乳腺癌微環境成纖維細胞對乳腺癌細胞表達TIGAR和Bcl-2的影響①
2018-03-07 11:45:21陳少芬李文林石小玉
中國免疫學雜志 2018年2期
劉 賽 陳少芬 李文林 石小玉
(南昌大學醫學部基礎醫學院,南昌 330006)
乳腺癌是女性常見癌癥,是導致女性因癌癥死亡的最主要原因[1,2]。自2000年以來,女性乳腺癌的發病率持續增長[3,4],在中國,乳腺癌是45歲以下女性癌癥死亡的主要原因,其死亡率呈上升趨勢[5]。乳腺癌是多因素、多因子參與的疾病,其發病原因和機制較為復雜,需深入研究。TP53誘導型糖酵解和細胞凋亡調節劑(TP53-induced glycolysis and apoptosis regulator,TIGAR)是腫瘤代謝調節因子和凋亡抑制蛋白,TIGAR能降低細胞中的果糖-2,6-二磷酸水平,導致糖酵解的抑制和細胞內活性氧水平的降低,從而保護細胞免受ROS相關凋亡[6]。B-cell lymphoma-2(Bcl-2)是Bcl家族中最主要的凋亡抑制蛋白,參與腫瘤細胞的凋亡調節[7],靶向Bcl-2是腫瘤分子治療需探索的一條新途徑[8,9]。成纖維細胞是腫瘤間質微環境中主要細胞,對腫瘤的發生和發展起著重要的作用[10,11],本文研究了成纖維細胞與乳腺癌細胞共生長條件下,成纖維細胞對乳腺癌細胞TIGAR和Bcl-2的表達及對乳腺癌生長的影響,試圖為乳腺癌的發病機制研究提供新資料。
1 材料與方法
1.1主要實驗材料 人乳腺癌細胞株MDA-MB-231購于中科院上海細胞資源中心,人皮膚成纖維細胞株CCC-ESF-1(ESF)購自中國醫科院基礎醫學研究所細胞中心,雌性裸鼠購自上海斯萊克實驗動物有限責任公司(許可證號SCXK滬2012-0002),胎牛血清(Wisent生物技術中國有限公司),DMEM-H培養基(美國Life Tech-Gibco公司),RevertAid first strand cDNA 試劑盒(美國 Thermo Fisher 公司),qPCR試劑IQSYBRGreen Supermix(美國Bio-Rad 公司),RNA提取試劑盒(上海吉瑪生物公司),相關基因引物(上海吉瑪生物公司),鼠抗β-actin單克隆抗體 (北京中衫金橋生物技術有限公司),鼠抗人Bcl-2單克隆抗體(美國Bioworld公司),鼠抗人TIGAR單克隆抗體(美國Santa Cruz公司),羊抗鼠Ig(美國Santa Cruz公司),一步法聚合物 PV6002試劑盒(北京中衫金橋生物技術有限公司),孔徑0.4 μm Transwell小室(美國Corning公司),Caspase-3活性熒光檢測試劑盒和Annexin V凋亡試劑盒(Abcam中國分公司)等。
1.2實驗方法
1.2.1細胞培養及共培養 將MDA-MB-231細胞和ESF細胞放入含10%胎牛血清的DMEM-H 培養基中,置于37℃、5%CO2培養箱中培養,維持飽和濕度,待細胞生長至80%~90%匯合時,進行傳代培養。實驗分為三組:ESF單獨培養組、ESF+MDA-MB-231共培養組、MDA-MB-231單獨培養組。用6孔板和Transwell小室對ESF細胞和MDA-MB-231細胞進行體外共培養,小室接種細胞量為1.5×105/室,6孔板接種細胞量為2×105/孔。
1.2.2RT-qPCR實驗 用Trizol 法提取細胞總RNA,取2 μl 提取的RNA溶液,用分光光度計檢測,觀察A260/A280、A260/A230 比值及連續波長吸收峰,計算RNA 溶液濃度,判斷RNA 提取質量,2.3>A260/A280>2.0時,可用于后續逆轉錄實驗。以β-actin為內參,β-actin引物:上游引物5′-GAGAAAATCTGGCACACC-3′,下游引物5′-GGATAGCACAGCCTGGATAGCAA-3′;Bcl-2引物:上游引物5′-CACCCCCTCGTCCAAGAATG-3′,下游引物 5′-GCCACTCGTAGCCCCTCTGC-3′;TIGAR引物:上游引物5′-CAGCGGTATTCCAGGATTAG-3′,下游引物5′-ACCTTAGCGAGTTTCAGTCAG-3′。逆轉錄合成cDNA,-20℃保存cDNA。熒光定量PCR 擴增實驗,反應條件:50℃ 3 min,95℃ 3 min;95℃ 10 s,55℃20 s(Bcl-2:63.5℃ 20 s),72℃ 30 s,40個循環;溶解曲線溫度65~95℃。實驗結果由熒光定量PCR軟件Bio-Rad CFX Manager自動進行統計和計算分析。
