肝X受體激動劑GW3965對高糖所致心肌細胞凋亡的影響及作用機制*
2017-07-18 11:48:06成永霞張大偉孫立新馮玉寬郭素芬劉貴波
陜西醫學雜志 2017年7期
關鍵詞:檢測
成永霞,張大偉,孫立新,馮玉寬,郭素芬,劉貴波Δ
1.牡丹江醫學院病理學教研室(牡丹江157011),2.牡丹江醫學院解剖教研室(牡丹江157011)
·基礎研究·
肝X受體激動劑GW3965對高糖所致心肌細胞凋亡的影響及作用機制*
成永霞1,張大偉2,孫立新1,馮玉寬1,郭素芬1,劉貴波2Δ
1.牡丹江醫學院病理學教研室(牡丹江157011),2.牡丹江醫學院解剖教研室(牡丹江157011)
目的:觀察GW3965對高糖誘導的心肌細胞凋亡的影響,并初步探討其作用機制。方法:取對數期H9c2心肌細胞,隨機分為對照組(Control組)、高糖誘導組(高糖組)和高糖誘導+激動劑處理組(GW3965組)。GW3965組細胞加入10 μmol/L GW3965預處理24 h,之后高糖組和GW3965組細胞加入33 mmol/L葡萄糖繼續誘導培養24 h。采用MTT法檢測細胞存活率,流式細胞術檢測細胞凋亡率,JC-1熒光染料法檢測線粒體膜電位,Western blot檢測LXRα、ERK和p38 MAPK信號通路相關蛋白的變化。結果:GW3965可以提高H9c2抑制細胞凋亡,提高細胞存活率,改善細胞線粒體膜電位變化,并抑制ERK和p38 MAPK通路活性。結論:GW3965通過介導LXRα和MAPK通路減輕高糖誘導的細胞凋亡和線粒體損傷,對心肌細胞具有保護作用。
近年來,伴隨我國經濟迅速發展,人民當代生活水平的提高以及飲食結構的改變,糖尿病呈現逐年攀升趨勢。糖尿病心肌病(Diabetic cardiomyopathy, DCM)是糖尿病患者的主要心臟并發癥之一,發病率高,危害性大,對患者健康造成嚴重危害,與心血管疾病的發生緊密相關,亦是導致糖尿病患者心力衰竭、休克甚至猝死的關鍵原因[1]。資料顯示,約70 %~80 %的臨床患者死于DCM[2]。雖然目前醫學界針對預防和治療DCM已經展開了大量研究,但收效甚微,并未顯著減少臨床患者的進展和死亡。因此深入探索DCM潛在的發病機制,尋找新型治療靶點對提高臨床患者的生存質量具有重大意義。
DCM的發病機制十分復雜,涉及眾多因素。伴隨研究者不斷的深入發掘和探索,目前認為血糖及心肌能量代謝紊亂、鈣水平異常、微血管病變、炎癥反應、氧化應激、線粒體功能障礙和心肌細胞凋亡等多種因素參與DCM的發病與進展,其中心肌細胞凋亡是DCM重要病理機制之一,有效抑制心肌細胞凋亡將有利于DCM的防治[3]。
