干旱脅迫對(duì)13個(gè)杜鵑品種生理特性的影響
2017-05-11 18:02:09胡肖肖唐宇力賈梅金荷仙
江蘇農(nóng)業(yè)科學(xué) 2017年6期
胡肖肖++唐宇力++賈梅++金荷仙

摘要:以13個(gè)品種杜鵑(Rhododendron)2年生苗為材料,研究不同程度干旱脅迫對(duì)杜鵑葉片形態(tài)、相對(duì)含水量、水分飽和虧缺、細(xì)胞膜透性(相對(duì)電導(dǎo)率)、最大光化學(xué)效率(Fv/Fm)、潛在光化學(xué)效率(Fv/Fo)的影響,評(píng)價(jià)杜鵑品種的抗旱性。研究結(jié)果表明,葉片形態(tài)、水分飽和虧缺、相對(duì)電導(dǎo)率值隨干旱脅迫程度增加而升高;相對(duì)含水量隨脅迫程度增加而降低;最大光化學(xué)效率及潛在光化學(xué)效率在重度脅迫下顯著下降。根據(jù)模糊隸屬函數(shù)綜合評(píng)價(jià)13個(gè)杜鵑品種的抗旱性,其抗旱性排序?yàn)閲?guó)旗紅>玉蝴蝶>大青蓮>胭脂蜜>大鴛鴦錦>藍(lán)櫻>狀元紅>紅月>新天地>紫萼>琉球紅>粉珊瑚>紅珊瑚。各杜鵑品種均可以通過(guò)其形態(tài)、生理特性來(lái)適應(yīng)干旱脅迫環(huán)境。
關(guān)鍵詞:杜鵑;干旱脅迫;抗旱性;生理特性;隸屬函數(shù)
中圖分類號(hào): S685.210.1文獻(xiàn)標(biāo)志碼: A文章編號(hào):1002-1302(2017)06-0101-04
杜鵑花屬(Rhododendron)為杜鵑花科(Ericaceae)中最大的屬,全世界有1 000余種,中國(guó)約有562種,為世界杜鵑分布中心之一[1]。杜鵑花是中國(guó)十大名花之一,其種類繁多,花色艷麗,具有較高的觀賞價(jià)值、經(jīng)濟(jì)價(jià)值和藥用價(jià)值,在園林中應(yīng)用廣泛。根據(jù)杜鵑的產(chǎn)地來(lái)源、親緣關(guān)系、形態(tài)習(xí)性和觀賞特征,把栽培的杜鵑屬品種分成東鵑、毛鵑、西鵑、夏鵑4個(gè)類型[2],由于起源與地理環(huán)境的不同,各杜鵑品種對(duì)環(huán)境的適應(yīng)性上表現(xiàn)出較大差異[3],因此對(duì)于不同類型的杜鵑品種,在栽培管理上需要采取相應(yīng)的管理措施。