1.2.3Western blot實驗 PBS分別洗滌細胞后用蛋白提取試劑盒提取總蛋白,BCA法測定蛋白濃度。10%聚丙烯酰胺凝膠電泳進行蛋白分離,轉移至PVDF膜,室溫下5%脫脂奶粉搖床封閉3 h,分別加入β-actin一抗、TIGAR一抗和Bcl-2一抗,4℃過夜孵育一抗,加入二抗并于搖床上室溫孵育1~2 h,ECL顯色曝光,觀察并拍照,用Image J軟件分析各條帶光密度 。
1.2.4Annexin V凋亡實驗 收集細胞,每一實驗樣本細胞數量為1×105,實驗組每管加5 μl Annexin V-Biotin ,對照組每管加5 μl 7-amino-actinomycin(7-AAD),用7-AAD以區別凋亡細胞與其他死細胞,室溫避光孵育 15 min;再在實驗組每管中加入Streptavidin-FITC,室溫避光孵育15 min,流式細胞儀檢測。
1.2.5Caspase-3活性熒光檢測實驗 提取細胞總蛋白,BCA法進行蛋白定量,實驗設空白對照孔和待測樣品孔,均加入含熒光底物AC-DEVD-AMC的Caspase-3反應緩沖液(45 μl /孔),空白每孔加5 μl細胞裂解液(含Cocktail),待測樣品每孔加10 μg(5 μl)細胞總蛋白,熒光分光光度計檢測AMC釋放量,分析熒光強度。
1.2.6人乳腺癌裸鼠移植瘤模型的建立 取6~7周齡雌性裸鼠,隨機實驗分組:①ESF+MDA-MB-231組;② MDA-MB-231組;③ESF組,分別向每組裸鼠右側胸壁第二乳腺注入100 μl細胞懸液(MDA-MB-231細胞濃度為2×106、ESF細胞濃度為0.5×106)。每隔1 d用游標卡尺測量瘤體的最長徑(a)和最短徑(b),以腫瘤長徑達5 mm為成瘤(成瘤約在細胞接種后的7 d左右)。 根據公式計算腫瘤體積并繪制腫瘤生長曲線,腫瘤的體積計算公式:V(mm3)=1/6πab2。在裸鼠成瘤30 d后將裸鼠頸椎脫臼處死,取出腫瘤組織放入4%福爾馬林溶液中固定,石蠟包埋切片,HE染色和免疫組織化學實驗。
1.2.7裸鼠移植瘤組織HE染色 石蠟切片經脫蠟至水后,用蘇木素-伊紅(HE)常規染色,在光學顯微鏡下觀察并拍照。
1.2.8裸鼠移植瘤組織免疫組織化學實驗 石蠟切片脫蠟至水,滴加3%H2O2,37℃孵育10 min,置入0.01 mol/L枸櫞酸緩沖液(pH6.0)中,入微波爐修復抗原,正常羊血清工作液封閉37℃正常血清工作液作用30 min,滴加一抗4℃孵育過夜,室溫平衡30 min,滴加相應二抗,37℃孵育30 min,DAB顯色,蘇木素復染,光學顯微鏡下觀察和拍照,Image-Pro Plus軟件分析。

2 結果
2.1與成纖維細胞共培養的乳腺癌細胞TIGAR和Bcl-2表達上調 RT-qPCR實驗檢測乳腺癌細胞MDA-MB-231的TIGAR和Bcl-2 mRNA表達,結果如圖1所示,與MDA-MB-231細胞單獨培養組比較,MDA-MB-231細胞+ESF細胞共培養組TIGAR mRNA表達顯著上調(P<0.05),是單獨培養組的1.53倍;共培養組MDA-MB-231細胞Bcl-2 mRNA表達顯著上調(P<0.05),是單獨培養組的1.44倍。Western blot實驗檢測MDA-MB-231細胞TIGAR和Bcl-2的蛋白表達,結果如圖2所示,與MDA-MB-231細胞單獨培養組比較,共培養組MDA-MB-231細胞TIGAR和Bcl-2蛋白表達顯著上調(P<0.05),分別是單獨培養組的1.49和1.81倍。上述結果說明乳腺癌微環境的成纖維細胞能上調乳腺癌細胞抗凋亡因子TIGAR和Bcl-2 mRNA和蛋白的表達。

圖1 RT-qPCR檢測體外培養的MDA-MB-231細胞抗凋亡因子TIGAR和Bcl-2 mRNA的表達Fig.1 mRNA expression of TIGAR and Bcl-2 in MDA-MB-231 cells detected by RT-qPCR in vitro Note: *.P<0.05.
