物種間關系網絡結構和形成機制的研究進展
2018-01-25 07:33:56李遠杰張玲
綠色科技 2018年14期
關鍵詞:機制
李遠杰 張玲
摘要:指出了生態學研究從網絡科學的發展中受益頗多,具體表現在:(1)網絡提供一系列工具描述和可視化生態系統;(2)為生態學中長期以來的核心問題(如物種共存機制)提供了一個新的視角。介紹了物種間相互作用網絡的研究方法和結構特點,綜述了網絡結構形成機制的假說及國內外研究進展,對網絡作為一個分析工具在生態及進化領域的應用前景進行了展望。
關鍵詞:關系網絡;網絡結構;機制
中圖分類號:S812
文獻標識碼:A
文章編號:1674-9944(2018)14-0010-04
1 引言
物種間相互作用普遍存在于自然界,沒有一個生物個體可以脫離與其他物種相互作用形成的網絡而單獨存在。物種間的相互作用對于種群的動態、協同進化、群落結構及生態系統功能維持具有重要作用。然而,在群落水平上研究物種間相互作用一直是一個棘手的問題,借助于社會學領域的網絡分析工具,來描述物種間相互作用的多樣性、評定生態系統穩定性和探討群落穩定性的維持機制,逐漸成為生態學的一個熱門領域。建立物種間關系網絡的具體方法是將每個物種或生物實體作為一個“節點( node)”,相互間作用作為一個“連接(link)”構建定性或定量關系矩陣(Interaction matrix),進而利用軟件如Pajek,Gephi和R軟件的Bipartite程序包等進行可視化,并使用社會網絡分析算法計算相應的網絡結構參數。
2 網絡的結構特征
物種間關系網絡通常表現出以下非隨機結構特征。
(1)不對稱性:指兩個相互作用的物種連接的不平衡性,包括連接數目的不對稱性與連接強度的不對稱性。網絡內的物種連接度(即每個物種的連接伙伴數目)符合冪律分布( Power-law distribution),大量物種有較少的連接伙伴,而僅有少量物種有較多的連接。連接強度指兩個物種相互作用的頻率,例如,對于傳粉網絡而言,植物與特定傳粉者的連接強度就是該傳粉者的訪問次數。通常僅有少量的物種具有較強的連接。網絡連接的不對稱性促進了群體中的物種共存,有利于維持物種多樣性。
(2)嵌套性:某一物種的一些連接伙伴是相對更為泛化的連接伙伴的一個子集。如,在傳粉網絡內,鱗翅目昆蟲訪問少量的植物,而蜜蜂訪問該網絡內的所有植物,與鱗翅目昆蟲相互作用的植物是與蜜蜂相互作用的植物種類的子集,即鱗翅日昆蟲的連接伙伴嵌套于蜜蜂的連接伙伴,呈現一定程度的嵌套性。這種嵌套結構有效地減少了種間競爭,增加共存物種的數量,從而保持群落內的生物多樣性,提高群落的穩定性。
(3)模塊性:網絡分成相對獨立、界限清晰的模塊的程度,物種在模塊內連接較為緊密,模塊間連接松散。模塊化可能通過限制物種間的相互作用及干擾在模塊間的擴散從而保持群落的穩定性。如Fortuna等人的研究表明,不同的蝙蝠棲息在不同群組的樹上而形成模塊.從而限制疾病在種群間的傳播。
