生物絮團對鯉魚養殖池塘浮游植物群落結構的影響
2017-06-05 13:39:33楊希李濤白海鋒高志袁永鋒賈秋紅
河北漁業 2017年5期
關鍵詞:群落結構
楊希+李濤+白海鋒+高志+袁永鋒+賈秋紅+高宏偉

摘要:為了解生物絮團技術在凈化池塘水質的同時對池塘初級生產力的影響,于2015年9月-10月對陜西團頭魴良種場鯉魚池塘的浮游植物群落結構進行了調查研究。結果顯示:生物絮團池塘共出現浮游植物6門28種,其中綠藻門有17種,占總種類數的65.4%,硅藻門有5種,占總種類數的19.2%,浮游植物群落結構類型屬綠藻-硅藻型。浮游植物生物密度變化范圍為1 372.82×104~3 093.71×104ind./L,平均值為2 217.07×104 ind./L;生物量變化范圍為3.53~12.87 mg/L,平均值為7.88 mg/L。浮游植物生物密度和生物量均在9月15日出現峰值,峰值期以后浮游植物密度和生物量在時間和空間上差異性顯著(P<0.05)。研究表明,生物絮團技術對池塘浮游植物群落結構影響較大,隨著絮團的逐漸形成浮游植物群落結構出現明顯變化,池塘藻相演替速度加快。
關鍵詞:生物絮團;鯉魚;浮游植物;群落結構
生物絮團技術是一種新型養殖技術,通過向養殖水體中補充有機碳源,調整碳氮比,使水體中的異養微生物通過同化作用吸收水中氨氮、亞硝酸氮等物質,從而達到凈化水質的效果。而微生物的大量繁殖過程絮凝了水中的絲狀藻類、原生動物、有機碎屑等物質形成絮狀物,這種生物絮團又可作為餌料蛋白,使蛋白得到二次利用,從而節省了成本,提高了養殖效益[1]。生物絮團因具有調節水質、減少病害發生、提高飼料蛋白利用率、降低養殖成本以及提高養殖動物消化和免疫性能等優點被廣泛應用于生產實踐[2]。目前,對于生物絮團技術在水產養殖上的應用研究較多[3-5],但是對其在養殖過程中的應用對浮游植物的影響研究未見報道。浮游植物是水生態系統中重要的初級生產者,是食物鏈中最低營養級,在決定水域生產性能上具有重要意義。浮游植物的種類組成、生物量、多樣性等特征與水環境之間關系密切,浮游植物對水質變化比較敏感,常常被用作水環境質量一項重要評價指標,水質的變化直接影響浮游植物群落結構的演替方向[6]。本研究通過對生物絮團池塘浮游植物群落的研究,科學地評價其生態環境狀況,以期為提高生物絮團在池塘養殖中的生態效應,促進池塘生態養殖技術推廣提供理論數據。
1材料與方法
1.1試驗池塘
試驗池塘選擇在西安草灘渭河南岸養殖區。池塘長方形,東西走向,日照充足,池壁用混凝土護坡,池底存有10 cm淤泥。池塘面積0.4 hm2,水深1.6 m。池塘主養鯉魚,規格為500~1 000 g/尾,養殖密度為15 000~22 500尾/hm2,池塘中混養少量鰱魚和鯽魚。池塘水源為深井水,池塘配有微孔增氧設備。
1.2試驗方法
1.2.1試驗設計
生物絮團采用鯉魚池塘原池培養,根據鯉魚每天的投飼量,全池潑灑飼料級蔗糖,維持池塘水體中碳氮比(C/N)為20∶1,定期(7~10 d)潑灑枯草芽孢桿菌。試驗期間間斷性開啟微孔增氧設備,維持水體溶解氧不低于5.0 mg/L。試驗設置對照組,對照組和實驗組各設置兩個平行池塘。試驗期間不換水,僅根據池塘水位適當添加新水,以彌補蒸發和滲漏引起的水分損失。在日常管理中,試驗塘和對照塘盡量做到一致,避免因池塘管理造成的誤差。