2.2與成纖維細胞共培養的乳腺癌細胞凋亡下降在與成纖維細胞ESF共培養條件下,乳腺癌細胞MDA-MB-231的抗凋亡因子TIGAR和Bcl-2表達上調。MDA-MB-231細胞Caspase-3凋亡蛋白酶活性實驗結果顯示(圖3A),MDA-MB-231細胞+ESF細胞共培養組MDA-MB-231細胞Caspase-3熒光強度顯著低于MDA-MB-231細胞單獨培養組(P<0.05),表明在ESF細胞共培養的微環境下乳腺癌細胞Caspase-3活性降低,這將有利于乳腺癌細胞生長。Annexin V細胞凋亡實驗結果顯示(圖3B),與單獨培養組比較,共培養組早期凋亡的MDA-MB-231細胞下降了2.12倍(4.85/2.29),說明乳腺癌微環境的成纖維細胞通過上調乳腺癌細胞抗凋亡因子TIGAR和Bcl-2的表達從而抑制乳腺癌細胞的凋亡。

圖2 Western blot檢測體外培養的MDA-MB-231細胞抗凋亡因子TIGAR和Bcl-2蛋白的表達Fig.2 Protein expression of TIGAR and Bcl-2 in MDA-MB-231 cells detected by Western blot in vitroNote: A.The electrophoretic bands of TIGAR and Bcl-2 proteins in each group detected by Western blot;B.The optical density of electrophoretic bands of TIGAR and Bcl-2 proteins in each group analyzed by Image J; *.P<0.05.

圖3 Caspase-3活性和Annexin V檢測MDA-MB-231 細胞凋亡Fig.3 MDA-MB-231 cell apoptosis detected by Caspase-3 activity and Annexin VNote: A.Caspase-3 activity in MDA-MB-231 cells detedted by fluorescence;B.MDA-MB-231 cell apoptosis detected by Annexin V and flow cytometry; *.P<0.05.
2.3裸鼠移植瘤微環境的成纖維細胞促進腫瘤生長 將ESF細胞和MDA-MB-231細胞接種于6~7周的雌性裸鼠右側胸壁第二乳腺,第5天,可在ESF+MDA-MB-231組和MDA-MB-231組的裸鼠乳腺觸及腫瘤小結節,7~10 d腫瘤最長徑達5 mm,成瘤率為100%,人乳腺癌裸鼠荷瘤模型構建成功(圖4A)。第30天,將荷瘤裸鼠頸椎脫臼處死,取瘤組織塊,制成石蠟切片,HE染色。400倍顯微鏡下觀察,可見正常乳腺小葉結構消失,細胞聚集成巢,排列紊亂,核大,出現組織結構的異型性,HE染色證實為癌組織(圖4B)。在接種后的第1天至第16天ESF+MDA-MB-231組與MDA-MB-231組的腫瘤體積差異無統計學意義(P>0.05),自第18天起ESF+MDA-MB-231組的腫瘤體積較MDA-MB-231組大(圖5和表1),表明成纖維細胞與乳腺癌細胞共同移植入雌性裸鼠乳腺內所形成的腫瘤生長快于乳腺癌細胞單獨移植,裸鼠移植瘤微環境成纖維細胞具有促進腫瘤生長的作用。

圖4 人乳腺癌荷瘤裸鼠移植瘤Fig.4 Human breast cancer-transplanted tumor in nude miceNote: A.The transplanted tumor in nude mice;B.The transplanted tumor tissues stained by HE (×400).