新近研究發現[4],肝X受體(Liver X receptors, LXRs)是人體內藥物治療潛在分子靶標,其激動劑可以被應用于多種疾病的治療。有文獻報道[3],LXRs激動劑T0901317能夠減輕高糖誘導的氧化應激、線粒體損傷和細胞凋亡。GW3965作為一種人工合成的LXRs的另一種強效激動劑,亦被證明具有心肌保護作用,能夠改變脂質代謝,誘導甘油三酸酯的合成,減輕缺血再灌注對心臟造成的損傷[5]。此外,GW3965還可以減輕胰島素抵抗,抑制氧化應激、炎癥反應、JNK和p38 MAPK信號通路的活性[6]。但GW3965對高糖誘導的心肌細胞凋亡及線粒體損傷的影響目前尚未見文獻報道。
因此,本研究采用高糖誘導H9c2建立體外細胞模型,同時加入LXRs激動劑GW3965進行干預,結合MTT、流式細胞術、JC-1法及Western blot等實驗技術分析細胞增殖、凋亡、線粒體膜電位以及ERK、p38 MAPK信號通路活性變化,探討GW3965對高糖誘導的心肌細胞損傷的保護作用,初步明確具體分子機制。
材料與方法
1 材 料 大鼠心肌細胞H9c2(本實驗室保存);胎牛血清(Hyclone公司);DMEM培養基(Gibco)公司;胰酶、RIPA裂解液、PMSF、BCA蛋白濃度測定試劑盒、一抗二抗去除液、IgG-HRP(碧云天生物公司);GW3965(Selleckchem公司);MTT(萬類生物公司);DMSO(Sigma公司);細胞凋亡檢測試劑盒、線粒體膜電位檢測試劑盒(凱基生物公司);TEMED(AMRESCO公司);預染蛋白分子量標準(Fermentas公司);PVDF膜(Millipore公司);ECL發光液(七海生物公司);LXRα抗體(Abcam公司);p38 MAPK抗體(BOSTER公司);p-p38 MAPK、ERK、p-ERK抗體(Bioss公司);內參抗體β-actin(Santa公司);BSA(Biosharp公司)。
2 研究方法
2.1 細胞處理與分組:H9c2心肌細胞接種于含10 % FBS的DMEM培養基中,37 ℃,5 % CO2環境下常規培養。將生長至對數期的細胞隨機分為三組:Control組、高糖組和GW3965組,胰酶消化細胞,顯微鏡下觀察到細胞變圓后終止反應,收集細胞,低溫條件下88 g離心3 min,去上清,加入1 ml培養基重新懸浮細胞。計數后按照實驗分組重新將細胞接種于6孔板中,每孔4×105個,37 ℃培養箱內培養24 h后GW3965組細胞加入10 μmol/L激動劑處理24 h,之后高糖組和GW3965組細胞均加入33 mmol/L葡萄糖繼續培養24 h。
2.2 MTT檢測細胞活力:取對數期H9c2細胞,胰酶消化收集細胞懸液,計數并調整細胞密度。取5×103個細胞接種于培養板內,按照實驗分組每組設置5個復孔,置于37 ℃,5 % CO2的培養箱內培養24 h,隨后按照2.1的方法處理各組細胞。到達指定時間后向各孔內加入5 mg/ml的MTT,37 ℃孵育4 h,去除上清液,加入200 μl DMSO溶解細胞形成的紫色結晶,酶標儀測定各組細胞在490 nm波長處的吸光值。