杜鵑在園林綠化及室內(nèi)觀賞栽培中,最易受到水分的影響[4-5],尤其對(duì)干旱脅迫比較敏感,缺水會(huì)導(dǎo)致其生長(zhǎng)受阻而死亡[6]。杜鵑園藝品種經(jīng)過(guò)引種馴化適應(yīng)新的環(huán)境,但是往往在實(shí)地應(yīng)用中存在諸如高溫、土壤供給水分不足等因素影響而導(dǎo)致杜鵑生長(zhǎng)不良的現(xiàn)象。目前,針對(duì)杜鵑園藝品種的耐旱性研究較少,主要集中在土壤水分脅迫對(duì)杜鵑生理生化指標(biāo)特性及抗旱性評(píng)價(jià)方面[4-5,7],對(duì)杜鵑在干旱脅迫下葉片形態(tài)以及熒光特性變化的研究甚少。本研究選取13個(gè)觀賞價(jià)值較高的杜鵑品種,通過(guò)設(shè)置不同干旱梯度的處理,研究不同品種杜鵑對(duì)土壤水分脅迫的生理、形態(tài)、熒光參數(shù)變化,以期篩選出耐旱的杜鵑品種,為西湖景區(qū)杜鵑花的水肥管理和栽培養(yǎng)護(hù),提供理論依據(jù)。
1材料與方法
1.1試驗(yàn)材料
供試杜鵑品種13個(gè)——大青蓮、紅珊瑚、粉珊瑚、大鴛鴦錦、胭脂蜜、紅月、藍(lán)櫻、新天地、琉球紅、狀元紅、國(guó)旗紅、玉蝴蝶和紫萼,購(gòu)于嘉興嘉善魏塘杜鵑盆景園,上述品種在西湖景區(qū)靈隱路段及靈隱勝境廣場(chǎng)應(yīng)用表現(xiàn)較好。2015年4月將長(zhǎng)勢(shì)一致的2年生苗木定植于高25 cm、直徑30 cm的普通塑料花盆內(nèi),盆土為普通園土,每盆盛土量一致,放置于杭州植物園科研中心溫室大棚(30°15′N,120°07′E)進(jìn)行試驗(yàn)。
1.2處理方法
于2015年10月7日給苗木澆水到飽和,隨后連續(xù)干旱處理。處理0 d,測(cè)得當(dāng)天土壤含水量為28.8%,并以此設(shè)置為對(duì)照組(CK);在干旱脅迫的15、35、55 d,測(cè)得其土壤含水量范圍依次為10%~20%、5%~9%、0~4%。按照Hsiao等對(duì)干旱脅迫程度的劃分方法[8-9],干旱15、35、55 d分別達(dá)到輕度干旱(LD)、中度干旱(MD)、重度干旱(SD)。在干旱處理的0、15、35、55 d分別觀察記錄植物葉片形態(tài),并測(cè)定葉片相對(duì)含水量、水分飽和虧缺以及細(xì)胞膜透性,并用Li-6400便攜式光合儀在溫室大棚內(nèi)活體測(cè)最大光化學(xué)效率(Fv/Fm)以及潛在光化學(xué)效率(Fv/Fo)。其中葉片形態(tài)評(píng)價(jià)參照文獻(xiàn)[10]進(jìn)行;采用烘干和飽和稱重法測(cè)定葉片含水量;采用電導(dǎo)法[11]測(cè)定葉片細(xì)胞膜透性。
