諸氏鯔蝦虎魚幼魚飼料纖維素適宜含量的初步研究
2017-03-31 01:11:07魏遠征林忠婷李建軍
廣東農業科學 2017年1期
魏遠征,林忠婷,李建軍
(廣東省實驗動物監測所/廣東省實驗動物重點實驗室,廣東 廣州 510663)
諸氏鯔蝦虎魚幼魚飼料纖維素適宜含量的初步研究
魏遠征,林忠婷,李建軍
(廣東省實驗動物監測所/廣東省實驗動物重點實驗室,廣東 廣州 510663)
為探討纖維素含量對諸氏鯔蝦虎魚幼魚生長的影響,配制了6種不同纖維素含量的配合飼料,飼喂60日齡幼魚8周,測定存活率(SR)、增重率(WG)、特定生長率(SGR)、攝食率(FR)、飼料系數(FCR)和肥滿度(FUL),切片觀察各組蝦虎魚的腸道,并測定魚體基本組分(粗蛋白、粗脂肪、水分和灰分)。結果表明,幼魚SR隨纖維素含量的增加逐漸下降;WG和SGR先上升后下降;FR持續上升,但組間無顯著差異;飼料纖維素含量超過2.98%時,幼魚FUL持續下降;纖維素含量由14.52%增加到20.63%時,FCR極顯著增大。同時纖維素含量影響蝦虎魚幼魚腸道形態,并對魚體粗蛋白、粗脂肪含量有顯著影響,但對水分、灰分均無顯著影響。諸氏鯔蝦虎魚幼魚飼料最適纖維素含量為6.74%。
諸氏鯔蝦虎魚;纖維素;生長;飼料利用
諸氏鯔蝦虎魚(Mugilogobius chulae)是一種廣泛分布于香港、菲律賓、日本、泰國及西太平洋海域的暖水性底層小型海水魚類,屬鱸形目(Perciformes)、蝦虎魚亞目(Gobioide)、蝦虎魚科(Gobiidae)、鯔蝦虎魚屬(Mugilogobius)[1]。該魚具有個體小、性成熟時間短、繁殖力強、周年產卵、對有毒有害物質敏感等諸多實驗動物開發優勢,已廣泛應用于海洋環境毒理學研究等領域[2-3]。
研究實驗魚的營養和飼料質量控制是實現實驗動物標準化的條件之一,而纖維素作為飼料中的固有成分,是飼料質量控制的重要組成部分。適宜含量的纖維素能促進腸道的蠕動,同時可作為其他營養素的稀釋劑和擴充劑,增大與消化酶的接觸面積,提高消化率,進而促進魚體對營養物質的吸收[4]。但過高含量的纖維素會抑制魚的生長[5-6]。目前飼料纖維素適宜含量的研究多集中在具有經濟價值的草食和雜食性魚類,如草魚(Ctenopharyngodon idella)[7]、羅非魚(Oreochromis mossambicus)[5]、虹鱒(Salmo gairdneri)[6]等,對肉食性魚類的研究非常有限。同時小型實驗魚類由于自身的特點,飼料開發難度較大,極少有針對纖維素適宜含量的研究。斑馬魚(Danio rerio)幼魚商品飼料中纖維素含量通常不超過2%[8-9],劍尾魚(Xiphophorus helleri)為5%[10]。由于適宜的纖維素含量受魚種類、發育階段等多種因素影響[11-12],且國內外尚無蝦虎魚的相關報道,因此飼料質量控制中需要開展諸氏鯔蝦虎魚適宜纖維素含量的研究。
