-1-4c重組蛋白。重金屬抗性分析是通過(guò)重金屬Cd2+脅迫含目的基因的菌株,通過(guò)目的蛋白的表達(dá),比較表達(dá)GSTOsMT-1-4c蛋白與GST蛋白的菌株的生長(zhǎng)狀況,結(jié)果表明1.0 mmol/L CdCl2處理3 h內(nèi),對(duì)照菌和重組菌增長(zhǎng)沒(méi)有明顯差異;處理3 h后,表達(dá)OsMT蛋白的重組菌增長(zhǎng)率明顯高于對(duì)照菌。說(shuō)明OsMT-1-4c可能與金屬離子相結(jié)合,從而使相應(yīng)的重組菌表現(xiàn)出耐受金屬離子的能力。通過(guò)本實(shí)驗(yàn)OsMT-1-4c表達(dá),將為提高對(duì)重金屬離子的耐受和毒性提供一個(gè)可行的基因策略,為今后在利用生物體抗重金屬修復(fù)上提供應(yīng)用前景[25]。
[1] Danielle L,Leonardo T. Lead poisoning:Successes and 21stcentury challenges[J]. Pediatrive in Review,2005,26(12):435-443.
[2] Stephan C. Toxic metal accumulation,responses to exposure and mechanisms of tolerance in plants[J]. Biochimie,2006,88(11):1707-1719.
[3] 梁琪惠,吳永勝,劉剛,等. Cr、As、Pb、Cd 復(fù)合污染對(duì)茶樹(shù)葉片酶活性和細(xì)胞膜透性的影響[J]. 南方農(nóng)業(yè),2012(7):1-6.
[4] 梁文斌,薛生國(guó),沈吉紅,等. 錳脅迫對(duì)垂序商陸葉片形態(tài)結(jié)構(gòu)及葉綠體超微結(jié)構(gòu)的影響[J]. 生態(tài)學(xué)報(bào),2011,31(13):3677-3683.
[5] 劉候俊,韓曉日,李軍,等. 土壤重金屬污染現(xiàn)狀與修復(fù)[J]. 環(huán)境保護(hù)與循環(huán)經(jīng)濟(jì),2012(7):4-8.
[6] 徐小遜,張世熔,解姍姍,等. 垂序商陸對(duì) Cd 脅迫的生長(zhǎng)響應(yīng)和富集特征研究[J]. 西南農(nóng)業(yè)學(xué)報(bào),2012,25(4):1358-1362.
[7] 王義,黃先飛,胡繼偉,等. 重金屬污染與修復(fù)研究進(jìn)展[J]. 河南農(nóng)業(yè)科學(xué)2012,41(4):1-6.
[8] 孔令芳,朱紅瓊,楊曉霞. 重金屬鉛和汞對(duì)蠶豆根尖細(xì)胞的微核效應(yīng)[J]. 大理學(xué)院學(xué)報(bào),2011,10(4):51-53.
[9] 李榮春. Pb、Cd及其復(fù)合污染對(duì)烤煙葉片生理生化及細(xì)胞亞顯微結(jié)構(gòu)的影響[J]. 植物生態(tài)學(xué)報(bào),2000,22(4):238- 242.
[10] 解凱彬,施國(guó)新,陳國(guó)祥,等. Hg2+對(duì)萍蓬草光合膜超微結(jié)構(gòu)及功能的影響[J]. 農(nóng)村生態(tài)環(huán)境,2002,18(1):26- 30.
[11] Bianka S,Margret S. Epidermal cell death in rice is confined to cells with a distinct molecular identity and is mediated by ethylene and H2O2through an autoamplified signal pathway[J]. Plant Cell,2009,21(1):184-196.
[12] 陳春,周啟星. 金屬硫蛋白作為重金屬污染生物標(biāo)志物的研究進(jìn)展[J]. 農(nóng)業(yè)環(huán)境科學(xué)學(xué)報(bào),2009,28(3):425-432.
[13] Gupreet K,Harminder P S,Daizy R B,et al. Lead (Pb)-induced biochemical and ultrastructural changes in wheat(Triticum aestivum)roots[J]. Protoplasma,2013,250(1):53-62.
[14] 楊梅,程安春,汪銘書(shū),等. 重組鴨α-干擾素基因工程菌表達(dá)條件的研究[J]. 四川農(nóng)業(yè)大學(xué)學(xué)報(bào),2007,25(3):337-342.
[15] 王晶,高燕,李靜,等. 水稻cDNA酵母表達(dá)文庫(kù)的構(gòu)建及重金屬鎘脅迫相關(guān)基因的初步篩選[J]. 廣東農(nóng)業(yè)科學(xué),2012,39(13):143-145.