1.3抗旱性綜合評(píng)價(jià)
根據(jù)干旱脅迫后測(cè)定的各項(xiàng)指標(biāo),采用模糊數(shù)學(xué)中的隸屬函數(shù)值法進(jìn)行抗旱性綜合評(píng)價(jià)。如果某一指標(biāo)與抗旱性呈正相關(guān),則采用隸屬函數(shù)計(jì)算,公式為:
X(μ)=(X-Xmin)/(Xmax-Xmin)。
如果某一指標(biāo)與抗旱性呈負(fù)相關(guān),則采用反隸屬函數(shù)計(jì)算,公式為:
X(μ)=1-(X-Xmin)/(Xmax-Xmin)。
式中:X(μ)為隸屬函數(shù)值,X為干旱脅迫下某一指標(biāo)的測(cè)定值,Xmin和Xmax為所有參試指標(biāo)中某一指標(biāo)的最大值和最小值。將參試材料所有指標(biāo)的隸屬函數(shù)值加起來(lái)求其平均值,用平均值大小綜合評(píng)價(jià)抗旱性強(qiáng)弱。
1.3數(shù)據(jù)分析
采用SPSS 16.0統(tǒng)計(jì)軟件進(jìn)行方差分析,同類數(shù)據(jù)顯著性差異運(yùn)用Duncans檢驗(yàn)法進(jìn)行多重比較。
2結(jié)果與分析
2.1干旱脅迫對(duì)葉片形態(tài)的影響
13個(gè)杜鵑花品種隨著干旱脅迫時(shí)間的延長(zhǎng),脅迫程度加重,土壤水分含量由飽和到輕度干旱(LD)、中度干旱(MD)、重度干旱(SD),葉片萎蔫指數(shù)也逐漸加重(表1)。不同時(shí)段處理下葉片萎蔫平均級(jí)別分別為0.00、0.87、2.31、3.41級(jí),顯然隨著干旱級(jí)別的增加,植物的萎蔫級(jí)別升高,葉片萎蔫程度加重。控水15 d(輕度干旱),紅珊瑚、粉珊瑚、紅月、新天地、狀元紅萎蔫級(jí)別達(dá)到1.33以上,其中新天地達(dá)到 1.67,顯著高于大青蓮、大鴛鴦錦、胭脂蜜、藍(lán)櫻、琉球紅、國(guó)旗紅、玉蝴蝶、紫萼;控水35 d(中度干旱),紅珊瑚、粉珊瑚、紅月、狀元紅這4個(gè)品種的葉片萎蔫指數(shù)達(dá)到3.00,國(guó)旗紅最低,僅有1.00;控水55 d(重度干旱),粉珊瑚與琉球紅的平均萎蔫級(jí)別增加至4.5級(jí),紅珊瑚葉片平均萎蔫級(jí)別增加至4級(jí),而國(guó)旗紅葉片平均萎蔫級(jí)別只有1.67級(jí),顯著低于其他各品種。
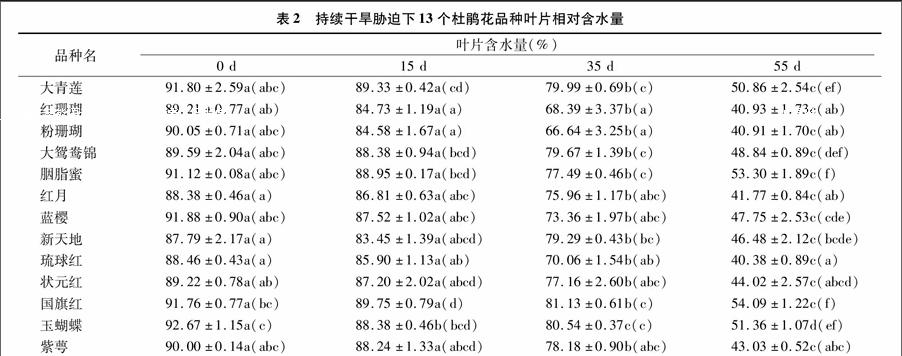
2.2干旱脅迫對(duì)葉片相對(duì)含水量的影響