本試驗在前期營養需求研究(粗蛋白、粗脂肪等)的基礎上,初步探討飼料纖維素含量對蝦虎魚幼魚生長性能和飼料利用的影響,以期為諸氏鯔蝦虎魚專用配合飼料的開發及相關標準的制定提供參考。
1 材料與方法
1.1 實驗魚和條件控制
以60日齡諸氏鯔蝦虎魚幼魚進行飼喂試驗,實驗魚為引種馴化的封閉群子9代(野生種群來源深圳大鵬灣)。試驗開始前,每天投喂等量的鹵蟲和混合實驗飼料,集中馴養1周。試驗時集中禁食24 h,挑取規格均勻、健壯的幼魚隨機分為6組,每組3個重復,每個重復40尾,平均體重為0.019~0.024 g。
飼養環境為室內加裝循環水系統的塑料水族箱(90 cm×45 cm×15 cm),條件控制為水溫24~26℃,鹽度22 ‰~26 ‰,pH≈7.5,氨氮 ≤ 0.5 mg/L,光周期12 h∶12 h。
1.2 飼料制備與投喂
以微晶纖維素作為飼料纖維素來源,添加量分別為0%、1%、4%、8%、16%、26%;白魚粉和豆粕為蛋白質源;魚油和豆油為脂肪源,分別配制相同粗蛋白、粗脂肪含量的6種飼料。所有飼料原料過孔徑380 μm篩網后,制粒機(XL,常州市智陽機械設備有限公司)制粒。濕顆粒飼料烘箱70℃干燥后,密封分裝于4℃保存備用。依據不同生長期蝦虎魚的口徑,手工碾磨并過篩處理,以獲得最適粒徑的飼料(60~ 90日齡:0.3~0.5 mm;90~120日齡:0.6~0.8 mm)。
以上述0%微晶纖維素作為對照,其余5種配合飼料編號依次為Ⅰ~Ⅴ,飼料原料與基本生化組成見表1。投喂時間為每天9:00和16:00,投喂量以實驗魚20 min內吃完為宜。試驗結束時,稱5 g飼料放入實驗缸中,10 min后回收,70℃烘干稱重,計算飼料的溶失率以校正實際攝食量。每天吸污并補充定量海水,測量水質并觀察魚活動情況。

表1 飼料原料及基本成分(%)分析
1.3 幼魚生長性能和飼料利用效果測定
8周生長實驗結束時,計算實驗魚存活率(SR)、增重率(WG)、特定生長率(SGR)、攝食率(FR)、飼料系數(FCR)和肥滿度(FUL):

采用SPSS軟件進行單因素方差(One-way ANOVA)分析,計算不同飼料投喂對蝦虎魚生長的影響,差異顯著時進行Duncan’s 多重比較。
1.4 飼料與魚體成分的測定
