ABCG1在腫瘤壞死因子α誘導的氧化應激中的機制研究*
2017-08-28 16:38:51薛嘉虹朱參戰胡艷超欒春紅
中國現代醫學雜志 2017年11期
關鍵詞:氧化應激
薛嘉虹,朱參戰,胡艷超,欒春紅
(西安交通大學醫學院第二附屬醫院 心內科,陜西 西安710004)
ABCG1在腫瘤壞死因子α誘導的氧化應激中的機制研究*
薛嘉虹,朱參戰,胡艷超,欒春紅
(西安交通大學醫學院第二附屬醫院 心內科,陜西 西安710004)
目的 探討三磷酸腺苷結合盒轉運體G1(ABCG1)在腫瘤壞死因子α(TNF-α)誘導的氧化應激中的作用及可能的機制。方法 人臍靜脈內皮細胞被特異性ABCG1 siRNA或ABCG1過表達質粒轉染或使用LXR(肝X受體)激活劑T0901317預處理,隨后給予腫瘤壞死因子(TNF-α)干預12 h。采用DCFHDAAM(2’7’-二氯熒光雙乙酸鹽)熒光探針檢測細胞內活性氧簇(ROS)的水平,分光光度儀測量還原型煙酰胺腺嘌呤二核苷酸磷(NADPH)氧化酶活性,實時熒光定量聚合酶鏈反應法(qRT-PCR)和Western blot檢測內皮細胞NADPH氧化酶亞型非吞噬細胞氧化酶4(NOX4)表達及超氧化物歧化酶(SOD)的表達。結果 ABCG1表達上調抑制TNF-α誘導的氧化應激,同時抑制促氧化應激的NADPH氧化酶的活性和NOX4的表達,促進抗氧化的SOD表達。相反,ABCG1表達下調進一步誘導ROS的產生,誘導NADPH氧化酶的活性和NOX4的表達,抑制SOD1表達。結論 ABCG1通過調節NADPH氧化酶/SOD抑制TNF-α誘導的氧化應激。
三磷酸腺苷結合盒轉運體G1;氧化應激;還原型煙酰胺腺嘌呤二核苷酸磷氧化酶;超氧化物歧化酶
研究表明,氧化應激以及在氧化應激過程中產生的活性氧(reactive oxygen species,ROS)與多種心血管疾病的發生、發展有著密切關系[1-4]。內源性ROS產生除了主要來源于線粒體呼吸鏈外,還原型煙酰胺腺嘌呤二核苷酸磷(reduced nicotinamide adenine dinucleotide phosphate,NADPH)氧化酶是體內ROS產生的另一主要來源[2-4]。此外,機體也存在著清除過多產生ROS的抗氧化系統[3],當氧化和抗氧化作用失衡,即可導致大量ROS的產生。
ABCG1屬于三磷酸腺苷結合盒轉運體G1(adenosine triphosphate binding cassette transporter G1,ABCG1)家族成員之一(矛盾),具有調節細胞內游離膽固醇流出,防止泡沫細胞形成的作用[5-7]。此外,有研究發現,ABCG1也在內皮細胞高度表達,其促進內皮細胞內固醇流出的作用對保護內皮細胞正常功能具有十分重要的作用[8-11]。既往研究中,發現ABCG1表達上調可以降低TNF-α誘導的血管內皮細胞損傷,其作用可能與抑制腫瘤壞死因子-α(tumour necrosis factor-α,TNF-α)誘導的血管內皮細胞氧化應激相關[12]。本研究擬進一步探討ABCG1抑制TNF-α誘導的氧化應激可能涉及的機制。
1 材料與方法
1.1 細胞培養及實驗干預
人臍靜脈內皮細胞(human umbilical vein endothelial cells,HUVECs)購自美國 ATCC 細胞庫。內皮細胞培養在含10%胎牛血清的低糖培養基(dulbecco's modified eagle medium,DMEM)中,37℃、5%二氧化碳CO2培養箱中靜置培養。培養的HUVECs給予10 ng/ml的TNF-α干預12 h,或者預先使用肝X受體(liver X receptors,LXRs)配體T0901317(5μg/ml,Sigma)預處理HUVECs 2 h,或HUVECs被轉染特異性ABCG1 siRNA或ABCG1過表達質粒,24 h后,細胞再給予10 ng/ml TNF-α干預12 h。正常培養的HUVECs為對照組。
1.2 核糖核酸提取及逆轉錄和實時熒光定量聚合酶鏈反應