葉片相對(duì)含水量能直觀地反映植物體內(nèi)水分生理的變化情況。植物在受到干旱脅迫時(shí),葉片相對(duì)含水量會(huì)隨著干旱脅迫程度的加深而下降,抗旱性越強(qiáng)降幅越小。由表2可見(jiàn),13個(gè)杜鵑品種的葉片相對(duì)含水量,不同時(shí)段干旱處理下存在
較大差異。處理0 d,葉片含水量在87.79%~92.67%,新天地含水量最低;控水15 d,玉蝴蝶下降幅度最大,其余12個(gè)品種下降幅度較小;控水35~55 d,葉片含水量大幅降低,其中重度干旱脅迫(55 d)下,葉片含水量只有40.38%~54.09%,琉球紅最低,只有40.38%,表現(xiàn)差異顯著(P<005)。
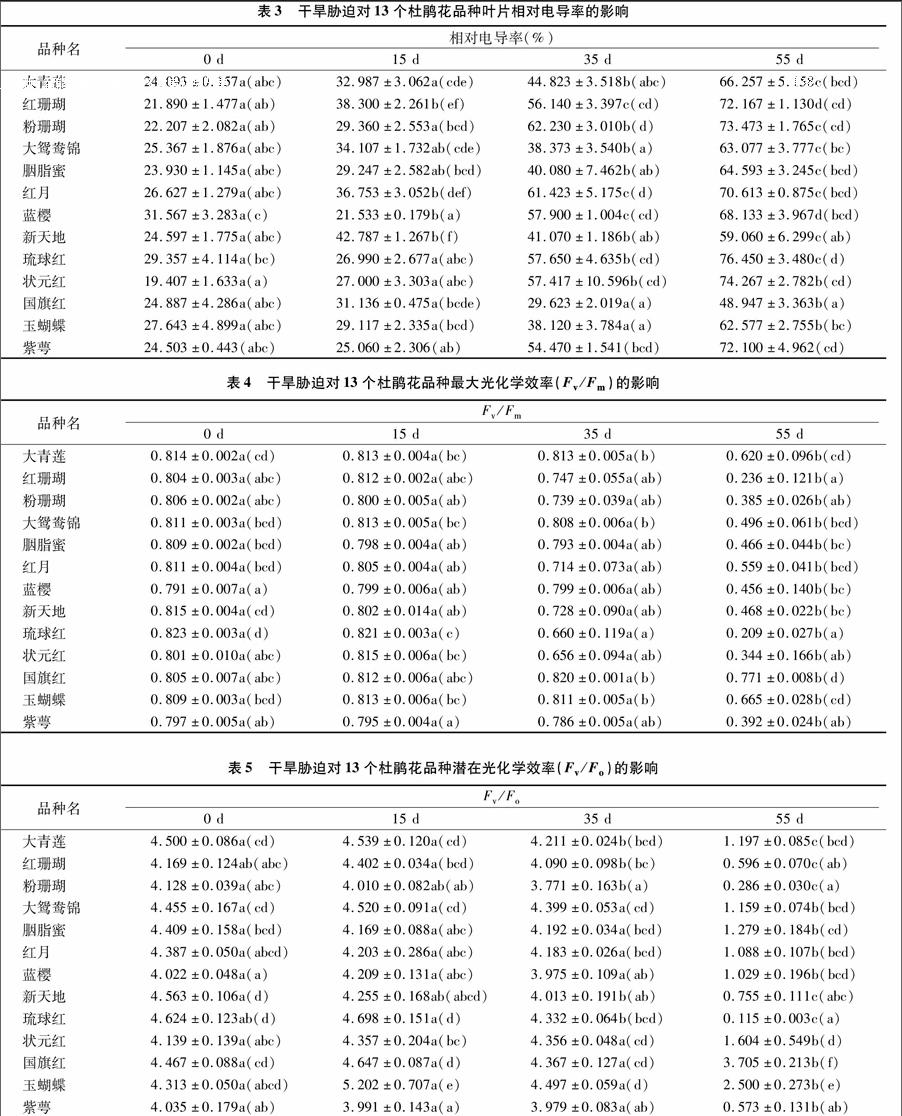
2.3干旱脅迫對(duì)葉片相對(duì)電導(dǎo)率的影響
電導(dǎo)率反映了在脅迫下細(xì)胞膜透性情況,外滲電導(dǎo)率低的種質(zhì)抵御干旱能力較強(qiáng),有利于維持細(xì)胞膜的穩(wěn)定性,保持細(xì)胞的正常生命代謝[12]。由表3可知,在輕度脅迫條件下,藍(lán)櫻和琉球紅的相對(duì)電導(dǎo)率較CK略有降低,其余杜鵑花品種葉片相對(duì)電導(dǎo)率都不同程度上升,且紅珊瑚、紅月、藍(lán)纓、新天地相對(duì)電導(dǎo)率與對(duì)照差異顯著,其余品種與對(duì)照差異不顯著;比較LD處理與MD處理下相對(duì)電導(dǎo)率值,除大鴛鴦錦、胭脂蜜、新天地、國(guó)旗紅、玉蝴蝶差異不顯著外,其他品種差異顯著(P<0.05),且國(guó)旗紅和玉蝴蝶2個(gè)品種在CK、LD和MD條件下,相對(duì)電導(dǎo)率值差異不顯著;比較MD處理與SD處理下相對(duì)電導(dǎo)率值,除紅月和狀元紅表現(xiàn)差異不顯著外,其余品種差異均顯著(P<0.05)。
2.4干旱脅迫對(duì)葉綠素?zé)晒鈪?shù)的影響
葉綠素可變熒光(Fv)反映光合作用中的光系統(tǒng)Ⅱ(PSⅡ)原初電子受體QA的還原情況,最大熒光產(chǎn)量(Fm)是PSⅡ反應(yīng)中心處于完全關(guān)閉時(shí)的量子產(chǎn)量,可反映通過(guò)PSⅡ的電子傳遞情況[13]。Fv/Fm是葉綠體PSⅡ最大光化學(xué)量子產(chǎn)量,它反映PSⅡ反應(yīng)中心內(nèi)的光能轉(zhuǎn)換效率。表4可見(jiàn),與CK相比,持續(xù)控水15~35 d,13個(gè)杜鵑品種的Fv/Fm值均無(wú)差異性;持續(xù)控水55 d,13個(gè)杜鵑品種的Fv/Fm下降明顯,但各品種下降幅度不一,國(guó)旗紅的Fv/Fm值最高(0.771),琉球紅最低,僅有0.209。