每組取150 g飼料樣品,參照GB 5009.5-2010、GB 5009.6-2003測定其粗蛋白、粗脂肪含量。粗蛋白含量采用凱氏定氮法,使用全自動定氮儀蒸餾裝置(KDN-812,上海纖檢儀器有限公司)測定樣品含氮量,再乘以6.25;粗脂肪含量用乙醚溶劑,采用索氏抽提法進行提取。每個樣品重復測定3次。飼料中的纖維素含量參照GB/T 5009.88-2008方法,采用酶解法進行測定。
試驗結束后,禁食24 h,對各組蝦虎魚計數并稱重,隨機取其中15尾進行體長和全長的測定。測定結束后每組隨機取20尾魚麻醉處理(MS222,Sigma USA),勻漿后進行生化成分測定。全魚水分、灰分、粗蛋白、粗脂肪成分的測定參照GB/T18654.10-2008的方法。其中樣品水分含量在105℃烘箱中烘至恒重進行測定,灰分含量于馬弗爐中550℃充分灼燒測定,其他成分測定方法與飼料測定方法相同。
1.5 組織切片的制備
8周生長實驗結束時,各組取3尾蝦虎魚禁食24 h,于10% 中性福爾馬林固定液中保存。常規石蠟包埋、固定后,切除頭部和尾部,采用自動切片機連續切片(厚度5 μm),蘇木精-伊紅(HE)染色,中性樹膠封片,Leica 顯微鏡觀察并拍照。
2 結果與分析
2.1 幼魚生長性能和飼料利用效果
8周生長實驗結束時,60日齡諸氏鯔蝦虎魚幼魚體重由0.019~0.024 g增加到0.128 ~0.179 g。各處理諸氏鯔蝦虎魚幼魚的SR、WG、 SGR、FR、FCR和FUL測定結果見表2。纖維素含量0.85%時,幼魚SR顯著高于其他組;隨著纖維素含量的增加,幼魚SR逐漸下降。SGR和WG均先增大后減小,其中纖維素含量由0.85%增加到2.98%時,WG顯著增大;由2.98%增加到7.23%時SGR顯著增大;超過14.52%時均顯著下降。纖維素含量為20.63%時FR最大,但各處理無顯著差異。纖維素含量為7.23%時,FCR最小;由14.52%增加到20.63%時,FCR顯著增大。FUL隨纖維素含量的增加而減小。
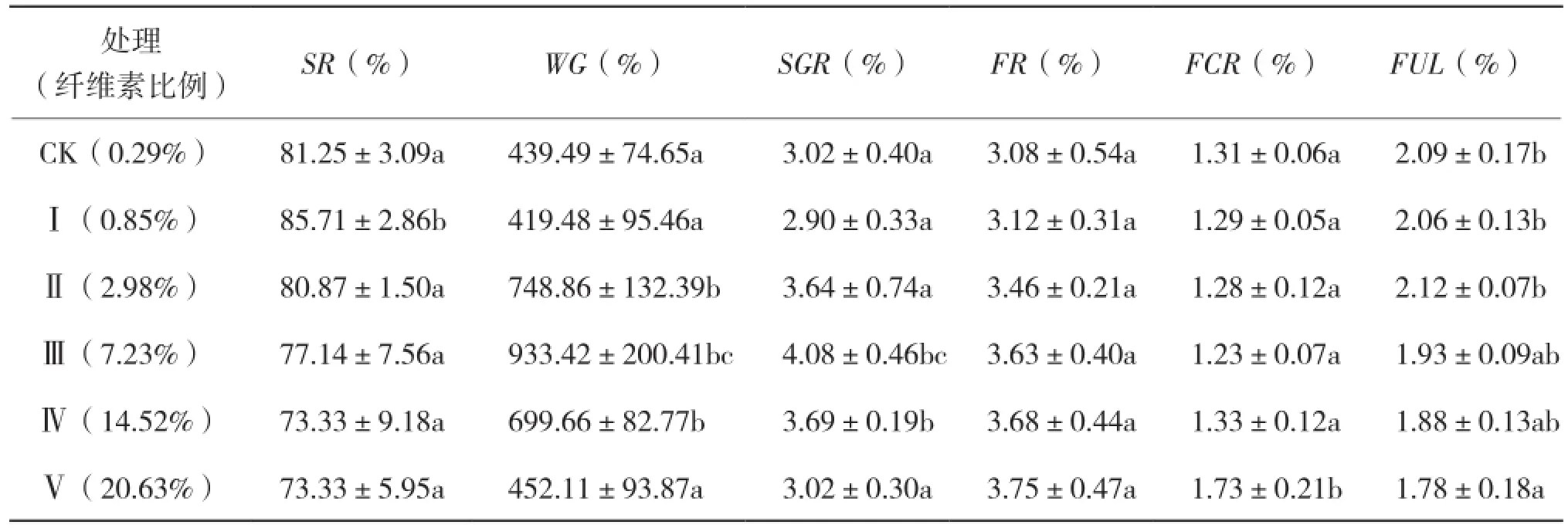
表2 飼料纖維素含量對蝦虎魚生長和飼料利用的影響
蝦虎魚幼魚的生長參數(SGR、WG)和飼料利用效果(FR、FCR)與纖維素含量的相關性分析見圖1。其中幼魚SGR和WG的回歸方程分別為:y=-0.0106x2+ 0.2203x+ 2.923(R2=0.8936)和y=-4.5661x2+ 93.291x+ 423.98(R2=0.8407),均隨飼料纖維素含量的增加呈現先上升后下降趨勢;SGR和WG最大時的纖維素含量分別為10.39%和10.22%。FR和FCR的回歸方程分別為:y=-0.0026x2+ 0.0833x+ 3.104(R2=0.9258)和y=0.0027x2- 0.0364x+ 1.3349(R2=0.9752),FR隨纖維素含量的上升而上升,FCR略下降后持續上升,且FCR最小時纖維素含量為6.74%(圖1)。