收集干預結束后的HUVECs,用Trizol(美國in vitrogen公司)一步法提取細胞總核糖核酸(ribonucleic acid,RNA)。以紫外分光光度計測定總RNA的純度(A260/A280)>1.8。取2μg總RNA逆轉錄成互補脫氧核糖核酸(complementary deoxyribonucleic acid,cDNA)。采用 SYBR Green I嵌合熒光法,以 3-磷酸甘油脫氫酶(glyceraldehyde-3-phosphate dehydrogenase,GAPD)基因為內參照,實時熒光定量聚合酶鏈反應(quantitative real time polymerase chain reaction,qRT-PCR)檢測HUVECs中NADPH氧化酶亞型NOX4及超氧化物歧化酶(superoxide dismutase,SOD) 信使核糖核酸 (messenger ribonucleic acid,mRNA)的表達。引物的序列如下:NOX4正向引物5'-CTGGTGAATGCCCTCAACTT-3',反向引物:5'-GGCCAGGAACAGTTGTGAAG-3';SOD1 正 向 引物:5'-TAGCGAGTTATGGCGACGAA-3',反向引物:5'-TGCTGTATTATCTCCAAACT-3';SOD2 正 向 引物:5'-GCCCTGGAACCTCACATCA-3',反向引物:5'-TGACCACCACCATTGAACTT-3';GAPDH 正向引物:5'-TCATCCCTGCCTCTACTG-3',反向引物:5'-T GCTTCACCACCTTCTTG-3'。反應條件為:95℃溫育10 s,95℃ 5 s,54℃ 20 s,讀板溫度;72℃ 12 s共 40個循環。每個反應做3個復孔。擴增完畢后行熔解曲線分析,確定擴增產物的特異性。因目的基因與內參照基因的擴增效率一致,qRT-PCR統計分析采用2-ΔΔCt法。ΔΔCt=對照組△Ct(目的基因Ct-管家基因Ct)-各組△Ct(目的基因Ct-管家基因Ct)。
1.3 Western blot免疫蛋白檢測
收集干預結束后的HUVECs,加入RIPA裂解液進行細胞裂解,于4℃離心10 min,棄除沉淀,用BCA法進行蛋白質定量。取50 g蛋白質加入1×上樣緩沖液中,在100℃加熱10 min使蛋白質變性。用10%十二烷基硫酸鈉(sodium dodecyl sulphate,SDS)聚丙烯酰胺凝膠(polyacrylamide gel electrophoresis,PAGE)進行電泳分離,轉硝酸纖維素膜(nitrocellulose filter membrane,NC),封閉液封閉 2 h,按說明書以1∶200加入兔抗人NOX4(美國Santa Cruz Biotechnology公司),兔抗人ABCG1(美國Santa Cruz Biotechnology 公司),4℃培育過夜,TBST(Tris-Hcl)緩沖溶液+吐溫洗3次,1∶2 000加入辣根過氧化物酶標記山羊抗兔二抗1∶5 000(美國Santa CruzBiotechnology公司),室溫培育1h,TBST洗3次,用增強化學發光液(enhanced chemilumincscence,ECL)(美國Amersham公司),冷電荷耦合元件(chargecoupled device,CCD) 照相機直接拍攝,Quality one軟件分析。β-actin(美國Santa Cruz Biotechnology公司)為對照組。
1.4 細胞內活性氧檢測
干預結束后的HUVECs和20 mmol DCFH-DA熒光探針(美國Sigma-Aldrich公司)37℃孵育20min,磷酸鹽緩沖溶液(phosphate buffered solution,PBS)清洗3次,熒光顯微鏡測定細胞內熒光素的強度以表示細胞內ROS產生的多少。
1.5 NADPH氧化酶活性檢測
根據細胞NADPH氧化酶活性光度法定量檢測試劑盒說明書進行(上海一基實業有限公司)。樣品準備好后,根據說明書依次加入緩沖液、反應液、底物液,即刻放入分光光度儀,在λ=340nm處測定NADPH氧化酶的活性,結果為樣品總活性-樣品非特異性活性。
1.6 ABCG1干擾和過表達