2.3 流式細胞術分析細胞凋亡:待H9c2細胞生長密度達90 %左右,加入胰酶消化細胞,待細胞變圓后終止消化,離心收集細胞沉淀,加入培養液重新懸浮細胞,計數并調整細胞密度,接種于6孔板內,置于37 ℃,5 % CO2培養箱內培養24 h,隨后按照2.1的方法處理各組細胞。到達指定時間后,309 g離心5 min收集細胞沉淀,PBS清洗2次,最后一次清洗殘留約50 μl緩沖液,加入500 μl Binding Buffer吹打懸浮細胞,加入5 ml Annexin V-FITC,輕輕混勻后加入5 μl Propidium Iodide,室溫避光孵育15 min,流式檢測。
2.4 JC-1法檢測線粒體膜電位:待H9c2細胞生長密度達90 %左右,胰酶消化收集細胞,離心收集細胞沉淀,加入培養液重新懸浮細胞,計數后接種于6孔板內,置于37 ℃,5 % CO2培養箱內培養24 h,隨后按照2.1的方法處理各組細胞。到達指定時間后,550 g離心5 min收集細胞沉淀,PBS清洗2次,加入500 μl JC-1工作液,輕輕吹打懸浮細胞,37 ℃條件下孵育20 min,550 g離心5 min,1×Incubation Buffer清洗細胞2次,去除上清,加入500μl 1×Incubation Buffer重新懸浮細胞,上流式分析線粒體膜電位變化。右上象限(UR)代表正常細胞,右下象限(LR)代表凋亡細胞。
2.5 Western blot檢測蛋白表達:收集各組細胞,加入含有1 % PMSF的RIPA裂解液,冰上靜置5 min充分裂解細胞,12000 rpm、4 ℃離心10 min,收集上清液,得到樣本蛋白;將BSA蛋白標準液進行梯度稀釋,繪制標準曲線,測定蛋白濃度。組裝電泳裝置,分別制備濃縮膠(5 %)和分離膠(10 %),灌膠、梳子封口、30 min后拔出梳子;緩沖液稀釋各樣本蛋白,取40 μg進行上樣,行SDS-PAGE,80 V恒壓電泳2.5 h。PVDF膜轉印、脫脂奶粉封閉,向雜交袋中加入對應一抗,4 ℃過夜孵育,TTBS清洗后加入IgG-HRP二抗,37 ℃孵育45 min。ECL顯色、暗室曝光,掃描膠片,Gel-Pro-Analyzer軟件分析目標條帶光密度值。
結 果
1 細胞凋亡率檢測 高糖誘導后,心肌細胞H9c2凋亡率為(20.02±1.07)%,同Control組(7.76±1.02)%比較顯著升高,差異具有統計學意義(P<0.01);采用激動劑GW3965干預后,細胞凋亡率為(13.04±1.19)%,與高糖組比較明顯降低,差異具有統計學意義(P<0.01)。說明激動劑GW3965可以抑制高糖誘導的H9c2細胞凋亡。
2 細胞存活率檢測 同對照組比較,高糖組OD值明顯降低,差異具有統計學意義(P<0.01),說明高糖抑制細胞增殖。與高糖組比較,GW3965組OD值顯著升高(P<0.01),細胞生存能力增強。由此可見,GW3965可以提高H9c2細胞的存活率(圖1)。
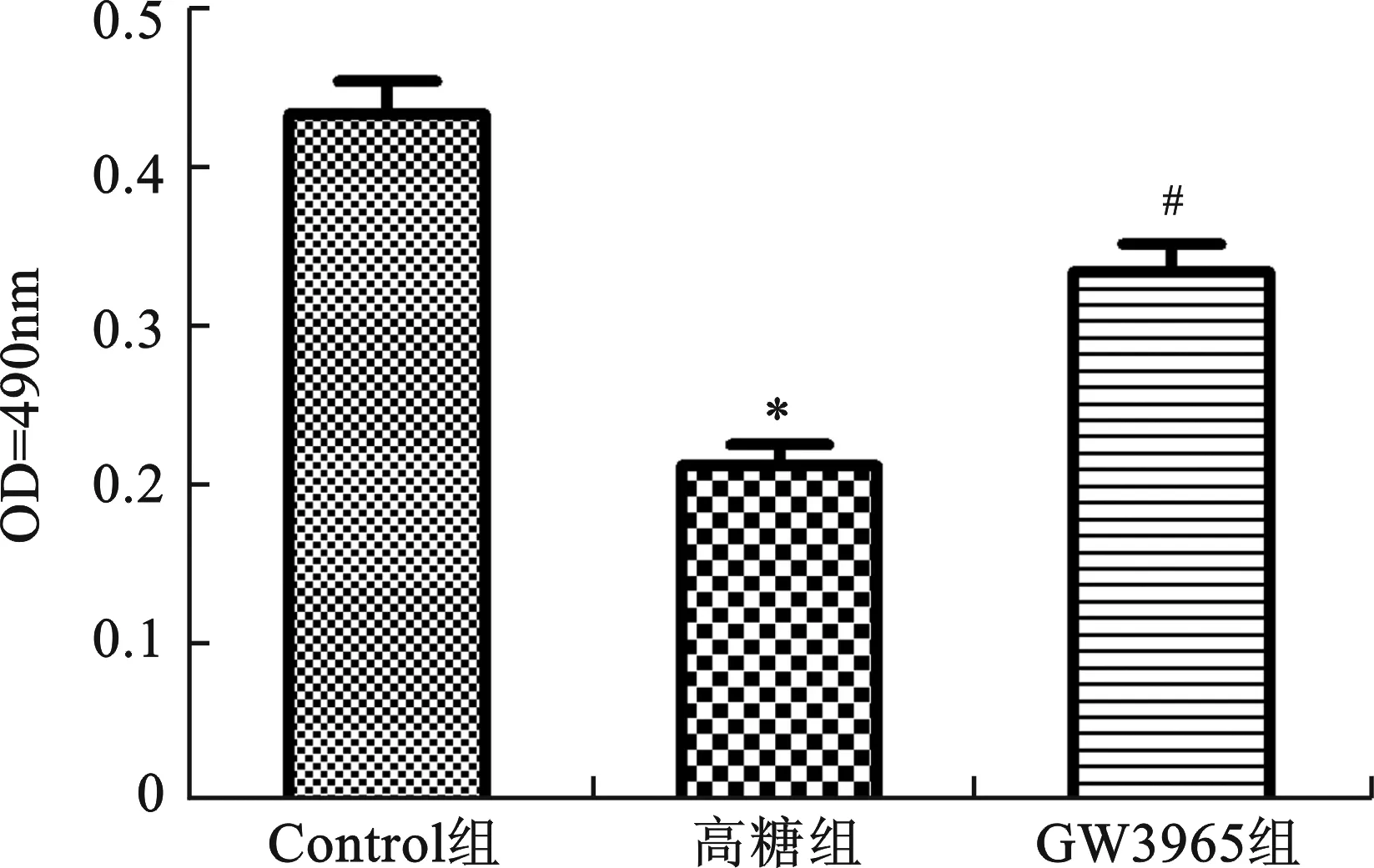