2.2 幼魚魚體組分
8周生長試驗結束時,對全魚基本組分進行生化測定,結果(表3)表明,飼料纖維素含量對幼魚全魚水分、灰分均無顯著影響;纖維素含量2.98%時,幼魚粗蛋白含量最高;粗脂肪含量隨纖維素含量的增加而下降,且由2.98%增加或超過7.23%時,粗脂肪含量顯著下降;且纖維素含量超過7.23%時,各組間粗脂肪含量均存在極顯著差異。
2.3 組織切片顯微觀察
大多數魚類的食物儲存、消化場所為前腸,吸收場所在中腸[13-14],因此8周試驗結束時,切片觀察蝦虎魚中腸(圖2,封二)。中腸腸絨毛多數表現為指狀、錐狀、齒狀,局部腸絨毛消失為平坦的腸壁;黏膜層和肌層厚度不均一,杯狀細胞豐富。隨著飼料纖維素含量的增加,多數腸絨毛由指狀逐漸過渡到錐狀、齒狀,腸壁黏膜上皮細胞高度增加,末端刷狀緣明顯且平滑;同時靠近腸壁的黏膜固有層和肌層厚度逐漸增加。此外飼料纖維素含量超過7.23%時,腸絨毛上的杯狀細胞顯著減少。
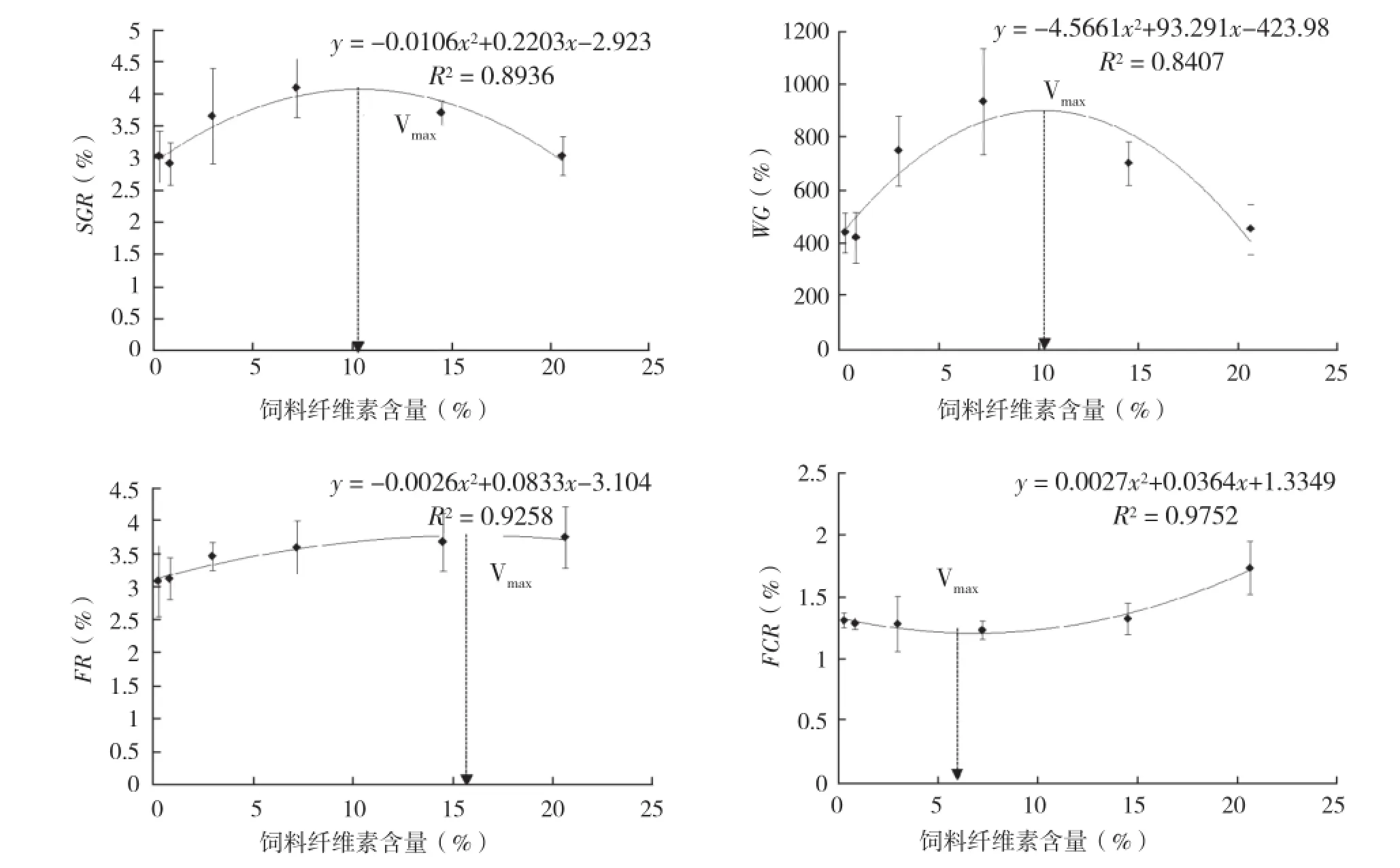
圖 1 蝦虎魚幼魚的生長參數和飼料利用效果與纖維素含量的相關性分析

表3 飼料纖維素含量對蝦虎魚全魚組分的影響
3 討論
3.1 諸氏鯔蝦虎魚的食性
通常認為肉食性魚類飼料中最適纖維素含量為2%~8%,雜食性為8%~12%,草食性為12%~20%。例如,斑點叉尾鮰(Ictalurus punctatus)飼料中粗纖維含量通常不超過7%[15-16];飼料纖維素含量超過8%時,虹鱒(Salmo gairdneri)生長就受到抑制[6];不引起湖紅點鮭(Salvelinus namaycush)和虹鱒(Salmo gairdneri)生長抑制的α-纖維素含量為8%[11],而南亞野鯪(Labeo rohita)為16%[12]。本研究中諸氏鯔蝦虎魚飼料纖維素含量為7.23%時,有較好的生長效果;纖維素含量增加到14.52%時,其生長也未受明顯抑制,說明該魚食性介于肉食和雜食,且具較好的可塑性。這與之前的研究相同,野生裸喉新蝦虎魚(Neogobius gymnotrachelus)、巖蝦虎魚(Gobius paganellus)、鱗蝦虎魚(Lepidogobius lepidus)食物組成中蝦蟹、橈足類、端足類動物所占比例均超過80%,為典型的肉食性魚類[17-18]。介于肉食和雜食的食性改變可能與室內長期馴化有關[19]。
3.2 飼料纖維素含量對幼魚生長性能和飼料利用的影響