參考既往實驗[13],選用化學合成的ABCG1干擾序列正向引物:5'-GAGUCUUUCUUCGGGAACATT-3',反向引物:5'-UGUUCCCGAAGAAAGACUCTT-3'(上海吉瑪生物制藥公司)。ABCG1 siRNA和隨機siRNA序列使用TurboFect siRNA轉染試劑(立陶宛Fermentas公司)轉染HUVECs,轉染24 h后細胞用于隨后的實驗干預。綠色熒光標記的ABCG1過表達質粒EX-Z0509-M61和對照載體EX-EGFP-M61由美國Gene CopoeiaTM公司設計合成并轉染HUVECs,Western blot驗證轉染效率,結果見[13]。
1.7 統計學方法
數據分析采用SPSS 19.0統計軟件,計量資料以均數±標準差(±s)表示,多組間比較采用單因素方差分析,兩兩比較用LSD-t檢驗,P<0.05為差異有統計學意義。
2 結果
2.1 ABCG1表達改變對TNF-α誘導的內皮細胞氧化應激的影響
TNF-α干預HUVECs后,內皮細胞內熒光強度較對照組增加,而使用T0901317預處理HUVECs組,內皮細胞內熒光強度則減弱,提示使用T0901317后內皮細胞內活性氧產生減少。當進一步使用ABCG1質粒轉染HUVECs,使內皮細胞ABCG1表達增高后,再給予TNF-α干預,內皮細胞內熒光強度類似T0901317干預組,同樣較單純TNF-α干預的內皮細胞熒光強度減弱。相反,使用ABCG1 siRNA轉染HUVECs后,結果發現細胞內熒光強度較單純TNF-α干預的內皮細胞有所增強,提示活性氧進一步產生。見圖1。

圖1 ABCG1表達改變對TNF-α誘導的內皮細胞氧化應激的影響 (×100)
2.2 ABCG1表達改變對NADPH氧化酶活性的影響
在TNF-α干預組,HUVECs的ABCG1表達較對照組降低(t=31.00,P=0.001),NADPH 氧化酶活性則較對照組增加(t=30.10,P=0.001),使用 T0901317預處理或ABCG1質粒轉染細胞后再給予TNF-α干預,結果發現,隨著內皮細胞ABCG1表達增加,NADPH氧化酶活性被逆轉,較TNF-α干預組降低(T0901317干預組 t=27.58,P=0.001,ABCG1質粒轉染組t=29.76,P=0.001),幾乎接近對照組的水平。在使用ABCG1 siRNA轉染組,ABCG1表達進一步降低,內皮細胞NADPH氧化酶活性較對照組增加(FABCG1表達=12.858,PABCG1表達=0.000;FNADPH氧化酶活性=700.108,PNADPH氧化酶活性=0.000)。見圖 2。

圖2 ABCG1表達改變對NADPH mRNA氧化酶活性的影響
2.3 ABCG1表達改變對NOX4表達的作用NOX4作為NADPH氧化酶亞單位之一,與NADPH氧化酶的其他亞單位相比較,主要表達在內皮細胞。本實驗結果提示,在TNF-α干預組,HUVECs的NOX4 mRNA表達和蛋白表達較對照組增加(t=48.00 和22.25,均P=0.000),而使用T0901317預處理或ABCG1質粒轉染細胞后再給予TNF-α干預,HUVECs的NOX4 mRNA表達和蛋白表達均較TNF-α干預組降低(在mRNA表達方面,tT0901317組=26.50,PT0901317組=0.000,tABCG1質粒轉染組=19.33,PABCG1質粒轉染組=0.000;蛋白表達方面 tT0901317組=24.50,PT0901317組=0.000;tABCG1質粒轉染組=21.60,PABCG1質粒轉染組=0.000)。使用ABCG1 siRNA轉染細胞后,內皮細胞NOX4 mRNA表達和蛋白表達較對照組增加(tNOX4mRNA表達=262.519,PNOX4mRNA表達=0.000;tNox4蛋白表達=304.004,PNOX4蛋白表達=0.000)。但與 TNF-α 干預組比較,差異無統計學意義(t=2.25和2.00,均P>0.05)。見圖3。
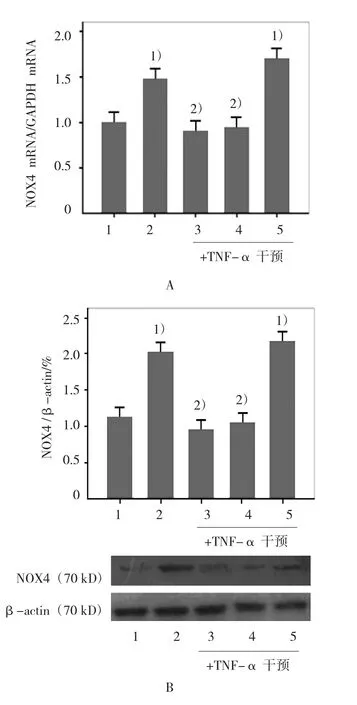
圖3 ABCG1表達改變對NOX4 mRNA和蛋白表達的影響
2.4 ABCG1表達改變對SOD mRNA表達的影響