與Control組比較,*P<0.01;與高糖組比較,#P<0.01
3 線粒體膜電位檢測 線粒體膜電位降低是細胞凋亡的早期時間,因此早期細胞凋亡率可以代表線粒體膜電位變化。結果顯示,高糖組細胞凋亡率為(27.78±2.66)%,與Control組(3.22±1.71)%比較顯著升高,具有統計學差異(P<0.01),說明高糖誘導后H9c2細胞線粒體膜電位顯著下降;激動劑GW3965干預后,細胞凋亡率(11.73±1.65)%與高糖組比較顯著下降,具有統計學差異(P<0.01)。由此可見,GW3965可以抑制H9c2細胞線粒體膜電位下降。

圖2 Western blot檢測凝膠電泳圖
4 LXRα、MAPK通路蛋白檢測 結果顯示,同Control組比較,高糖組細胞內MAPK、ERK蛋白的表達無明顯變化,LXRα、p-p38 MAPK、p-ERK蛋白的表達顯著增加,差異具有統計學意義(P<0.01)。提示高糖處理H9c2細胞后,LXRα表達增加且MAPK信號通路被激活。采用激動劑GW3965處理后,同高糖組比較,LXRα蛋白的表達顯著升高,p-p38 MAPK、p-ERK蛋白的表達顯著下降,差異具有統計學意義(P<0.01,P<0.05)。由此可見,GW3965能夠促進LXRα表達,并有效抑制MAPK信號通路活性(圖2、3)。

與Control組比較,*P<0.01;與高糖組比較,#P<0.05,ΔP<0.01
討 論
糖尿病心肌病(DCM)是一種特異性心肌病,發生于糖尿病患者心肌細胞原發性受損傷所引起廣泛的心肌組織結構異常,最終表現為左心室肥厚、心肌收縮和(或)舒張功能障礙[7]。DCM能夠誘導患者心功能異常,引發心肌缺血或心力衰竭,是臨床糖尿病患者死亡的一個關鍵原因,發病機制錯綜復雜,目前尚不完全清楚。細胞凋亡是有機體由基因調控的細胞自由性死亡過程,又稱細胞程序性死亡[8]。心肌細胞凋亡已經被業界證明是DCM病變的重要特點,一直備受關注。持續凋亡會引起心肌肥大、間質纖維化,使心肌組織受損,并進一步誘導心功能障礙[7],最終導致心理衰竭。因此有效抑制細胞凋亡被認為有利于DCM的治療。
MAPK是生物體內重要信號轉導通路之一,參與調控細胞增殖、生長、分化、分裂以及凋亡等多種生物學過程,主要包括JNK、ERK和p38三條途徑。研究發現[9],p-p38 MAPK和p-ERK1/2的表達在糖尿病大鼠心臟中大量表達,并伴隨心肌細胞凋亡。在2型糖尿病誘導的DCM大鼠模型中,曲美他嗪通過抑制ERK和p38 MAPK的磷酸化減少心肌纖維化,抑制心肌細胞凋亡[10]。此外,T0901317激活的LXRs可以調控MAPK通路活性,進而減輕腦出血造成的神經元細胞凋亡和腦損傷[11]。提示MAPK通路參與調控心肌細胞凋亡,很可能受LXRs表達的影響。