1.2.2樣品采集與鑒定
浮游植物樣品采集方法參照《湖泊生態調查觀測與分析》[7]。依據池塘情況,每5 d采樣一次,同時監測池塘水質,采樣時間選擇在上午8:30-9:30,采樣地點設置在池中央和池邊。水樣用5 000 mL有機玻璃采水器在水面下50 cm處采水樣1 000 mL,現場用魯哥氏液(加入量1%~1.5%)進行固定,固定好的樣本隨后帶回實驗室靜置24 h,用虹吸管濃縮至50 mL。依據相關參考文獻[8-10],在生物顯微鏡(放大倍數×400~×1 000)下鑒定種類并計數。
1.3數據處理與分析
浮游植物生物密度計算公式:
N=(V1 ×n)/(V×V2 )[11]
式中: N為1 000 mL水樣中浮游植物個體數(ind./L);V1為水樣沉淀濃縮后的體積(L);
n為計數所得的藻類個體數(ind.);V為采集到水樣的體積(1 000 mL);V2為計數體積(L)。
浮游植物生物量依據藻類種類濕重換算統計。
試驗數據采用SPSS 19.0軟件包進行數據統計分析。生物絮團池與對照池,以及生物絮團池塘不同時間的浮游植物平均密度、平均生物量的比較均采用單因素方差分析進行。
2結果與分析
2.1水質指標
試驗期間,生物絮團池塘水溫變化范圍為23.6~26.7 ℃,呈現隨氣溫逐漸下降的趨勢;透明度變化范圍為22~35 cm,隨時間的推移呈下降趨勢;pH變化幅度為6.8~7.7,溶解氧為662~9.33 mg/L,電導率為803~875 μs/cm,三者保持較穩定狀態;氨氮含量為0.406~2.162 mg/L,亞硝酸氮為0.066~0.242 mg/L,兩者表現出先升高后下降的波動趨勢。從整體來看,在時間尺度上,除氨氮和亞硝酸氮外,其余水質指標無顯著性差異(P>0.05)。對比對照池塘,生物絮團池塘透明度、氨氮、亞硝酸氮三項指標差異顯著(P<0.05)。
2.2浮游植物群落結構
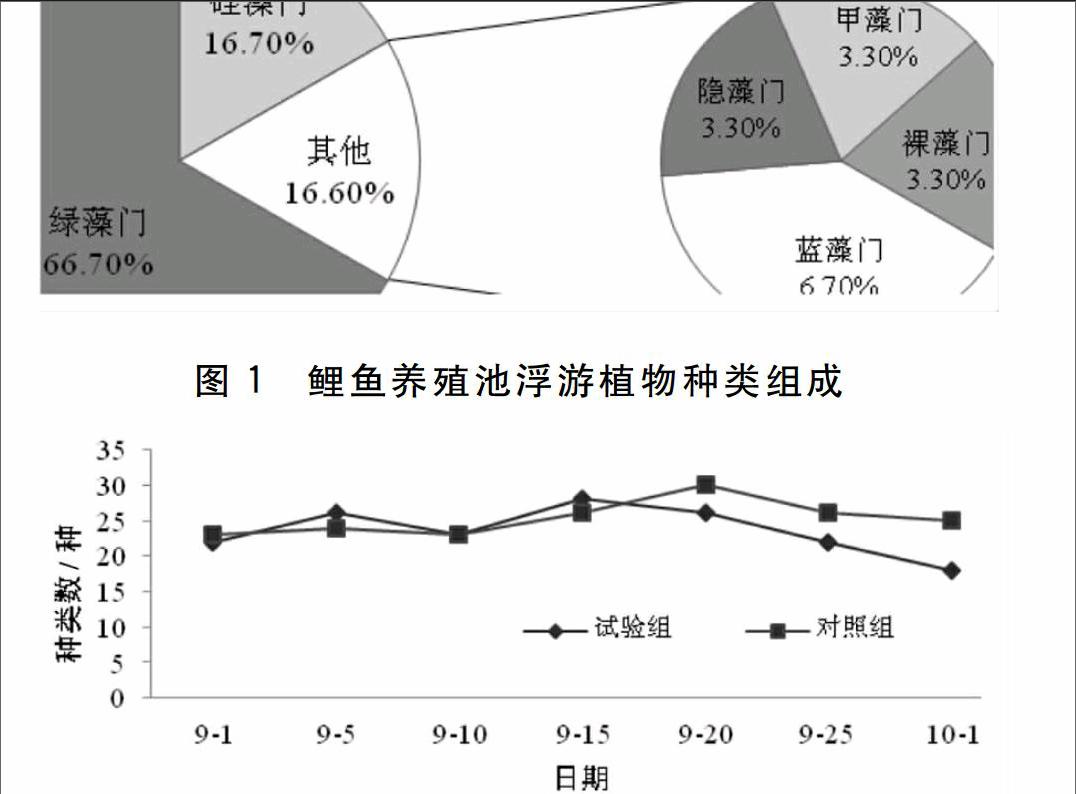
生物絮團池塘和對照池塘在試驗期間共鑒定出浮游植物6門30種。其中綠藻門20種,占總種類數的66.7%;硅藻門有5種,占16.7%;其他門共占16.6%,分別為藍藻門2種,隱藻門1種,甲藻門1種,裸藻門1種(見圖1)。生物絮團池塘浮游植物以綠藻門和硅藻門為主,其次是藍藻門,藻相呈綠藻-硅藻型。對照池塘以綠藻門和藍藻門為主,其次是硅藻門,藻相呈現綠藻-藍藻型。兩組池塘在種類數上差異不顯著(P>005),變化范圍為21~30種,種類總體呈逐漸上升的趨勢。
從時間尺度上看,生物絮團池塘浮游植種類變化呈現波浪型,在試驗第15 d(9月15日)出現高峰值(28種),其中綠藻門17種,占總種類數的654%,硅藻門5種,占19.2%,而在試驗結束時(10月1日)出現低谷值(21種)。在時間尺度上,生物絮團池塘浮游植物種類數存在顯著性差異(P<0.05),對照池差異不顯著(P>0.05)(圖2)。
試驗期間生物絮團池塘和對照池塘浮游植物密度均呈先升后降的趨勢,生物絮團池塘7月15日密度達到最大值,顯著高于其他時間(P<005),而對照池塘密度高峰期出現9月20日(見圖3)。生物絮團池塘浮游植物生物密度平均值為2 217.07×104 ind./L,在時間上的變化范圍為1 372.82×104~3 093.71×104 ind./L;對照池塘浮游植物密度平均值為2 610.55×104 ind./L,變化范圍在1 965.23×104~4 156.88×104 ind./L。兩者密度均值差異不顯著(P>0.05),但是從9月15日以后,兩者浮游植物密度在時間尺度上出現顯著性差異(P<0.05)。