圖5 人乳腺癌裸鼠移植瘤生長曲線Fig.5 Growth curve of human breast cancer-transplanted tumor in nude mice


圖6人乳腺癌荷瘤裸鼠移植瘤組織TIGAR的表達
Fig.6ExpressionofTIGARofhumanbreastcancer-transplantedtumortissuesinnudemice
Note: A.The immunohistochemical optical micrograph (×400) of TIGAR in the transplanted tumor tissues of ESF+MDA-MB-231 groups;B.The immunohistochemical optical micrograph (×400) of TIGAR in the transplanted tumor tissues of MDA-MB-231 groups;C.The optical density of immunohistochemical staining of TIGAR in the transplanted tumor tissues of ESF+MDA-MB-231 groups and MDA-MB-231 groups analyzed by Image-pro-plus 5.1 software; *.P<0.05.

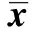

Group10d12d14d16d18d20dESF+MDA?MB?231(n=16)65 11±10 0979 86±21 65107 51±37 01128 55±41 96159 02±53 891)188 32±68 621)MDA?MB?231(n=16)64 96±11 1280 07±22 01105 67±38 32125 01±38 32138 22±47 04156 66±58 23Group22d24d26d28d30dESF+MDA?MB?231(n=16)229 98±87 781)268 83±96 541)312.17±105 691)359 21±116 721)432 09±127 861)MDA?MB?231(n=16)176 47±61 36211 56±73 35243 85±89 06285 36±99 01325 77±106 54
Note:Compared with MDA-MB-231 group,1)P<0.01.


圖7人乳腺癌荷瘤裸鼠移植瘤組織Bcl-2的表達
Fig.7ExpressionofBcl-2ofhumanbreastcancer-transplantedtumortissuesinnudemice
Note: A.The immunohistochemical optical micrographs (×400) of Bcl-2 in the transplanted tumor tissues of ESF+MDA-MB-231 groups;B.The immunohistochemical optical micrographs (×400) of Bcl-2 in the transplanted tumor tissues of MDA-MB-231 groups;C.The optical density of immunohistochemical staining of Bcl-2 in the transplanted tumor tissues of ESF+MDA-MB-231 groups and MDA-MB-231 groups analyzed by Image-pro-plus 5.1 software; *.P<0.05.
2.4TIGAR和Bcl-2在人乳腺癌裸鼠移植瘤組織的表達 免疫組化實驗檢測TIGAR和Bcl-2在裸鼠移植瘤組織中的表達,TIGAR和Bcl-2的陽性結果為細胞質出現棕黃色顆粒 (圖6A、B和圖7A、B)。每組隨機選取5張切片,每張切片選擇5個不同視野在×400倍光學顯微鏡下進行觀察拍照,采用Image-pro-plus 5.1軟件測量兩組移植瘤組織切片TIGAR和Bcl-2的光密度,計算平均光密度,結果顯示(圖6C和圖7C),與MDA-MB-231組比較,ESF+MDA-MB-231組移植瘤組織TIGAR和Bcl-2的平均光密度值大(P<0.05),ESF+MDA-MB-231組荷瘤裸鼠移植瘤組織中TIGAR和Bcl-2的表達顯著高于MDA-MB-231組移植瘤組織。表明乳腺癌細胞與成纖維細胞共生長的微環境可促進移植瘤組織TIGAR和Bcl-2的表達。
