油菜MADS-box家族基因AGL11的克隆、表達(dá)及轉(zhuǎn)化油菜的研究
2017-11-16 08:30:21李彥杰周大祥
西南農(nóng)業(yè)學(xué)報(bào) 2017年10期
關(guān)鍵詞:油脂
熊 書(shū),李彥杰,周大祥,3 *
(1.重慶三峽醫(yī)藥高等專(zhuān)科學(xué)校 基礎(chǔ)醫(yī)學(xué)部,重慶 萬(wàn)州 404120;2.重慶三峽學(xué)院 生命科學(xué)與工程學(xué)院,重慶 萬(wàn)州 404100;3.重慶大學(xué) 生命科學(xué)學(xué)院,重慶市基因功能與調(diào)控重點(diǎn)實(shí)驗(yàn)室,重慶 400300)
油菜MADS-box家族基因AGL11的克隆、表達(dá)及轉(zhuǎn)化油菜的研究
熊 書(shū)1,李彥杰2,周大祥2,3 *
(1.重慶三峽醫(yī)藥高等專(zhuān)科學(xué)校 基礎(chǔ)醫(yī)學(xué)部,重慶 萬(wàn)州 404120;2.重慶三峽學(xué)院 生命科學(xué)與工程學(xué)院,重慶 萬(wàn)州 404100;3.重慶大學(xué) 生命科學(xué)學(xué)院,重慶市基因功能與調(diào)控重點(diǎn)實(shí)驗(yàn)室,重慶 400300)
【目的】本研究為油菜種子中油脂合成分子調(diào)控機(jī)理提供新的線(xiàn)索。【方法】通過(guò)定量PCR分析油菜AGL11基因的組織表達(dá)模式,同時(shí)構(gòu)建AGL11的干擾載體。【結(jié)果】生物信息學(xué)分析發(fā)現(xiàn)油菜AGL11具有完整且典型的MADS-box保守結(jié)構(gòu)域,蛋白整體形態(tài)呈現(xiàn)典型的DNA核酸結(jié)合結(jié)構(gòu),屬于MADS-box家族轉(zhuǎn)錄調(diào)控因子,親源關(guān)系與擬南芥AGL11蛋白最近。定量PCR研究發(fā)現(xiàn),AGL11在油菜盛花期和果夾5DAP(盛花后5 d 的種子)表達(dá)量最高,實(shí)驗(yàn)成功構(gòu)建了RNA干擾載體。【結(jié)論】AGL11基因在油菜種子發(fā)育和油脂積累過(guò)程中強(qiáng)烈表達(dá),該基因可能在種子發(fā)育及油脂的合成中發(fā)揮著重要作用。RNA干擾載體的構(gòu)建成功將為后續(xù)獲得轉(zhuǎn)基因油菜,觀察轉(zhuǎn)基因油菜胚的發(fā)育和種子油脂的積累,從而確定AGL11在油菜胚的發(fā)育及油脂的合成中發(fā)揮著重要作用。
油菜;MADS-box;AGL11;RNAi;油脂合成
【研究意義】MADS-box基因是植物中一類(lèi)極為重要的轉(zhuǎn)錄調(diào)控因子,在調(diào)控生長(zhǎng)發(fā)育和信號(hào)傳導(dǎo)中發(fā)揮著重要的作用[1-3]。大量研究發(fā)現(xiàn)MADS-box家族基因AGLs參與植物花器官的發(fā)育[4-5],在果實(shí)的發(fā)育過(guò)程中也起著重要的作用[6-7]。影響種子發(fā)育的一些轉(zhuǎn)錄因子同時(shí)影響著種子中油脂的積累。LeafyCotyledon1,2(LEC1,LEC2),F(xiàn)usca3(FUS3)和WRINKLEDl(WRI1)等基因都是種子發(fā)育過(guò)程中油脂等物質(zhì)積累的關(guān)鍵調(diào)控因子。