[16] 張紅曉,王世華,賀春玲,等. 水稻金屬硫蛋白ricMT基因的耐受性分析[J]. 廣東農(nóng)業(yè)科學(xué),2011,38(24):117-119.
[17] Li Y J,Wu H,Wei X X,et al. Subcellular distribution of Cd and Zn and MT mRNAexpression in the hepatopancreas of Sinopotamon henanense after single and co-exposure to Cd and Zn[J]. Comparative Biochemistry and Physiology(Part C):Toxicology & Pharmacology,2015,167:117-130.
[18] Rhonda C F,Liang Z M,Karam B S. Analysis of type 1 metallothionein c DNAs in Vicia faba[J]. Plant Mol Biol,1997,33(4):583-591.
[19] Kayo Y,Hitoshi N,Rie T,et al. Effects of soy protein isolate on LEC rats,a model of Wilson disease:mechanisms underlying enhancement of liver cell damage[J]. Biochemical and Biophysical Research Communications,2003,303:271-274.
[20] 唐中華,郭曉瑞,張洋洋,等. 擬南芥 MT-II 過(guò)量表達(dá)提高抗旱性[J]. 植物研究,2005,25(4):415-418.
[21] 唐青藍(lán),許慶忠,張禮林,等. pGE-4T-3-MFGF21表達(dá)載體的構(gòu)建、表達(dá)與純化[J]. 生物技術(shù),2014,24(2):1-24.
[22] 王娟,王桂萍,張紅生,等. 海州香薷金屬硫蛋白(Eh MT1)的原核表達(dá)和純化及其多克隆抗體的制備[J]. 南京農(nóng)業(yè)大學(xué)學(xué)報(bào),2010,33(4):25-30.
[23] Carstens C P,Waeshe A. Codon bias-adjusted BL21 derivatives for protein expression[J]. Strategies,1999,12:49 -51.
[24] 何永吉,劉進(jìn)平,李穎君,等. 華溪蟹功能性重組金屬硫蛋白的表達(dá)及抗血清制備[J]. 環(huán)境科學(xué)學(xué)報(bào),2014,34(9):2434-2439.
(責(zé)任編輯 楊賢智)
Vector construction, prokaryotic expression and resistance to heavy metals of OsMT-1-4c
TANG Wei1,ZHANG Mei2,WANG Xiao-lan1
(1.College of Life Science, Guangzhou University,Guangzhou 510006,China;
2.South China Botanical Garden, Chinese Academy of Sciences,Guangzhou 510650,China)
In order to study the resistance of paddy rice to heavy metals, PGEX4T-3-OsMT-1-4c, the prokaryotic expression vector of metallothione for paddy rice was constructed, which was then transformed into strain Rossetta to induce the expression, about 33.643 kDa of GST-OsMT-1-4c fusion protein was obtained. The obtained precipitation was dissolved with 1×PBS and conducted ultrasonic fragmentation at low temperature,followed by the centrifugation and electrophoresis detection, and then was gained through the purification. Furthermore, when the recombinant vector and empty vector strains were induced to express under the stress of heavy metal Cd2+, it was found that the strain to express protein GST-OsMT-1-4c owned stronger resistance to heavy metals than that of protein GST.
rice;metallothionein; prokaryotic expression; purification;resistance to heavy metals
X592
A
1004-874X(2017)01-0008-07
2016-11-01
國(guó)家自然科學(xué)基金(30971912)
唐偉(1989-),男,在讀碩士生,E-mail:907264698@qq.com
王小蘭(1973-),女,博士,教授,E-mail:1372848641@qq.com
唐偉,張美,王小蘭. OsMT-1-4c基因原核表達(dá)載體的構(gòu)建、表達(dá)及重金屬抗性研究[J].廣東農(nóng)業(yè)科學(xué),2017,44(1):8-14.
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國(guó)語(yǔ)大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車(chē)工程學(xué)報(bào)(2017年2期)2017-07-05 08:13:02
國(guó)際商務(wù)財(cái)會(huì)(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19

- 廣東農(nóng)業(yè)科學(xué)的其它文章
- 利用SSR標(biāo)記對(duì)秈稻品種(系)聚類(lèi)分析的比較研究
- 新技術(shù)認(rèn)知對(duì)農(nóng)戶親環(huán)境行為的影響機(jī)理研究
——以菜農(nóng)生物農(nóng)藥施用為例 - 農(nóng)業(yè)生產(chǎn)環(huán)節(jié)外包驅(qū)動(dòng)因素研究
——來(lái)自廣東省的問(wèn)卷 - 海南島橡膠風(fēng)害的重現(xiàn)期預(yù)測(cè)
- 基于信息熵的山西太谷植被指數(shù)最佳分辨率尺度分析
- 近30年珠海市耕地生態(tài)安全評(píng)價(jià)及未來(lái)預(yù)測(cè)