3 討論
癌癥的發生和發展不僅與腫瘤本身惡性增殖和侵襲等有關,還與它所處的微環境即腫瘤微環境密切相關[12]。腫瘤微環境包括細胞外基質、成纖維細胞(癌相關成纖維細胞,CAFs)、血管、免疫細胞和因子等[13]。其中成纖維細胞占腫瘤微環境的大部分,它通過分泌細胞因子和對細胞外基質的修飾等方式作用腫瘤細胞,對腫瘤的生長起著重要作用[14,15]。為了模擬乳腺腫瘤微環境,我們采用Transwell 方法,將人乳腺癌細胞MDA-MB-231與人成纖維細胞ESF在同一培養液內培養,但細胞不互混,建立了人乳腺癌細胞與人成纖維細胞體外共培養模型;將人乳腺癌細胞MDA-MB-231與人成纖維細胞ESF共同植入裸鼠乳腺內,建立了人乳腺癌細胞與人成纖維細胞體內共生長模型。通過體內體外模擬的乳腺腫瘤微環境模型,研究成纖維細胞對乳腺癌細胞TIGAR和Bcl-2表達的影響及對乳腺癌生長的影響。
在惡性腫瘤發生和發展的過程中,抑制癌細胞凋亡起著重要作用。Bcl家族在細胞凋亡信號轉導途徑中發揮重要作用,Bcl家族的表達和調控是影響細胞凋亡的關鍵因素之一[16,17]。Bcl-2是Bcl家族中最主要的凋亡抑制蛋白,與多數腫瘤的發生、發展有關,通過抑制細胞凋亡和抵抗其他形式的細胞死亡導致細胞數增多,對腫瘤的生長具有促進作用[18]。TIGAR是一凋亡抑制因子,可導致細胞內糖酵解下降和糖有氧氧化下降,減少ROS(活性氧族)的產生,保護細胞免于ROS相關的凋亡,在氧化應激下TIGAR可降低自噬水平促進細胞存活,在能量限制下,可以通過增加呼吸率提升能量供給,減少饑餓誘導的細胞死亡[19,20]。
本研究體外實驗結果顯示,在共培養條件下,成纖維細胞能上調乳腺癌細胞TIGAR和Bcl-2的表達,RT-qPCR和Western blot的實驗結果相互印證。隨后研究了TIGAR和Bcl-2表達上調對乳腺癌細胞凋亡的影響,采用Annexin V-Biotin流式細胞術和Caspase-3活性熒光檢測乳腺癌細胞凋亡,實驗結果顯示,成纖維細胞+乳腺癌細胞共培養組的乳腺癌細胞凋亡低于乳腺癌細胞單獨培養組,表明腫瘤微環境的成纖維細胞通過上調乳腺癌細胞TIGAR和Bcl-2的表達從而抑制乳腺癌細胞凋亡促進腫瘤生長。我們的體內實驗結果顯示,成纖維細胞+乳腺癌細胞共生長組的荷瘤裸鼠乳腺癌組織TIGAR和Bcl-2的表達上調,成纖維細胞與乳腺癌細胞共同移植入雌性裸鼠乳腺內所形成的腫瘤體積大于乳腺癌細胞單獨移植,表明基質成纖維細胞可促進乳腺癌細胞生長,且涉及乳腺癌微環境成纖維細胞上調乳腺癌細胞的抗凋亡因子TIGAR和Bcl-2的表達,TIGAR和Bcl-2的高表達致使荷瘤裸鼠乳腺癌生長加快。
綜上所述,乳腺癌微環境成纖維細胞可上調乳腺癌細胞TIGAR和Bcl-2的表達,促進TIGAR和Bcl-2的抗凋亡作用,從而加快乳腺癌的生長。乳腺癌微環境成纖維細胞上調乳腺癌細胞TIGAR和Bcl-2表達的信號轉導通路有待進一步研究。
[1] Ferlay J,Soerjomataram I,Dikshit R,etal.Cancer incidence and mortality worldwide:sources,methods and major patterns in globocan 2012[J].Int J Cancer,2015,136(5):E359-E386.
[2] Torre LA,Islami F,Siegel RL,etal.Global cancer in women:burden and trends[J].Cancer Epidemiol Biomarkers Prev,2017,26(4):444-457.
[3] Fitzmaurice C,Dicker D,Pain A,etal.The global burden of cancer 2013[J].JAMA Oncol,2015,1(4):505-527.