從圖4可見,試驗期間試驗池塘和對照池塘浮游植物生物量的變化趨勢與密度基本一致。試驗池塘浮游植物生物量范圍在3.53~12.87 mg/L之間,平均值為7.88 mg/L。對照池塘的生物量在9月20日出現最高值,為18.85 mg/L,此后一直降低,試驗結束時生物量顯著降低,出現最低值,為4.62 mg/L(P<0.05)。從時間上來看,試驗池塘和對照池塘生物量在9月15日之前無顯著性差異(P>0.05),9月20日之后出現顯著性差異(P<0.05)。
3討論
浮游植物是水生態系統的初級生產者,能通過食物鏈將營養物質傳遞到更高營養層,對生態系統的物質循環和能量流動起到積極的推動作用[12]。浮游植物的群落結構以及光合作用受到養殖水環境的影響巨大,這些環境因子包括水溫、透明度、氮、磷以及養殖動物等。
氮、磷是生命組織中的必需元素,對浮游植物來說,適量的氮、磷能促進其生長。有研究顯示,通常情況下水中的浮游植物在吸收氮元素時,氨氮優先于其他氮形式,并且氨氮的含量往往直接影響浮游植物的生長[13]。在一定范圍內浮游植物生物密度隨著N/P的升高而增加,一旦遠離正常比值(16)時,浮游植物的生長繁殖將受到抑制[14]。從本試驗可以看出,生物絮團池塘的浮游植物的種類數、生物密度和生物量均在9月15日之后開始降低或減少,而對照組這種下降點則延遲了一段時間。這種差異分析認為,生物絮團一般從開始形成到成熟需要10~15 d的時間[15],在這段時間里水體中的氨氮、亞硝酸氮含量變化不大,當生物絮團系統處于穩態后,其對池塘水中氨氮和亞硝酸氮的去除率逐漸增大,導致池塘中可供浮游植物生長所需的營養元素減少,從而使浮游植物在試驗中后期出現漸減的現象。
生物絮團技術的應用需要充足的溶解氧,因此在實施過程中需要不斷地充氣攪水,使生物絮團懸浮于水中,以方便魚類攝食,另一方面防止絮團下沉。調查發現,在試驗池由于生物絮團的懸浮導致水體透明度相比對照池降低,從而影響了浮游植物的光合作用。有研究表示[16],透明度是影響水體中浮游植物種類和數量變化的重要因素之一。此外透明度能夠直觀反映水體中的懸浮物數量,其與浮游植物生物量成負相關[17]。本試驗發現,由于透明度和水溫的變化影響,試驗池塘浮游植物群落結構發生了變化,藻相從喜溫、適宜較高透明度的綠藻-藍藻型演變到適應新環境的綠藻-硅藻型。
以上研究表明,生物絮團技術在鯉魚養殖過程中對水中氨氮和亞硝酸氮的去除以及水質凈化具有顯著效果。但是其對養殖水體浮游植物群落結構也存在顯著影響,尤其對池塘浮游植物的光合作用影響較大,眾所周知,池塘養殖水體中的溶解氧絕大部分來源于水體浮游植物光合作用。因此,在水產養殖中積極應用推廣生物絮團技術時,合理地控制生物絮團生物量和有效的管理模式是生物絮團技術生態高效養殖模式的關鍵,也是今后研究的主要方向。
參考文獻:
[1]
羅文,王廣軍,龔望寶,等.生物絮團技術對彭澤鯽生長及養殖水質的影響[J].南方農業學報,2014,45(2):318-322