這些基因的突變、過(guò)量表達(dá)或異位表達(dá)都會(huì)對(duì)植物種子發(fā)育和油脂等物質(zhì)的積累產(chǎn)生重要的影響。作為CCAAT-Box結(jié)合因子HAP3的亞基之一,LEC1控制植物胚胎發(fā)育過(guò)程中的各個(gè)方面,是胚胎發(fā)育進(jìn)程中關(guān)鍵的調(diào)控因子。擬南芥LEC1基因的過(guò)量表達(dá)可影響其它種子發(fā)育相關(guān)的轉(zhuǎn)錄因子如FUS3和WRIl等基因的表達(dá),致使脂肪酸合成關(guān)聯(lián)基因的表達(dá)水平顯著提高,最終導(dǎo)致種子中脂肪酸和油脂的含量增加[8]。WRI1編碼一個(gè)AP2/EREB結(jié)構(gòu)域蛋白,在亞麻薺中超表達(dá)WRI1基因,可以提高種子質(zhì)量和油脂存儲(chǔ)[9]。【前人研究進(jìn)展】目前,AGLs調(diào)控植物油脂合成的研究較少,在擬南芥中,隸屬于 MADS-box 家族的 AGL15 對(duì)胚的發(fā)育也具有重要調(diào)控作用,它可以直接調(diào)控 LEC2、ABI3 及 FUS3基因的表達(dá),進(jìn)而影響胚的發(fā)育[10]。GmSEP1屬于 AGL2 /SEP 亞家族,GmSEP1 在大豆生殖器官花、莢和種子表達(dá),在四輪花器官中有不同程度的表達(dá),推測(cè)可能與大豆花器官特化和種子發(fā)育相關(guān)[11]。擬南芥基因 AGL66 和 AGL104 特異性調(diào)控花粉的成熟[12]。前期研究在油菜胚發(fā)育和油脂合成表達(dá)數(shù)據(jù)庫(kù)中發(fā)現(xiàn)油菜AGL11基因在胚中表達(dá)較高,初步認(rèn)為該基因可能與油脂合成有關(guān)。【本研究切入點(diǎn)】本實(shí)驗(yàn)通過(guò)克隆甘藍(lán)型油菜(BrassicanapusL.)的AGL11基因,同時(shí)構(gòu)建AGL11的干擾載體,為后續(xù)實(shí)驗(yàn)進(jìn)一步確定AGL11通過(guò)控制油脂合成相關(guān)基因的表達(dá),從而調(diào)節(jié)油脂的合成。【擬解決的關(guān)鍵問(wèn)題】本研究結(jié)果為探索油菜種子中油脂合成的分子調(diào)控機(jī)理提供新的線(xiàn)索,通過(guò)基因工程手段提高油菜油脂含量,為改良油菜品質(zhì)奠定基礎(chǔ)。
1 材料與方法
1.1 供試材料
甘藍(lán)型油菜Brassicanapuscv. Westar作為研究材料,實(shí)驗(yàn)所用的RNAi干擾質(zhì)粒和農(nóng)桿菌菌株GV3101由重慶大學(xué)基因工程中心提供。
1.2 試驗(yàn)方法
1.2.1 提取RNA與熒光定量PCR 以BnAGL11基因設(shè)計(jì)熒光定量PCR引物BnAGL11-F(5′-AACCGCAGTCCCGTTCTCTC-3′)和BnAGL11-R(5′-AGTCCATTTCTTCTTTTGCA-3′),引物由上海生工生物工程有限公司合成。利用Qiagen Plant RNA Kit試劑盒提取油菜各組織的RNA,利用Thermo Scientific First Strand cDNA Synthesis Kit試劑盒反轉(zhuǎn)錄獲得cDNA,-80 ℃保存?zhèn)溆谩晒舛縋CR利用TaKaRa公司SYBR Green Ⅰ試劑盒,在CFX96熒光定量PCR儀上進(jìn)行,分析BnAGL11基因在油菜不同組織的表達(dá)情況。反應(yīng)程序:95 ℃預(yù)變性20 s,95 ℃變性3 s,60 ℃退火30s,60 ℃延伸15 s,40個(gè)循環(huán)。每個(gè)樣品重復(fù)3次,數(shù)據(jù)采用2—△△Ct法計(jì)算。