Fv/Fo為潛在光化學(xué)效率,可以反映光反應(yīng)中心PSⅡ的潛在活性。由表5可知,控水15 d,13個(gè)杜鵑品種的Fv/Fo與對(duì)照差異均不顯著;控水35 d,除大青蓮、紅珊瑚、粉珊瑚、
新天地、琉球紅外,其余杜鵑品種的Fv/Fo值與對(duì)照差異不顯著;控水55 d,大鴛鴦錦、胭脂蜜、紅月、藍(lán)櫻、狀元紅、國(guó)旗紅、玉蝴蝶、紫萼與CK組比較差異顯著。
2.513個(gè)杜鵑品種抗旱性綜合評(píng)價(jià)
由表6可知,各品種平均隸屬函數(shù)值在0.551~0.687之間,其中國(guó)旗紅、玉蝴蝶、大青蓮、胭脂蜜的隸屬函數(shù)值相對(duì)較大,分別為0.687、0.668、0.662、0.657,琉球紅、紅珊瑚、粉珊瑚的隸屬函數(shù)平均值較小,為0.577、0.551、0.554,剩余杜鵑品種隸屬函數(shù)值介于上述品種之間。說(shuō)明國(guó)旗紅、玉蝴蝶、大青蓮、胭脂蜜抗旱性最強(qiáng),琉球紅、紅珊瑚、粉珊瑚抗旱性最弱。
3結(jié)論與討論
逆境條件下植物外部形態(tài)發(fā)生明顯變化,葉片變化最為明顯,在干旱脅迫下會(huì)發(fā)生運(yùn)動(dòng),如葉片萎蔫、方位改變和葉角變化等[14]。眾多研究者采用葉片萎蔫等級(jí)來(lái)鑒定植株在干旱脅迫下的葉片受害程度,且級(jí)別越高,受害程度越大[10,15-16]。本研究中,各品種的萎蔫程度隨干旱脅迫程度增加而增大,這與葉片含水量降低有直接關(guān)系。葉片相對(duì)含水量可以準(zhǔn)確反映植物體內(nèi)的水分狀況,在持續(xù)控水條件下,土壤水分含量降低,杜鵑進(jìn)行新陳代謝和蒸騰作用消耗大量的水分,杜鵑的葉片相對(duì)含水量出現(xiàn)一定幅度下降。含水量降低,改變了原生質(zhì)膜透性,原生質(zhì)膜是細(xì)胞內(nèi)外的一層屏障,可以有效地防御水分脅迫等逆境引起的傷害,從而維持細(xì)胞結(jié)構(gòu)的穩(wěn)定性。植物處于干旱脅迫下時(shí),細(xì)胞膜透性增大,穩(wěn)定性降低,大量離子外滲,葉片細(xì)胞膜外滲液的電導(dǎo)率增大,細(xì)胞膜的選擇透性改變或喪失,從而導(dǎo)致細(xì)胞膜損傷[17-18]。本試驗(yàn)中,各品種葉片細(xì)胞膜外滲液的電導(dǎo)率隨干旱脅迫加劇而增加,且抗旱性較強(qiáng)的品種的相對(duì)電導(dǎo)率變化較小。