[4] Dubey AK,Gupta U,Jain S.Breast cancer statistics and prediction methodology:a systematic review and analysis[J].Asian Pac J Cancer Prev,2015,16(10):4237-4245.
[5] Chen W,Zheng R,Baade PD,etal.Cancer statistics in China,2015[J].CA Cancer J Clin,2016,66(2):115-132.
[6] Bensaad K,Tsuruta A,Selak MA,etal.TIGAR,a p53-inducible regulator of glycolysis and apoptosis[J].Cell,2006,126(1):107-120.
[7] Akl H,Vervloessem T,Kiviluoto S,etal.A dual role for the anti-apoptotic Bcl-2 protein in cancer:mitochondria versus endoplasmic reticulum [J].Biochim Biophys Acta,2014,1843(10):2240-2252.
[8] Schenk RL,Strasser A,Dewson G.Bcl-2:Long and winding path from discovery to therapeutic target [J].Biochem Biophys Res Commun,2017,482(3):459-469.
[9] Garner TP,Lopez A,Reyna DE,etal.Progress in targeting the Bcl-2 family of proteins [J].Curr Opin Chem Biol,2017,39:133-142.
[10] Alphonso A,Alahari SK.Stromal Cells and integrins:confor ming to the needs of the tumor microenvironment[J].Neoplasia,2009,11(12):1264-1271.
[11] Xing F,Saidou J,Watabe K.Cancer associated fibroblasts (CAFs) in tumor microenvironment[J].Front Biosci,2010,1(15):166-179.
[12] Allen M,Louise Jones J.Jekyll and Hyde:the role of the microenvironment on the progression of cancer[J].J Pathol,2011,223(2):162-176.
[13] Casey SC,Amedei A,Aquilano K,etal.Cancer prevention and therapy through the modulation of the tumor microenvironment[J].Se min Cancer Biol,2015,35 Suppl:S199-223.
[14] Folgueira MA,Maistro S,Katayama ML,etal.Markers of breast cancer stromal fibroblasts in the primary tumour site associated with lymph node metastasis:a systematic review including our case series[J].Biosci Rep,2013,33(6):921-929.
[15] Soysal SD,Tzankov A,Muenst SE.Role of the tumor microenvironment in breast cancer[J].Pathobiology,2015,82(3-4):142-152.
[16] Guo B,Zhai D,Cabezas E,etal.Humanin peptide suppresses apoptosis by interfering with Bax activation[J].Nature,2003,423(6938):456-461.
[17] 劉 芳,趙世敏,張 威,等.沙棘多糖對急性肝損傷小鼠氧化應激的抑制作用及其對 BCL-2/Bax和PPAR-γ的調控[J].中國免疫學雜志,2016,32(3):358-361.
Liu F,Zhao SM,Zhang W,etal.Inhibitory effects of seabuckthorn polysaccharide on oxidative stress in mice with actue liver injury and modulatory effect on Bcl-2/Bax and PPAR-γ expression[J].Chin J Immunol,2016,32(3):358-361.
[18] Jagani H,Kasinathan N,Meka SR,etal.Antiapoptotic Bcl-2 protein as a potential target for cancer therapy:a mini review[J].Artif Cells Nanomed Biotechnol,2016,44(5):1212-1221.
[19] Wanka C,Steinbach JP,Rieger J.Tp53-induced glycolysis and apoptosis regulator (TIGAR) protects glioma cells from starvation-induced cell death by up-regulating respiration and improving cellular redox homeostasis[J].J Biol Chem,2012,287(40):33436-33446.
[20] Cheung EC,Ludwig RL,Vousden KH.Mitochondrial localization of TIGAR under hypoxia stimulates HK2 and lowers ROS and cell death [J].Proc Natl Acad Sci U S A,2012,109(50):20491-20496.
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中老年保健(2022年6期)2022-08-19 01:41:48
小讀者(2021年2期)2021-03-29 05:03:48
華人時刊(2019年13期)2019-11-17 14:59:54
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
中國生殖健康(2019年2期)2019-08-23 08:11:42
中國生殖健康(2019年6期)2019-01-06 09:20:12
文苑(2018年22期)2018-11-19 02:54:14
祝您健康(2018年5期)2018-05-16 17:10:16
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