1.2.2 生物信息學(xué)分析 各種植物相關(guān)基因和氨基酸序列都來(lái)自NCBI數(shù)據(jù)庫(kù),蛋白結(jié)構(gòu)域分析在http://pfam.wustl.edu/上進(jìn)行。利用ClustalX2.1比對(duì)蛋白質(zhì)序列,用MEGA 5構(gòu)建進(jìn)化樹(shù),利用SWISS MODEL(http://swiss model.expase.org)預(yù)測(cè)蛋白質(zhì)的三維結(jié)構(gòu)。
1.2.3 干擾載體構(gòu)建 以油菜盛花期的花cDNA為模板,用BnAGL11的正向前后引物(For-BnAGL11-F:5′-CAATTACTTTGCTCATAACA-3′;For-BnA GL11-R:5′-CAATTACTTTGCTCATAACA-3′)擴(kuò)增出正向基因片段For-BnAGL11,將此片段以及RNAi載體用XbaⅠ和SalⅠ酶切,純化回收后連接到RNAi載體上,轉(zhuǎn)化到大腸桿菌,酶切驗(yàn)證。反向前后引物(Rev-BnAGL11-F:5′-CAATTACTTTGCTCATAGCA-3′; Rev-BnAGL11-R:5′-ACCTTCGTTGTTTTTTGAA T-3′)操作同上。
1.3 數(shù)據(jù)分析
應(yīng)用SPSS19.0統(tǒng)計(jì)軟件對(duì)實(shí)驗(yàn)數(shù)據(jù)進(jìn)行t檢驗(yàn),P<0.05表示具有顯著性差異。
2 結(jié)果與分析
2.1 蛋白結(jié)構(gòu)分析
圖1表示,通過(guò)與擬南芥的AGL11蛋白結(jié)構(gòu)比對(duì)分析,發(fā)現(xiàn)甘藍(lán)型油菜的AGL11氨基酸序列與擬南芥同源性很高,二者都具有完整且典型的MADS-box保守結(jié)構(gòu)域。大量研究發(fā)現(xiàn)MADS-box家族基因參與植物花器官的發(fā)育,在果實(shí)的發(fā)育過(guò)程中起著重要的作用,推測(cè)AGL11蛋白編碼的基因?qū)儆贛ADS-box家族轉(zhuǎn)錄調(diào)控因子。

At表示擬南芥;Bn表示甘藍(lán)型油菜圖1 AGL11的氨基酸序列比對(duì)和同源性分析Fig.1 Amino acid sequence alignment and homology analysis of AGL11

圖2 SWISS MODEL三維蛋白質(zhì)結(jié)構(gòu)預(yù)測(cè)Fig.2 SWISS MODEL three-dimensional protein structure prediction
2.2 蛋白質(zhì)的三維結(jié)構(gòu)預(yù)測(cè)