LXRs屬于核受體超家族成員,通過結合特異性反應原件LXRE調節靶基因的表達,包括LXRα和LXRβ兩種亞型。LXRα高表達于肝臟、腎臟、脂肪和腸等代謝活躍的組織,在膽固醇代謝、糖脂代謝、炎癥反應和天然免疫中扮演重要角色。研究證實,LXRs在炎癥反應、糖脂代謝、氧化應激和細胞凋亡中發揮重要調控作用,是人體中潛在的藥理學靶受體。腹腔注射LXRs激動劑GW3965能夠抑制心肌缺血再灌注引起的氧化應激、內質網應激,減輕線粒體功能障礙和心肌細胞凋亡[12]。人工合成的肝X受體T0901317能夠降低ROS水平,恢復ΔΨm,抑制Cyt-C釋放,降低促凋亡蛋白Bax的表達,減輕高糖誘導的H9c2細胞凋亡[3]。此外,GW3965在2型糖尿病小鼠模型中亦被證明能夠改善心肌功能障礙[6]。但被GW3965激活的LXRα是否可以抑制高糖誘導的心肌細胞凋亡目前尚未證實。因此本研究對GW3965在高糖誘導的H9c2細胞凋亡中作用和機制進行初步探索。
本實驗采用高糖誘導H9c2建立體外細胞模型,同時加入LXRs激動劑GW3965進行干預。采用MTT實驗檢測細胞存活率,流式細胞儀分析細胞凋亡率,JC-1熒光染料法檢測線粒體膜電位,Western blot檢測LXRα和MAPK通路相關蛋白的表達。結果顯示,高糖誘導心肌細胞凋亡,抑制細胞增殖,GW3965可以顯著提高H9c2細胞的存活率,降低高糖誘導的H9c2細胞凋亡,差異具有統計學意義。說明LRXs激動劑能夠減輕高糖對心肌細胞造成的損傷,進一步證明了這類藥物對心臟功能的保護效應。本實驗還顯示,高糖引起心肌細胞線粒體膜電位下降,采用GW3965干預后H9c2細胞線粒體膜電位顯著升高,差異具有統計學意義。由此可見,GW3965對心肌細胞的保護作用同線粒體膜電位升高有關。此外,高糖作用于H9c2細胞后,LXRα的表達顯著下降,p38 MAPK和ERK信號通路被激活,而GW3965能夠增加LXRα的表達,減少p-p38 MAPK、p-ERK的表達,有效抑制p38 MAPK和ERK信號通路活性。
綜上所述,GW3965激活的LXRs對心肌細胞具有保護作用,通過抑制ERK、p38 MAPK通路的活性提高線粒體膜電位,改善線粒體功能障礙,有效抑制心肌細胞早期凋亡。說明LXRs很可能是糖尿病心肌病臨床治療的潛在藥理學靶受體。但本研究只是從細胞水平初步探討了GW3965改善高糖誘導心肌細胞損傷的分子機制,后續仍需在動物水平進一步研究其對DCM大鼠心室功能的影響,從而為臨床治療提供充分的理論依據。
[1] Codinach Huix P, Freixa Pamias R. Diabetic cardiomyopathy: concept, heart function, and pathogenesis[J]. An Med Interna, 2002, 19(6):313-320.
[2] Krum H, Gilbert RE. Demographics and concomitant disorders in heart failure [J]. Lancet, 2003, 362(9378):147-158.