蝦虎魚幼魚SR隨飼料中纖維素含量的增加逐漸下降,可能與幼魚消化道發育不完善、缺乏消化纖維素相關酶有關。幼魚對飼料中纖維素含量的耐受性較成魚低,如2 g規格的尼羅羅非魚(Oreochromis niloticus)飼料中最適纖維素含量小于10%,而5 g規格的魚苗為14.14%[20]。
幼魚SGR和WG均隨纖維素含量的增加先上升后下降,回歸方程分析表明,SGR和WG最大時纖維素含量分別為10.39%和10.22%;但FCR最小時,纖維素含量為6.74%,說明幼魚生長與飼料利用效果的最適纖維素量有差異。同時纖維素含量的增加導致FR持續增加,FUL持續下降,由于FUL是一個衡量魚體能量儲備水平的粗略指標,FUL的變化可預示魚體營養狀態的改變,因此纖維素含量對幼魚生長、攝食和能量儲存產生顯著影響:纖維素含量過高會導致胃腸蠕動加快,消化酶與飼料的接觸時間變短,進而導致消化率下降;消化率的下降又會對魚體的營養攝入和積累產生影響。這與其他魚的研究結果相同,21%纖維素添加組飼喂的斑點叉尾鮰(Ictalurus punctatus)的生長速度優于未添加組[21];虹鱒(Salmo gairdneri)飼喂高纖維素含量飼料時,飼料干物質的消化率顯著下降[6];隨著飼料纖維素含量的增加,虹鱒(Oncorhynchus mykiss)對飼料的干物質消化率、蛋白和能量利用率均下降[22]。同時本研究中纖維素含量由2.98%增加到7.23%時,幼魚全魚粗脂肪含量顯著下降,且與FUL的變化趨勢相同,同樣證實纖維素含量對魚體能量儲存有影響,因此飼料適宜纖維素含量需要綜合考慮生長性能和魚體能量儲存,不足或過量均對生長不利。
3.3 飼料纖維素含量對幼魚腸道形態的影響
纖維素含量對蝦虎魚幼魚腸道形態有影響,可能是腸道黏膜對纖維素刺激的一種適應性反應,這與大多數研究結果相同[23-24]:由于纖維素不被腸消化吸收,使腸壁蠕動時承受機械壓力增大,腸黏膜厚度增加以增強對機械作用的承受能力。當飼料纖維素含量超過7.23%時,腸絨毛上杯狀細胞GC含量顯著減少,說明纖維素過多會導致消化率下降,進而影響營養物質的吸收。因此綜合考慮生長、飼料利用效果以及形態指標的結果,諸氏鯔蝦虎魚幼魚飼料中最適纖維素含量宜選用FCR回歸分析結果,為6.74%。
本研究選用微晶纖維素作為飼料主要的纖維素源,由于來源單一以及單因素梯度法自身的缺陷,本試驗存在一定的局限性。鑒于飼料碳水化合物組成的復雜性以及纖維素對飼料蛋白利用的影響,蝦虎魚配合飼料中纖維素含量的最適范圍仍需進一步研究。
[1] 伍漢霖,鐘俊生. 中國動物志硬骨魚綱鱸形目(五)蝦虎魚亞目[M]. 北京:科學出版社,2008.