通過(guò)SWISS MODEL在線(xiàn)蛋白質(zhì)三級(jí)結(jié)構(gòu)分析,發(fā)現(xiàn)該蛋白具有1個(gè)α螺旋,2個(gè)β片層結(jié)構(gòu),還有一些不規(guī)則卷曲,其整體形態(tài)呈現(xiàn)典型的DNA核酸結(jié)合結(jié)構(gòu)(圖2),也進(jìn)一步證明該基因編碼的蛋白為1個(gè)轉(zhuǎn)錄因子,具有典型MADS-box保守結(jié)構(gòu)域,屬于MADS-box家族基因。
2.3 進(jìn)化樹(shù)分析
為了分析油菜BnAGL11蛋白和擬南芥AGL蛋白的遺傳進(jìn)化關(guān)系,用NCBI下載的蛋白序列,采用MEGA程序建立了系統(tǒng)發(fā)育樹(shù)。從圖3可以看出,BnAGL11蛋白與AtAGL11,AtAGL1,AtAGL5,AtSHP2蛋白組成一個(gè)小的亞家族,與AtAGL11蛋白親源關(guān)系最近,也表明BnAGL11可能與AtAGL11在功能上可能有一定的相似性。
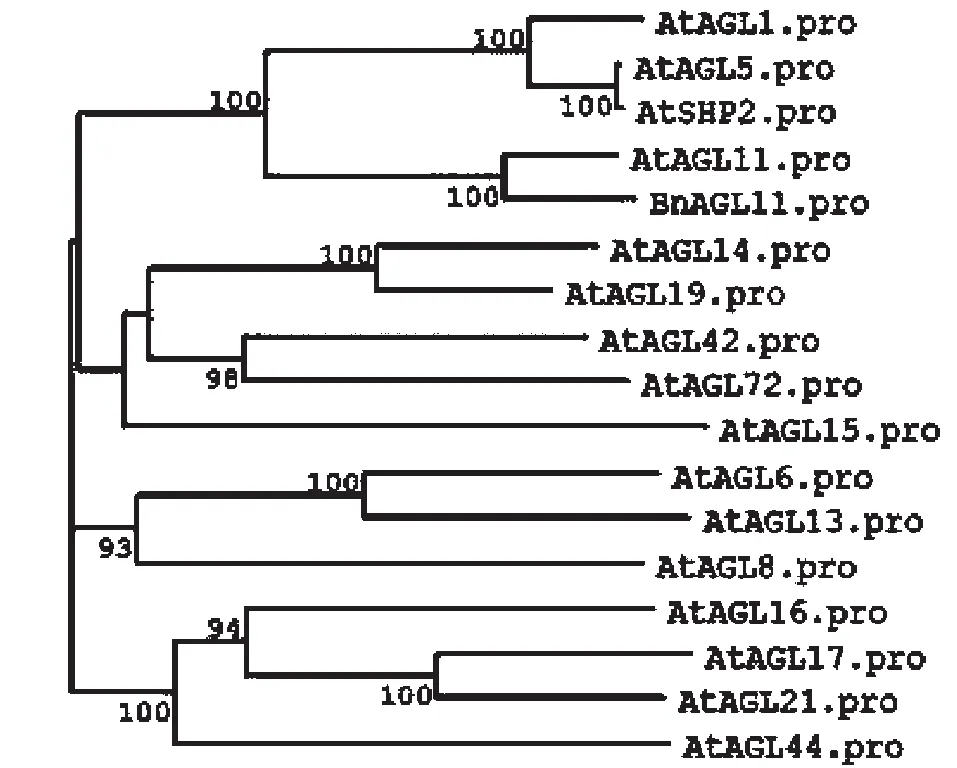
圖3 BnAGL11的系統(tǒng)發(fā)育樹(shù)Fig.3 Phylogenetic tree of BnAGL11
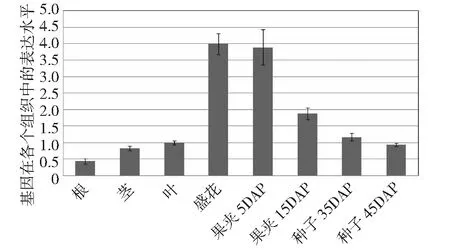
DAP表示盛花后幾天的種子圖4 甘藍(lán)型油菜AGL11基因的組織表達(dá)模式分析Fig.4 Analysis of tissue expression patterns of BnAGL11 gene
2.4 定量PCR表達(dá)分析
由圖4可知,AGL11在各個(gè)組織中均有表達(dá),其中在盛花期表達(dá)量最高,在果夾5DAP時(shí)期表達(dá)量接近盛花期,隨后逐漸降低,二者是其他組織表達(dá)量的2倍以上,油菜種子在DAP5和DAP15之間是胚迅速膨大、油脂快速的積累時(shí)期,定量PCR的結(jié)果說(shuō)明AGL11在油菜胚的發(fā)育過(guò)程及其油脂的合成中可能具有重要的作用。
2.5 干擾載體構(gòu)建及轉(zhuǎn)化油菜
為了進(jìn)一步驗(yàn)證AGL11在油菜胚的發(fā)育過(guò)程及其油脂的合成中可能具有重要的功能,采用RNAi干擾技術(shù),降低AGL11基因在油菜中的表達(dá),然后觀察轉(zhuǎn)基因油菜的表型。為了構(gòu)建RNAi干擾載體,首先以AGL11基因cDNA為模板,擴(kuò)增出了100 bp的干擾載體正向和反向片段(圖5A),然后分別插入干擾載體中,獲得重組質(zhì)粒,進(jìn)一步酶切驗(yàn)證插入片段大小,發(fā)現(xiàn)插入片段1000 bp(圖5B),與預(yù)期一致,證明RNAi載體構(gòu)建成功。隨后將載體導(dǎo)入農(nóng)桿菌GV310菌株中,采用農(nóng)桿菌介導(dǎo)的葉盤(pán)法,用油菜下胚軸和子葉為受體,進(jìn)行遺傳轉(zhuǎn)化,目前正在獲得陽(yáng)性抗性苗。隨后將進(jìn)一步獲得轉(zhuǎn)基因油菜植株,觀察轉(zhuǎn)基因油菜胚的發(fā)育和種子油脂的積累,確定AGL11在油菜胚的發(fā)育及油脂的合成中發(fā)揮著重要的作用。
3 討 論