網絡的結構通常與種間關系類型相關,互惠網絡包括植物一傳粉者網絡、植物一種子散布者網絡、植物一螞蟻網絡、寄主一共生者網絡等通常表現出較高的嵌套性,而拮抗性網絡,如食物網,嵌套性較弱、模塊性較強,尤其在植物植食者網絡表現明顯。
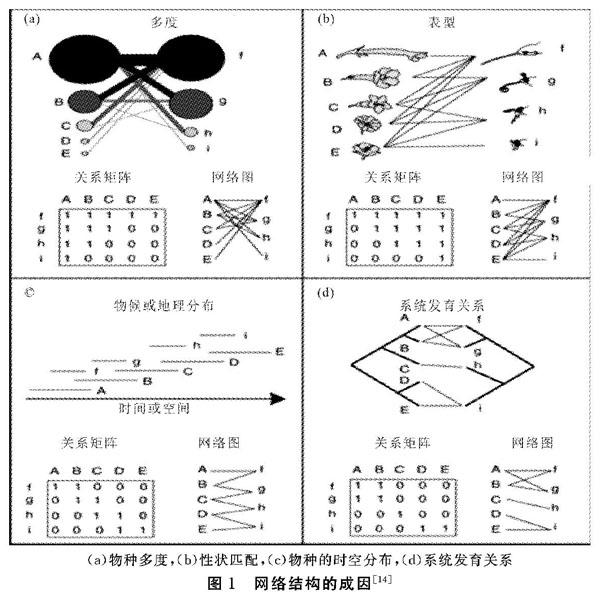
3 網絡結構的成因
隨著群落水平上的生態網絡研究的蓬勃發展,研究人員已經從描述網絡的結構和動態,而逐漸深入到對網絡結構形成的原因的探討。目前,對于網絡結構成因的解釋,有以下幾個假說。
3.1 中性假說
中性假說( Neutrality hypot,hesis)認為是物種問的相互作用是隨機的,因此,如圖1(a),多度較高的物種比稀有種有更高的幾率發生相互作用,也兇而擁有更多的連接伙伴。如,淡水魚一寄生蟲及哺乳動物一跳蚤兩個寄主一寄生網絡的高度的連接不對稱性可以由基于物種相對多度建立的模型預測,表明多度較高的寄主被較多種類的寄生感染,有更大的幾率與專性寄生建立關系。
3.2生物限制假說
生物限制假說(Biological constraint hypothesis)認為網絡內物種間的相互作用不是隨機的,而是受物種的表型、物候、空間分布和系統發育關系等性狀互補導致的限制性連接( Forbidden links)影響。如圖1(b),傳粉網絡中,長喙的傳粉者通常訪問花冠較長的植物;空間分布或物候不重疊的兩個物種無法發生相互作用(圖1c)。親緣關系相近的物種通常有相似的連接伙伴(圖1d)。實際研究中,也發現花型較大的植物更多地被大嘴的鳥類拜訪。
3.3多因素假說
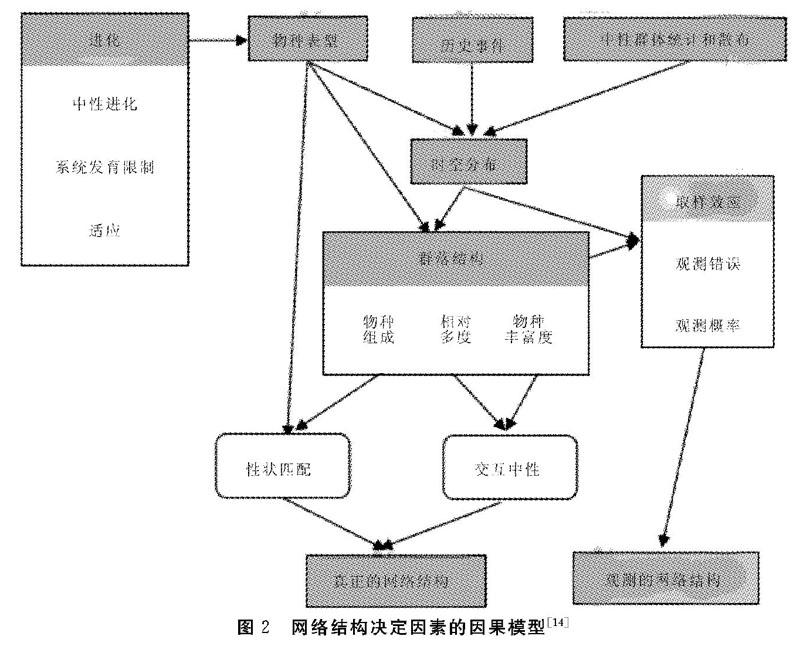
Vazquez等綜合了上述兩個假說,提出網絡結構是由多元等級因素作用的多個生態過程共同決定的(圖2),包括物種表型(Species phenotypes)、中性群體統計和散布(Netural demography and dispersal)和由此產:生的物種時空分布( Spatio - temporal distribution)、群落結構( Community structure)和取樣效應(Sampling effect)。我們所觀察到的網絡結構除去取樣偽影才是真正的網絡結構(True network structure),它是物種在交互中性(Interaction neutrality)與性狀匹配(Trait matching)的過程中相互作用累積的結果。Vazquez等提出上述網絡形成機制模型后,建立了對應的一套統計方法以便于其他研究者應用該理論框架。Sayago等使用了該理論模型和統計方法檢驗寄主性狀(胸徑、木質密度和樹皮粗糙度)、多度和物種空間分布重疊是否能預測定量的附生植物一寄主網絡。結果表明:物種多度、空間分布重疊及寄主大小(胸徑)可以極大預測物種間相互作用和幾個網絡特性參數,系統發育信號在網絡結構中不明顯。之后,該理論框架及統計方法迅速應用于更多的網絡研究中,探討不同類型的物種間相互作用網絡結構的影響因素。Vitoria等補充了該理論框架使之更加精確地闡明物種的進化歷史對網絡結構形成的貢獻。