葉綠素?zé)晒鈪?shù)能夠?qū)χ参矬w內(nèi)的光合機(jī)構(gòu)運(yùn)轉(zhuǎn)情況進(jìn)行無(wú)損傷診斷,可以反映逆境脅迫下PSⅡ反應(yīng)中心的受損程度[19]。Li等將葉綠素?zé)晒鈪?shù)作為干旱脅迫品種比較的主要依據(jù),認(rèn)為水分脅迫會(huì)對(duì)植株葉片的Fv/Fm產(chǎn)生影響[20]。Fv/Fm值一般恒定為0.80~0.85,當(dāng)植物受到環(huán)境脅迫時(shí),F(xiàn)v/Fm值會(huì)發(fā)生顯著變化,因此,它是研究植物光合作用和生理生化的一個(gè)使用最廣的熒光參數(shù)[21]。本研究發(fā)現(xiàn)杜鵑在15~35 d控水干旱脅迫下,各品種Fv/Fo和Fv/Fm下降幅度較小,與CK差異不顯著,說(shuō)明中度干旱脅迫對(duì)其還未產(chǎn)生傷害。這一測(cè)定結(jié)果還可能與10月份的杭州天氣有關(guān),其溫度在12~25 ℃,濕度在60%~90%,陰雨天氣占2/3左右,植株在高濕度環(huán)境中,蒸騰作用減弱,失水較少。在重度干旱脅迫下,13個(gè)杜鵑品種的Fv/Fm值才顯著下降,且各品種下降幅度不一,國(guó)旗紅的Fv/Fm值為0.771,表明受傷害程度較小,玉蝴蝶、大青蓮的值也在0.60以上,表現(xiàn)出較強(qiáng)的抗旱能力。
綜合各指標(biāo)隸屬函數(shù)值比較13個(gè)品種抗旱性強(qiáng)弱,由強(qiáng)到弱依次為國(guó)旗紅>玉蝴蝶>大青蓮>胭脂蜜>大鴛鴦錦>藍(lán)櫻>狀元紅>紅月>新天地>紫萼>琉球紅>粉珊瑚>紅珊瑚。在園林植物應(yīng)用中,結(jié)合植物觀賞特性,擇優(yōu)選擇較耐旱的杜鵑品種應(yīng)用于實(shí)際中,有利于后期對(duì)植物的水肥管理和栽培養(yǎng)護(hù)。
參考文獻(xiàn):
[1]余樹(shù)勛. 杜鵑花[M]. 北京:金盾出版社,1992.
[2]黃茂如. 杜鵑花[M]. 上海:上海科學(xué)技術(shù)出版社,1998.
[3]方瑞征,閔天祿. 杜鵑屬植物區(qū)系的研究[J]. 云南植物研究,1995,17(4):359-379.
[4]柯世省,楊敏文. 水分脅迫對(duì)云錦杜鵑抗氧化系統(tǒng)和脂類過(guò)氧化的影響[J]. 園藝學(xué)報(bào),2007,34(5):1217-1222.