油菜是全球重要的經(jīng)濟(jì)作物,同時(shí)在我國(guó)的油料作物中排名第一,是優(yōu)良植物蛋白和食用植物油的重要來(lái)源。統(tǒng)計(jì)顯示,我國(guó)食用植物油使用量的35 %來(lái)自于菜籽油,而植物餅粕使用量的25 %來(lái)自于菜籽餅粕,增加油菜產(chǎn)油率和油脂品質(zhì)改善始終是油菜育種的重要內(nèi)容[13]。據(jù)統(tǒng)計(jì),對(duì)油菜產(chǎn)油率而言,每提高1 %的油菜油脂含量,相當(dāng)于增加2.5 %的菜籽產(chǎn)量。現(xiàn)在,我國(guó)主要油菜品種含有40 %左右平均油脂含量,而國(guó)外一般油菜品種達(dá)到了42 %~45 %,部分甘藍(lán)型油菜品種油脂含量高達(dá)47 %。這必然會(huì)導(dǎo)致油菜種植業(yè)經(jīng)濟(jì)效益低下,打擊農(nóng)民種植油菜的積極性,制約了進(jìn)一步擴(kuò)大油菜種植面積和油菜業(yè)的發(fā)展。為此,進(jìn)一步研究油菜油脂合成的分子調(diào)控機(jī)理,了解控制油脂合成的關(guān)鍵基因,為通過(guò)基因工程手段提高油菜油脂含量,改良油菜品質(zhì),提高油菜產(chǎn)業(yè)生產(chǎn)效益奠定基礎(chǔ)。

圖5 RNA干擾載體的構(gòu)建Fig.5 Construction of RNAi vector
研究表明,過(guò)量表達(dá)或異位表達(dá)一些轉(zhuǎn)錄因子,可以對(duì)植物種子發(fā)育和油脂等物質(zhì)的積累產(chǎn)生重要的影響。如擬南芥LEC1基因的過(guò)量表達(dá)可使脂肪酸合成關(guān)聯(lián)基因的表達(dá)水平顯著提高,最終導(dǎo)致種子中脂肪酸和油脂的含量增加[8]。在擬南芥中超量表達(dá)WRI1基因,顯著提高了種子中油脂的含量(提高10 %~20 %),且不影響轉(zhuǎn)基因植株的生長(zhǎng)。實(shí)驗(yàn)表明,糖酵解和脂肪酸代謝進(jìn)程的基因直接受到WRI1調(diào)控[14]。此基因的效應(yīng)已在大田中得到證明,在玉米中超量表達(dá)WRI1,使玉米種子中油脂的含量增加了30 %[15]。研究發(fā)現(xiàn),油菜Testa16在植物發(fā)育中扮演著多重角色,參與油菜胚胎發(fā)育和脂肪酸合成[16]。將大豆Dof類(lèi)型的轉(zhuǎn)錄因子基因gmDof4和gmDof11轉(zhuǎn)化到擬南芥中,可以提高轉(zhuǎn)基因植株種子油脂含量[17]。
通過(guò)定量PCR研究證明AGL11基因在種子發(fā)育和油脂積累過(guò)程中表達(dá)強(qiáng)烈,表明該基因可能在這些發(fā)育過(guò)程中發(fā)揮著重要作用,后續(xù)實(shí)驗(yàn)通過(guò)獲得抑制AGL11基因表達(dá)轉(zhuǎn)基因油菜,并對(duì)轉(zhuǎn)基因油菜進(jìn)行胚分析和油脂含量測(cè)定,觀察在轉(zhuǎn)基因植株的種子和胚發(fā)育是否被強(qiáng)烈抑制,胚中脂肪酸含量是否顯著降低,通過(guò)上述研究,明確了AGL11基因在控制油菜種子中脂肪酸合成中的作用。
[1]Saha G, Park J I, Jung H J, et al. Genome-wide identification and characterization of MADS-box family genes related to organ development and stress resistance inBrassicarapa[J]. Bmc Genomics, 2015, 16(1): 1-21.