SOD是體內重要的抗氧化酶。當TNF-α干預HUVECs后,HUVECs的SOD1和SOD2 mRNA表達較對照組降低(t=3.62 和 3.10,均 P <0.05),而使用T0901317預處理或ABCG1質粒轉染細胞后,隨著ABCG1表達增加,內皮細胞的SOD1和SOD2 mRNA表達較TNF-α干預組增加(FSOD1mRNA=800.119,PSOD1mRNA=0.000;FSOD2mRNA=504.138,PSOD2mRNA=0.000)。當使用ABCG1 siRNA轉染內皮細胞使ABCG1表達降低后,內皮細胞SOD1 mRNA表達較TNF-α干預組進一步降低(t=3.22,P <0.05),而 SOD2 mRNA表達較TNF-α干預組輕微降低,但差異無統計學意義(t=2.051,P=0.06)。見圖 4。

圖4 ABCG1表達改變對SOD1和SOD2 mRNA表達的影響
3 討論
氧化應激為各種原因導致機體內氧化與抗氧化之間平衡失調,活性氧簇產生過多,使機體處于促氧化狀態,引發各種各樣心血管疾病。當前研究發現,炎癥因子TNF-α能促進體外培養的內皮細胞的氧化應激產生,而作為脂質轉運體的ABCG1可以通過影響NADPH氧化酶的促氧化作用和SOD的抗氧化作用從而調節TNF-α誘導的氧化應激的產生。
研究表明,ABCG1是促進細胞內膽固醇流出的重要膜轉運體之一,能防止泡沫細胞形成,并有防止內皮細胞激活,改善內皮功能失調的作用[5-11]。ABCG1表達增加可以抑制內皮細胞炎癥因子如TNF-α、IL-6及ICAM等的釋放[9-10],從而有助于防治動脈粥樣硬化的發生、發展。本研究進一步發現,ABCG1表達增加可抑制內皮細胞氧化應激的產生[11-12]。通過使用T0901317預處理內皮細胞,結果發現T0901317能降低TNF-α誘導的內皮細胞氧化應激。由于T0901317作為肝X受體配體,具有促進脂質轉運體ABCG1表達的作用[5],因此提示T0901317干預后的抗氧化應激的作用可能與促進ABCG1表達相關,進一步通過ABCG1轉染使ABCG1表達增加也發現內皮細胞氧化應激明顯降低,而ABCG1干擾使ABCG1表達抑制則促進炎癥因子誘導的內皮細胞氧化應激。正如TERASAKA[8]和TABET等[14]的研究所示,高密度脂蛋白降低氧化應激的作用依賴于ABCG1的表達,間接提示ABCG1具有抑制氧化應激的作用。而筆者既往研究發現,ABCG1能調節高糖誘導的氧化應激[11],進一步提示促進ABCG1表達可能具有抑制多種原因誘導的內皮細胞氧化應激產生的作用,但機制尚不明確。
雖然線粒體電子傳遞鏈及黃嘌呤氧化酶、一氧化氮合酶和NADPH氧化酶等均可產生ROS,但研究顯示,NADPH氧化酶是人血管系統中產生的ROS主要來源之一[2-4]。其中NOX4可能作為血管內皮細胞中主要的NADPH氧化酶參與內皮細胞內ROS的生成。研究顯示,TNF-α作為體內重要的炎癥因子,可以通過激活NADPH氧化酶促進氧化應激的產生[15]。本研究同樣發現,TNF-α能促進NADPH氧化酶的活性和表達增加,ABCG1表達改變不僅影響著NADPH氧化酶的活性,還影響著NOX4的基因和蛋白表達,從而提示ABCG1可能通過抑制血管內皮細胞中NADPH氧化酶的表達和活性發揮抗氧化應激的作用。
另一方面,機體是一個有機的整體,當氧化應激產生過多時,抗氧化系統可代償性激活,以減少ROS的產生[3]。體內已知的抗氧化系統包括抗氧化酶類系統和非酶類抗氧化劑,其中SOD是體內重要的抗氧化酶,可將毒性高的ROS轉換為毒性較低的過氧化氫(hydrogen peroxide,H2O2),隨后由谷胱甘肽過氧化物酶和H2O2酶繼續作用,使H2O2轉變成完全無害的水(H2O)和氧氣(Oxygen,O2)。SOD 有 3種亞型,其中有核細胞主要表達SOD1和SOD2。本研究發現,TNF-α抑制抗氧化的SOD的表達,而ABCG1表達增加促進TNF-α干預下內皮細胞SOD1和SOD2基因的表達,而ABCG1表達降低則抑制內皮細胞SOD1基因的表達,SOD2表達輕微降低,但差異無統計學意義。提示ABCG1可能通過調節血管內皮細胞中抗氧化的SOD的表達進一步發揮抗氧化應激的作用。
總之,本研究提示ABCG1具有拮抗TNF-α誘導的氧化應激的作用,其機制可能涉及調節NADPH氧化酶/SOD的促氧化應激和抗氧化應激間的平衡。期望今后的臨床治療將促進ABCG1表達作為新型的抗氧化措施,為心血管疾病提供新的防治方案。