4結語和展望
對于物種間關系網絡的研究,國內起步較晚,研究相對較少。但近些年,國內學者也開始逐漸涉及該領域。最早的一篇網絡研究來自于熱帶曲^雙版納地區,探討微生境和植物的性狀對榕屬植物一鳥類取食者關系網絡的影響,結果發現植物大小和樹冠體積顯著影響鳥類對榕屬植物的拜訪,網絡的嵌套度受森林類型和干擾程度的影響。傳粉網絡相較于其它類型的關系網絡研究得更多,其中,Fang和Huang和Zhao對處在中國生物多樣性熱點地區的傳粉網絡進行了深入的研究。對于網絡的理論研究,Yan和Zhang發現非單調性相互作用( Non - monotonous interaction)有利于物種共存和生態網絡穩定,并且其團隊通過對森林內植物的種子和鼠類關系的研究,發現種子物理和化學防御特征決定互惠與捕食關系網絡的形成。Gu利用已發表的種子散布網絡數據進行分析,發現泛化種由于擁有較多關系伙伴,降低了單個關系伙伴對其的影響力,因此減慢其進化的速率。近些年,不斷地有關于生態網絡的綜述報道。
從1970 - 2013年,物種間關系網絡的文章數量逐步增長,一些學者認為網絡分析方法代表生態學研究的新范式,因為網絡分析不僅是描述和量化復雜的生態群落的有力工具,也幫助解決群落構建、物種共存及營養級生態位等關鍵的生態問題;但有學者認為網絡分析存在一定缺陷如網絡參數的生態學含義的模糊性和不嚴謹性。盡管存在一定爭議,網絡分析方法作為一個探索工具已經越來越多地應用于生態學研究中。
隨著計算機技術及分析方法的發展,物種間關系網絡將從二元網絡( Binary network)向多層網絡(Multi-layer network)研究邁進,多層網絡可以同時描述群落內多個類型的物種間關系,如捕食、互惠和競爭等,將更加接近真實的群落結構。同時,物種間關系網絡結構的成因將被深入探討,對物種間相互作用和群落的時空變化動態有進一步理解。此外,網絡研究將從描述、理解向預測方向發展,尤其是預測人類活動引起的環境變化對物種間相互作用的影響。目前已有少量相關研究被報道,Staniczenko等預測了生境改變對物種間關系網絡的影響,對生物多樣性保護具有重要意義。
猜你喜歡
四川勞動保障(2021年9期)2022-01-18 05:11:08
文苑(2018年21期)2018-11-09 01:23:06
當代陜西(2018年9期)2018-08-29 01:21:00
當代陜西(2017年12期)2018-01-19 01:42:33
暨南學報(哲學社會科學版)(2016年9期)2017-01-15 13:52:00
中國衛生(2016年9期)2016-11-12 13:28:08
中國衛生(2015年9期)2015-11-10 03:11:12
醫學研究雜志(2015年12期)2015-06-10 06:57:46
中國衛生(2014年3期)2014-11-12 13:18:12
中國火炬(2014年4期)2014-07-24 14:22:19