[2]Guo X, Chen G, Cui B, et al. Solanum lycopersicum agamous-like MADS-box protein AGL15-like gene, SlMBP11, confers salt stress tolerance[J]. Molecular Breeding, 2016, 36(9): 1-15.
[3]Ludovico D, Martin M K. MADS reloaded: evolution of the AGAMOUS subfamily genes[J].New Phytologist, 2014, 201: 717-732.
[4]Wei X C, Wang L, Yu J, et al. Genome-wide identification and analysis of the MADS-box gene family in sesame[J].Gene, 2015, 569(1): 66-76.
[5]Sun L M, Zhang J Z, Hu C G, et al. Characterization and Expression Analysis of PtAGL24, a SHORT VEGETATIVE PHASE/AGAMOUS-LIKE 24 (SVP/AGL24)-Type MADS-Box Gene from Trifoliate Orange (PoncirustrifoliataL. Raf.)[J]. Frontiers in Plant Science, 2016, 721 (7): 23-28.
[6]Ocarez N, Mejía N. Suppression of the D-class MADS-box AGL11 gene triggers seedlessness in fleshy fruits[J]. Plant Cell Reports, 2016, 35(1): 239-254.
[7]Wells C E, Vendramin E, Tarodo S J, et al. A genome-wide analysis of MADS-box genes in peach [Prunuspersica(L.) Batsch][J].Bmc Plant Biology, 2015, 15(1): 1-15.
[8]Tan H, Yang X, Zhang F, et al. Enhanced seed oil production in canola by conditional expression ofBrassicanapusLEAFY COTYLEDON1 and LEC1-LIKE in developing seeds[J].Plant Physiology, 2011,156: 1577-1588.
[9]An D H, Michung S. Overexpression of Arabidopsis WRI1 enhanced seed mass and storage oil content inCamelinasativa[J].Plant Biotechnology Reports, 2015,9(3): 137-148.
[10]Zheng Y, Ren N, Wang H, et al. Global identification of targets of the Arabidopsis MADS domain protein AGAMOUS-Like15[J]. Plant Cell, 2009,21(9): 2563-2577.
[11]Huang F,Chi Y,Gai J,et al. Identification of transcription factors predominantly expressed in soybean flowers and characterization of GmSEP1 encoding a SEPALLATA1-like protein[J].Gene,2009,438: 40-48.
[12]Adamczyk B J,F(xiàn)ernandez D E. MIKC* MADS domain heterodimers are required for pollen maturation and tube growth in Arabidopsis[J].PlantPhysiol,2009,149: 1713-1723.
[13]王漢中. 我國(guó)油菜產(chǎn)業(yè)發(fā)展的歷史回顧與展望[J]. 中國(guó)油料作物學(xué)報(bào),2010,32:300-302.
[14]Maeo K, Tokuda T, Ayame A, et al. An AP2-type transcription factor WRINKLED1 ofArabidopsisthalianabinds to AW-box sequence conserved among proximal upstream regions of genes involved in fatty acid synthesis[J].Plant Journal, 2009, 60: 476-487.