[1]MADAMANCHI N R,RUNGE M S.Redox signaling in cardiovascular health and disease[J].Free Radical Biology&Medicine,2013,61(8):473-501.
[2]DAN S,WEISS D,LASSèGUE B,et al.Superoxide production and expression of nox family proteins in human atherosclerosis[J].Circulation,2002,105(12):1429-1435.
[3]BO C,LU Y,CHEN Y,et al.The role of Nrf2 in oxidative stress-induced endothelial injuries[J].Journal of Endocrinology,2015,225(3):83-99.
[4]GARCíA-REDONDO A B,AGUADO A,BRIONES A M,et al.NADPH oxidases and vascular remodeling in cardiovascular diseases[J].Pharmacological Research,2016,114(10):110-120.
[5]DEMINA E P,MIROSHNIKOVA V V,SCHWARZMAN A L.Role of the ABC transporters A1 and G1,key reverse cholesterol transport proteins,in atherosclerosis[J].Molecular Biology,2016,50(2):223-230.
[6]MAULDIN J P,NAGELIN M H,WOJCIK A J,et al.Reduced expression ofATP-binding cassette transporterG1 increases cholesterol accumulation in macrophages of patients with type 2 diabetes mellitus[J].Circulation,2008,117(21):2785-2792.
[7]XUE J H,YUAN Z,WU Y,et al.High glucose promotes intracellular lipid accumulation in vascular smooth muscle cells by impairing cholesterol influx and efflux balance[J].Cardiovascular Research,2010,86(1):141-150.
[8]TERASAKA N,YU S,YVAN-CHARVET L,et al.ABCG1 and HDL protect against endothelial dysfunction in mice fed a highcholesterol diet[J].Journal of Clinical Investigation,2008,118(11):3701-3713.
[9]WHETZEL A M,STUREK J M,NAGELIN M H,et al.ABCG1 deficiency in mice promotes endothelial activation and monocyteendothelial interactions[J].Arteriosclerosis Thrombosis&Vascular Biology,2010,30(4):809-817.
[10]XUE S,WANG J,ZHANG X,et al.Endothelial ATP-binding cassette G1 in mouse endothelium protects against hemodynamic-induced atherosclerosis[J].Biochemical& Biophysical Research Communications,2016,477(2):247-254.