[5]張曉勤,吳克利,薛大偉. 濕害對(duì)不同大麥基因型抗氧化酶系統(tǒng)的影響[J]. 浙江大學(xué)學(xué)報(bào)(農(nóng)業(yè)與生命科學(xué)版),2009,35(3):315-320.
[6]李波,吳月燕,崔鵬. 水分脅迫對(duì)兩種基因型杜鵑生理生化特性的影響[J]. 浙江農(nóng)業(yè)學(xué)報(bào),2011,23(5):988-994.
[7]周廣,孫寶騰,張樂(lè)華,等. 井岡山杜鵑葉片抗氧化系統(tǒng)對(duì)高溫脅迫的響應(yīng)[J]. 西北植物學(xué)報(bào),2010,30(6):1149-1156.
[8]Hsiao T C. Plant responses to water stress[J]. Ann Rev Plant Physiol,1973,24:519-570.
[9]柯世省,金則新. 干旱脅迫與復(fù)水對(duì)夏臘梅幼苗光合生理特性的影響[J]. 植物營(yíng)養(yǎng)與肥料學(xué)報(bào),2007,13(6):1166-1172.
[10]田治國(guó),王飛,張文娥,等. 萬(wàn)壽菊屬不同品種初花期抗旱特性分析[J]. 西北植物學(xué)報(bào),2011,31(7):1390-1399.
[11]李合生,孫群,趙世杰,等. 植物生理生化實(shí)驗(yàn)原理和技術(shù)[M]. 北京:高等教育出版社,2000:184-185.
[12]薛瑞,周廣奇,胡新文,等. 柱花草種質(zhì)抗旱性綜合評(píng)價(jià)[J]. 中國(guó)農(nóng)學(xué)通報(bào),2009,25(11):224-233.
[13]朱新廣,王強(qiáng),張其德,等. 冬小麥光合功能對(duì)鹽脅迫的響應(yīng)[J]. 植物營(yíng)養(yǎng)與肥料學(xué)報(bào),2002,8(2):177-180.
[14]Navas M L,Garnier E. Plasticity of whole plant and leaf trait in Rubia peregrine in response to light,nutrient and water availability[J]. Acta Oecologica-International Journal of Ecology,2002,23(6):375-383.
[15]周招娣,張日清,馬錦林,等. 6個(gè)油茶物種苗期抗旱性的初步研究[J]. 經(jīng)濟(jì)林研究,2014,32(2):53-57.
[16]王曉娟,周蘭英. 干旱脅迫對(duì)紅花檵木生長(zhǎng)及生理特性的影響[J]. 四川農(nóng)業(yè)大學(xué)學(xué)報(bào),2015,33(1):22-32.
[17]Bohnert H J,Jebsen R G. Strategies for engineering water-stress tolerance in plants[J]. Trends in Biotechnology,1996,14(3):89-97.
[18]Munns R. Comparative physiology of salt and water stress[J]. Plant Cell and Environment,2002,25(2):239-250.
[19]Misra A N,Srivastava A,Strasserr J,et al. Utilization of fast chlorophyll a fluorescence technique in assessing the saltion sensitivity of mung bean and Brassica seedlings[J]. Journal of Plant Physiology,2001,158(9):1173-1181.
[20]Li L,van Staden J. Effcets of plant growth regulators on the antioxidant system in callus of two maize cultivars subjected to water stress[J]. Plant Growth Regulation,1998,24(1):55-56.
[21]張守仁. 葉綠素?zé)晒鈩?dòng)力學(xué)參數(shù)的意義及討論[J]. 植物學(xué)通報(bào),1999,16(4):444-448.杜玉玲,潘晨慧,李丹陽(yáng),等. 利用隸屬函數(shù)值法對(duì)7種一、二年生花卉耐鹽性的綜合評(píng)價(jià)[J]. 江蘇農(nóng)業(yè)科學(xué),2017,45(6):105-111.
doi:10.15889/j.issn.1002-1302.2017.06.026