[15]Shen B, Allen W B, Zheng P, et al. Expression of ZMLEC1 and ZMWRI1 increases seed oil production in maize[J].Plant Physiology, 2010, 153: 980-987.
[16]Deng Wei, Hen G, Peng F, et al.TransparentTesta16 plays multiple roles in plant development and is involved in fatty acid synthesis and embryo development inBrassicanapus[J]. Plant Physiology, 2012, 160: 978-989.
[17]Wang H W, Zhang B, Hao Y J, et al. The soybean Dof-type transcription factor genes, GmDof4 and GmDof11, enhance lipid content in the seeds of transgenic Arabidopsis plants[J]. Plant Journal, 2007, 52: 716-729.
CloningandExpressionandTransformationofMADS-boxFamilyGeneAGL11inBrassicanapus
XIONG Shu1, LI Yan-jie2, ZHOU Da-xaing2,3*
(1.Department of Basic Medicine, Chongqing Three Gorges Medical College, Chongqing Wanzhou 404120, China; 2.College of Life Science and Engineering, Chongqing Three Gorges University, Chonqqing Wanzhou 404100, China; 3.College of Life Science, Chongqing University, Chongqing Key Lab of Genetic Function and Regulation,Chongqing Wanzhou 400030, China)
【Objective】This study aimed to provide a new clue for the exploration of the molecular regulation mechanism of oil synthesis inBrassicanapusseed. 【Method】The tissue expression pattern ofAGL11 gene by qPCR was analyzed, and its RNA interference vector was constructed. 【Result】Bioinformatics analysis showed that AGL11 protein ofBrassicanapushad a complete and typical MADS-box conserved domain, the overall shape of protein showed typical DNA nucleic acid binding structure, the AGL11 belonged to the MADS-box family transcription factor, and its genetic relationship was the closest with AGL11 protein ofArabidopsis. The qPCR study found thatAGL11 gene expression was the highest in the flowering stage and fruit clamp 5DAP (5 days seed after flowering), and its RNA interference vector was successfully constructed in the experiment. 【Conclusion】TheAGL11 gene was strongly expressed in seed development and oil accumulation, which may play an important role in the seed development and the oil synthesis. The successful construction of RNA interference vector would be favourable to further obtain the transgenicBrassicanapus, observe the embryo development and seed oil accumulation of transgenicBrassicanapusand thus determine theAGL11 gene to play an important role in the embryo development and oil synthesis.
Brassicanapus; MADS-box;AGL11; RNAi; Oil synthesis
1001-4829(2017)10-2174-05
10.16213/j.cnki.scjas.2017.10.003
2016-11-10
重慶市教委科學(xué)技術(shù)研究項(xiàng)目(KJ1502601);重慶市自然科學(xué)基金(cstc2016jcyjA2132);重慶市教育委員會(huì)項(xiàng)目(KJ1401001)
熊 書(shū)(1987-),女,講師,碩士,主要從事分子生物學(xué)研究;*為通訊作者:周大祥(1979-),男,副教授,博士,主要從事分子生物學(xué)研究,E-mail:dqzhou79@163.com。
Q945.79
A
(責(zé)任編輯 陳 虹)
猜你喜歡
食品工業(yè)(2023年12期)2023-12-12 12:56:16
糧油食品科技(2022年6期)2022-11-25 12:18:34
糧油食品科技(2022年6期)2022-11-25 12:18:30
基層中醫(yī)藥(2020年8期)2020-11-16 00:55:20
環(huán)境衛(wèi)生工程(2020年3期)2020-07-27 01:19:22
瘋狂英語(yǔ)·新讀寫(xiě)(2020年3期)2020-06-06 09:06:14
塑料助劑(2019年3期)2019-07-24 08:51:22
中國(guó)糧油學(xué)報(bào)(2019年4期)2019-07-12 09:06:38
食品與機(jī)械(2018年11期)2019-01-03 06:15:34
中國(guó)糧油學(xué)報(bào)(2018年12期)2018-01-26 13:32:04