[11]XUE J,WANG C,ZHU C,et al.ATP-binding cassette transporter G1 protects against endothelial dysfunction induced by high glucose[J].Diabetes Research&Clinical Practice,2013,101(1):72-80.
[12]薛嘉虹,宋安齊,李歡,等.三磷酸腺苷結合盒轉運體G1表達上調降低腫瘤壞死因子-α誘導的血管內皮細胞損傷[J].中國現代醫學雜志,2013,23(5):1-5.
[13]XUE J,WEI J,DONG X,et al.ABCG1 deficiency promotes endothelial apoptosis by endoplasmic reticulum stress-dependent pathway[J].The Journal of Physiological Sciences,2013,63(6):435-444.
[14]TABET F,LAMBERT G,CUESTA TORRES L F,et al.Lipidfree apolipoprotein A-I and discoidal reconstituted high-density lipoproteins differentially inhibit glucose-induced oxidative stress in human macrophages[J].Arteriosclerosis Thrombosis&Vascular Biology,2011,31(5):1192-1200.
[15]LIN C P,HUANG P H,LAI C F,et al.Simvastatin attenuates oxidative stress,NF-κB activation,and artery calcification in LDLR-/-mice fed with high fat diet via down regulation of tumor necrosis factor-α and TNF receptor 1[J].PloS One,2015,10(12):e0143686.
ABCG1 inhibits TNF-α-induced oxidative stress by regulating NADPH oxidase and superoxide dismutase*
Jia-hong Xue,Can-zhan Zhu,Yan-chao Hu,Chun-hong Luan
(Department of Cardiovascular Medicine,the Second Affiliated Hospital of College of Medicine,Xi'an Jiaotong University,Xi'an,Shaanxi 710004,China)
ObjectiveTo explore the role of ATP-binding cassette sub-family G member 1 (ABCG1)in oxidative stress production induced by tumor necrosis factor α (TNF-α)and its possible mechanisms.MethodsHuman umbilical vein endothelial cells (HUVECs)were transfected with specific ABCG1 siRNA or ABCG1 overexpression plasmid or pretreated with liver X receptor agonist T0901317,then were cultured with TNF-α for 12 hours.Intracellular reactive oxygen species (ROS)levels were measured using 6-carboxy-2,7-dichlorodihydrofluorescein diacetate,diacetoxymethyl ester(CDCFHDA-AM)fluorescence and nicotinamide adenine dinucleotide phosphate-oxidase (NADPH oxidase)activity was measured by spectrophotometer.Real time PCR and Western blot were employed to measure the expression of Nox4,one of NADPH oxidase subunit and the expression of superoxide dismutase (SOD).ResultsABCG1 upregulation inhibited TNF-α-induced oxidative stress.Furthermore,the activity of NADPH oxidase and the expression of Nox4 were also suppressed by ABCG1 overexpression,but the expression of antioxidant SOD was promoted.Conversely,downregulation of ABCG1 by ABCG1 siRNA both increased the ROS production and promoted the NADPH oxidase activity and Nox4 expression.However,the expression of SOD1 was inhibited.ConclusionsThe results suggest thatABCG1 attenuates TNF-α-induced oxidative stress by regulating NADPH oxidase and SOD.
adenosine triphosphate binding cassette transporter G;oxidative stress;reduced nicotinamide adenine dinucleotide phosphate oxidase;superoxide dismutase
R363
A
10.3969/j.issn.1005-8982.2017.11.003
1005-8982(2017)11-0014-06
2016-10-27
國家自然科學基金(No:81100210)
猜你喜歡
中成藥(2021年5期)2021-07-21 08:39:04
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
中成藥(2018年6期)2018-07-11 03:01:24
中成藥(2018年5期)2018-06-06 03:11:43
天然產物研究與開發(2016年6期)2016-06-05 10:29:26
西南軍醫(2016年6期)2016-01-23 02:21:19
新疆醫科大學學報(2015年10期)2015-12-26 12:33:30
吉林大學學報(醫學版)(2015年4期)2015-12-17 07:48:13
實用中西醫結合臨床(2015年7期)2015-02-28 16:30:23
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