[2] 李建軍,林忠婷,陳小曲,等. 四種重金屬離子對諸氏鯔蝦虎魚的單一和聯合毒性[J]. 海洋環境科學,2014(2):236-241.
[3] 李建軍,吳美慧,葉惠欣,等. 不同發育期諸氏鯔蝦虎魚對鉆井液的敏感性比較[J]. 中國比較醫學雜志,2013,23(4):48-51.
[4] Ensminger M E,Oldfield J E,Heinemann W W. Feeds and nutrition digest:formerly,feeds and nutrition-abridged[M]. 1990.
[5] Dioundick O,Stom D. Effects of dietary α-cellulose levels on the juvenile tilapia,Oreochromis mossambicus(Peters)[J]. Aquaculture,1990,91(3):311-315.
[6] Hilton J,Atkinson J,Slinger S. Effect ofincreased dietary fiber on the growth of rainbow trout(Salmo gairdneri)[J]. Canadian Journal of Fisheries and Aquatic Sciences,1983,40(1):81-85.
[7] 毛永慶,蔡發盛,林鼎. 幼鯇(草魚)對蛋白質、糖、脂肪、無機混合鹽和纖維素日需要量的研究//魚類學論文集第四輯[C].北京:科學出版社,1995:81-92.
[8] Rd S A,Garris H W,Jones W T,et al. Growth and survival of zebrafish(Danio rerio)fed different commercial and laboratory diets[J]. Zebrafish,2009,6(3):275-80.
[9] Lawrence C. The husbandry of zebrafish(Danio rerio):A review[J]. Aquaculture,2007,269(1-4):1-20.
[10] Tamaru C S,Cole B,Bailey R,et al. A manual for commercial production of the swordtail,Xiphophorus helleri[M]. Center for Tropical and Subtropical Aquaculture,2001.
[11] Poston H A. Response of lake trout and rainbow trout to dietary cellulose[R]. Center for Integrated Data Analytics,Wisconsin Science Center,1986.
[12] Ashraf M,Abbas S,Rehman M H,et al. Effect of different levels of α-cellulose on growth and survival of Rohu(Labeo Rohita)Fingerlings[J]. Global Journal of Animal Scientific Research,2014,2(4):321-326.
[13] 宋霖. 黃顙魚和中華絨螯蟹胃腸道形態和功能對投飼的響應[D]. 蘇州:蘇州大學,2013.
[14] 李貴生,周黎華,梁旭方. 3H-亮氨酸在幼草魚腸道中的吸收積累作用[J]. 水生態學雜志,2003,23(5):4-6.
[15] Lovell T. Nutrition and Feeding of Fish[J]. Aquaculture,1989,267(1-4):13-70.
[16] Robinson E H,Manning B B,Li M H. 12 Feeds and feeding practices[J]. Developments in aquaculture and fisheries science,2004,34:324-348.
[17] Grossman G D,Coffin R,Moyle P B. Feeding ecology of the bay goby(Pisces:Gobiidae). Effects of behavioral,ontogenetic,and temporal variation on diet[J]. Journal of Experimental Marine Biology and Ecology,1980,44(1):47-59.