[2] 白海鋒,袁永鋒,賈秋紅,等.生物絮團對養魚池塘水質凈化效果試驗[J].水產養殖,2015,(9):32-33
[3] 鄧應能,趙培,孫運忠,等.生物絮團在凡納濱對蝦封閉養殖試驗中的形成條件及作用效果[J].漁業科學進展,2012,33(2):69-75
[4] 李朝兵,王廣軍,余德光,等.生物絮團對鳙生長、肌肉氨基酸成分及營養評價的影響[J].江蘇農業科學,2012,40(11):242-245
[5] 夏耘,邱立疆,郁二蒙,等.生物絮團培養過程中養殖水體水質因子及原核與真核微生物的動態變化[J].中國水產科學,2014,21(1):75-83
[6] 劉冬艷,孫軍,張利永.膠州灣浮游植物水華期群落結構特征[J].應用生態學報,2003,14(11):1963-1966
[7] 黃祥飛.湖泊生態調查觀測與分析[M].北京:中國標準出版社,1999
[8] 周鳳霞,陳劍虹.淡水微型生物與底棲動物圖譜[M].4版.北京:化學工業出版社,2011
[9] 韓茂森,束藴芳.中國淡水生物圖譜[M].北京:海洋出版社,1995
[10] 梁象秋,方紀祖,楊和荃.水生生物學(形態和分類)[M].北京:中國農業出版社,1996
[11] 趙文.水生生物學[M].北京:中國農業出版社,2005
[12] 賈秋紅,陳曉霞,王博涵,等.異育銀鯽苗種培育池塘浮游植物群落結構動態變化[J].河北漁業,2016(1):9-11
[13] 劉峰,高云芳,王立欣,等.水域沉積物氮磷賦存形態和分布的研究進展[J].水生態學雜志,2011,32(4):137-144
[14] Schanz F, Juon H. Two different methods of evaluating nutrient limitation of periphyton bioassays,using water from the River Rhine and eight of its tributaries[J]. Hydrobiologia,1983,102:187-195
[15] 李志斐,王廣軍,余德光,等.生物絮團對養殖水體水質和微生物群落功能的影響[J].上海海洋大學報,2015,24(4):503-512
[16] 蔣嫣紅,程婧蕾,王麗卿.公園水體浮游植物與環境因子的關系[J].生態學雜志,2012,31(3):606-613
[17] 何志輝,李永涵.無錫市河埒口高產魚池水質的研究Ⅱ.浮游生物[J].水產學報,1983,7(4):287-298
猜你喜歡
熱帶農業科學(2016年11期)2017-01-21 14:59:21
熱帶農業科學(2016年11期)2017-01-21 14:57:08
河北漁業(2015年12期)2015-12-21 14:37:31
河北漁業(2015年11期)2015-11-18 16:39:54
河北漁業(2015年11期)2015-11-18 16:37:49
河北漁業(2015年10期)2015-10-15 11:14:57
湖北農業科學(2015年18期)2015-10-13 21:34:01
河北漁業(2015年7期)2015-07-21 11:06:52
河北漁業(2015年6期)2015-06-24 11:04:37
安徽農學通報(2015年5期)2015-03-23 23:51:20