[3] Cheng Y, Feng Y, Zhu M,etal. Synthetic liver X receptor agonist T0901317 attenuates high glucose-induced oxidative stress, mitochondrial damage and apoptosis in cardiomyocytes [J]. Acta Histochem, 2014, 116(1):214-221.
[4] Viennois E, Mouzat K, Dufour J,etal. Selective liver X receptor modulators (SLiMs): what use in human health? [J]. Mol Cell Endocrinol, 2012, 351(2):129-141.
[5] Lei P, Baysa A, Nebb HI,etal. Activation of Liver X receptors in the heart leads to accumulation of intracellular lipids and attenuation of ischemia-reperfusion injury [J]. Basic Res Cardiol, 2013, 108(1):323.
[6] He Q, Pu J, Yuan A,etal. Liver X receptor agonist treatment attenuates cardiac dysfunction in type 2 diabetic db/db mice [J]. Cardiovasc Diabetol, 2014, 13(149).
[7] 趙莉平, 安 榮, 丁 雯,等. 糖心樂對糖尿病心肌病大鼠心功能及心肌細胞凋亡的影響 [J]. 陜西中醫, 2014, 35(5):615-618.
[8] 安 榮,孔 維,趙莉平,等.糖心樂對糖尿病心肌病大鼠心肌Bnip-3、Bcl-2表達的影響[J].陜西中醫,2013,34(9):1248-1251.
[9] Soetikno V, Sari FR, Sukumaran V,etal. Curcumin prevents diabetic cardiomyopathy in streptozotocin-induced diabetic rats: possible involvement of PKC-MAPK signaling pathway [J]. Eur J Pharm Sci, 2012, 47(3):604-614.
[10] Zhang L, Ding WY, Wang ZH,etal. Early administration of trimetazidine attenuates diabetic cardiomyopathy in rats by alleviating fibrosis, reducing apoptosis and enhancing autophagy [J]. J Transl Med, 2016, 14(1):109.
[11] Wu CH, Chen CC, Lai CY,etal. Treatment with TO901317, a synthetic liver X receptor agonist, reduces brain damage and attenuates neuroinflammation in experimental intracerebral hemorrhage [J]. J Neuroinflammation, 2016, 13(1):62.
[12] He Q, Pu J, Yuan A,etal. Activation of liver-X-receptor alpha but not liver-X-receptor beta protects against myocardial ischemia/reperfusion injury [J]. Circ Heart Fail, 2014, 7(6):1032-1041.
(收稿:2017-01-10)
The effect of liver X receptors agonist GW3965 on myocardial apoptosis induced by high glucose and its mechanism
Cheng Yongxia, Zhang Dawei,Sun Lixin,et al.
Pathology Teaching and Research Section,Mudanjiang Medical University (Mudanjiang 157011)
Objective: To explore the effect of GW3965 on myocardial apoptosis induced by high glucose and its mechanism. Methods: H9c2 myocardial cells were randomly divided into three groups: control group, high glucose group and GW3965 group. Cells in GW3965 group were pretreated with 10μmol/L GW3965 for 24 h. Then cultures in high glucose group and GW3965 group were induced by 33 mmol/L glucose for 24 h. MTT was used to detect cells survival. The apoptosis was detected by flow cytometry and mitochondrial membrane potential by JC-1 fluorescence staining. Western blot was used to detect the expression of proteins related to LXR, ERK and p38 MAPK signal pathway. Results: GW3965 could inhibit the apoptosis of H9c2 cells, increase the rate of cells survival and influence mitochondrial membrane potential. GW3965 inhibited the activity of ERK and p38 MAPK pathway. Conclusion: GW3965 can inhibit the apoptosis and mitochondrial damage induced by high glucose via regulating LXRα and MAPK pathway, resulting a protective effect on myocardial cells.
Myocardium @GW3965 Hyperplycemia Diabetic cardiomyopathies/physiopathology @Liver X receptors Apoptosiss
*國家青年基金資助項目(81500629)
心肌 @GW3965 高血糖癥 糖尿病心肌病/病理生理學 @肝X受體 細胞凋亡
R392.3
A
10.3969/j.issn.1000-7377.2017.07.001
△通訊作者
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