[18] Skora K E,Rzeznik J. Observations on diet composition of neogobius melanostomus Pallas 1811(Gobiidae,Pisces)in the Gulf of Gdansk(Baltic sea)[J]. Journal of Great Lakes Research,2001,27(3):290-299.
[19] 陳乃松,肖溫溫,梁勤朗,等. 飼料中脂肪與蛋白質比對大口黑鱸生長,體組成和非特異性免疫的影響[J]. 水產學報,2012,36(8):1270-1280.
[20] 郝彥周,王瑞軍,朱衛孝. 魚類飼料中粗纖維適宜含量的研究概述[J]. 齊魯漁業,2007(9):43-45.
[21] Dupree H K,Sneed K E. Response of channel catfish fingerlings to different levels of major nutrients in purified diets[R]. Center for Integrated Data Analytics Wisconsin Science Center,1966.
[22] Glencross B. The influence of soluble and insoluble lupin non-starch polysaccharides on the digestibility of diets fed to rainbow trout(Oncorhynchus mykiss)[J]. Aquaculture,2009,294(3):256-261.
[23] Navarrete P,Fuentes P,la Fuente L,et al. Shortterm effects of dietary soybean meal and lactic acid bacteria on the intestinal morphology and microbiota of Atlantic salmon(Salmo salar)[J]. Aquaculture Nutrition,2013,19(5):827-836.
[24] 陸紅佳,張磊,劉金枝,等. 納米甘薯渣纖維素對高脂膳食大鼠腸道內環境及形態的影響[J]. 食品科學,2015(5):172-178.
(責任編輯 崔建勛)
Preliminary study on suitable cellulose content in formulated diet for juvenile Mugilogobius chulae
WEI Yuan-zheng,LIN Zhong-ting,LI Jian-jun
(Guangdong Laboratory Animals Monitoring Institute/ Key Laboratory of Guangdong Laboratory Animals,Guangzhou 510663,China)
To assess the effects of cellulose level on growth and feed efficiency of juvenile goby(Mugilogobius chulae)and provide reference for exploitation of compound feed and the relevant standards development process,six purified diets with different cellulose levels,that is CK(0.29%),Ⅰ(0.85%),Ⅱ(2.98%),Ⅲ(7.23%),Ⅳ(14.52%)and Ⅴ(20.63%)were formulated. The protein and lipid contents were kept constant in all diets. At the end of 8 weeks growth experiment,Survival rate(SR),Weight gain(WG),Specific growth rate(SGR),Feed conversion ratio(FCR),Feeding rate(FR)and Fullness(FUL)were measured. Gobys’ intestinal morphology and body compositions with different diertary cellulose levels were also measured. The results showed that juvenile fishes’ SR decreased with the rising of cellulose levels. The SR of I(0.85%)was significant different with other groups(P<0.05). All groups’ WG and SGR showed upgraded firstly and then decreased with the increase of dietary cellulose. When cellulose level increased to 7.23%,WG and SGR reached a maximum. Though the FR highly increased related to the increase of cellulose,there was no significant difference among groups. When dietary cellulose content increased to 20.63%,FCR significantly increased(P<0.05). The FUL continuously decreased when dietary cellulose exceeded 2.98%. Meanwhile,dierary cellulose content had effects on the development of goby’s intestinalmorphology. The whole body crude protein and lipid were also significantly influenced(P<0.05)by dietary cellulose,but mositure and ash were not influenced. The optimum cellulose content in diet for juvenile M. chulae is 6.74%.
Mugilogobius chulae;cellulose;growth;feed utilization efficiency
S963.1
A
1004-874X(2017)01-0149-07
2016-11-02
廣東省科技計劃項目(2014A030304033);國家科技支撐計劃項目(2015BAI09B05)
魏遠征(1987-),男,碩士,研究實習員,E-mail:yzwei87@163.com
李建軍(1972-),男,碩士,高級工程師,E-mail:ljj@gdlami.com
魏遠征,林忠婷,李建軍. 諸氏鯔蝦虎魚幼魚飼料纖維素適宜含量的初步研究[J].廣東農業科學,2017,44(1):149-155.
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
發明與創新(2016年38期)2016-08-22 03:02:52
