shRNA沉默趨化素對小鼠主動脈平滑肌細胞增殖能力的影響及可能的機制*
2018-04-10 02:47:20牛美芝楊輝劉福壘
中國現代醫學雜志 2018年10期
關鍵詞:小鼠
牛美芝,楊輝,劉福壘
(1.山東省肥城礦業中心醫院 心內科,山東 肥城 271608;2.山東省泰安市中心醫院,山東 泰安 271000)
冠狀動脈粥樣硬化性心臟病(冠心病)作為嚴重威脅人類健康的常見病,近年來發病率呈逐年上升趨勢,冠狀動脈粥樣硬化是其重要的病理基礎[1],研究表明[2],冠狀動脈粥樣硬化作為血管壁的一種慢性炎癥疾病,是炎癥反應、血管內皮細胞功能異常、血管平滑肌細胞異常增殖及遷移等共同作用的結果。有研究指出[3],血管平滑肌細胞增殖及遷移異常在動脈粥樣硬化發生及進展中發揮基礎性作用。趨化素(Chemerin)作為新發現的一種趨化因子和脂肪因子,與肥胖、糖脂代謝異常等密切相關[4],研究發現[5],Chemerin在人冠狀動脈周圍脂肪組織及冠狀動脈病變部位的血管平滑肌細胞中高表達。亦有研究指出[6],Chemerin與其受體人趨化因子樣受體1(chemokine receptor-like 1,CMKLR1)結合通過活化下游相應信號通路而參與了冠心病發生及進展過程。本研究通過轉染Chemerin基因RNA干擾慢病毒載體,觀察其對小鼠主動脈平滑肌細胞(aortic smooth muscle cell,ASMC)增殖能力的影響,并探討可能的機制。
1 材料與方法
1.1 主要試劑與設備
SPF級ICR小鼠購自河南省實驗動物中心[許可證號SCXK(豫)2010-0002],雌雄不限,8~10周齡,飼養于標準環境下。DMEM培養基、胎牛血清、青鏈霉素混合液均購自美國Gibco公司,0.25%胰蛋白酶、Trizol總RNA提取試劑盒和Lipofectamine 2000轉染試劑盒均購自美國Invitrogen公司,慢病毒表達載體試劑盒購自日本TaKaRa公司,293T包裝細胞購自廣州賽業生物公司,血小板源性生長因子-BB(platelet-derived growth factor,PDGF-BB)購自上海古朵生物科技有限公司,逆轉錄試劑盒和聚合酶鏈反應試劑盒均購自大連寶生物公司,Chemerin及內參引物均由生工生物工程(上海)股份有限公司設計合成,兔抗小鼠α平滑肌肌動蛋白抗體購自北京中衫金橋生物技術有限公司,小鼠抗大鼠Chemerin單克隆抗體購自美國Enzo Life Sciences公司,兔抗鼠細胞外調節蛋白激酶1/2(ERK1/2)單克隆抗體購自美國Cell Signalling公司,小鼠抗大鼠p-ERK1/2多克隆抗體、鼠抗c-Jun氨基末端激酶(JNK)單克隆抗體購自美國Biosource公司,鼠抗p-JNK單克隆抗體購自上海瑞齊生物科技有限公司,兔抗山羊二抗購自武漢博士德生物技術有限公司,5-溴脫氧尿嘧啶核苷(Brdu)抗體購自美國Sigma公司,凝膠電泳分析系統購自美國Bio-Rad公司,實時熒光定量聚合酶鏈反應(quantitative real-time polymerase chain reaction,qRTPCR)儀購自美國ABI公司。
1.2 方法
1.2.1Chemerin基因RNA干擾慢病毒載體構建及鑒定根據小鼠Chemerin基因mRNA序列為干擾靶點,設計其靶向干擾的短發夾RNA序列(shRNA),正向:5'-GATCCGCTTTGTGAGGTTGGAATTTAATTCAAG AGATTAAATTCCAACCTCACAAAGTTTTTTACGCGT G-3',反向:5'-AATTCACGCGTAAAAAACTTTGTGAG GTTGGAATTTAATCTCTTGAATTAAATTCCAACCT CACAAAGCG-3'。根據試劑盒說明書,構建利用293T細胞對慢病毒進行包裝,采用梯度稀釋后,利用熒光計數對病毒滴度進行檢測。
1.2.2小鼠ASMC培養、鑒定及分組利用頸椎脫臼法將小鼠處死,無菌操作下將胸主動脈取出,用眼科剪剪成小塊狀,胰酶預消化后,利用組織塊貼壁法對ASMC進行原代細胞培養。細胞貼壁生長后進行傳代培養,并采用免疫熒光法對ASMC進行鑒定。取傳代培養細胞,分成4組:Sham組、PDGF組、對照序列組和Chemerin沉默組,其中,Chemerin沉默組和對照序列組分別用攜帶Chemerin干擾基因和對照基因序列的慢病毒進行感染,分別向PDGF組、對照序列組和Chemerin沉默組細胞中加入終濃度為20 ng/ml的PDGF-BB,Sham組則加入等量的PBS。
1.2.3qRT-PCR檢測各組細胞中Chemerin基因表達取各組傳代培養48 h細胞,加入細胞裂解液進行裂解,用Trizol總RNA提取試劑盒對總RNA進行提取,用紫外分光光度計對總RNA純度進行檢測,取A260/A280≥1.80作為合格樣品。用逆轉錄試劑盒進行逆轉錄,獲得模板單鏈cDNA,以cDNA為模板進行PCR。引物序列:Chemerin引物,正向:5'-TACAGGTG GCTCTGGAGGAGTTC-3',反向:5'-CTTCTCCCGTTTG GTTTGATTG-3';β-actin 引物,正向:5'-AGCCATGTA CGTAGCCATCC-3',反向:5'-CTCTCAGCTGTGGTGGT GAA-3'。PCR 反應條件:92℃ 1 min,92℃ 30 s,58℃30 s,73℃ 30 s,連續進行 40次循環。用2-△△Ct法獲得各組細胞中Chemerin基因相對表達量。
1.2.4細胞計數實驗取各組細胞,胰酶消化后離心,接種于96孔板中,調整細胞密度為1×104個/孔,每組設置6個反應復孔,4~6 h后改用100μl完全培養基進行培養。在加入PDGF-BB后培養48 h時,去除培養基,用含EDTA的胰酶30μl消化后,加入培養液70μl,充分吹打,使細胞充分混合均勻,取細胞懸液10μl,于顯微鏡下利用計數板對各組細胞進行計數。
1.2.5BrdU摻入法測定各組細胞增殖能力取各組傳代培養48 h細胞,接種于96孔板,將10μmol/L的BrdU溶液加入各孔,于恒溫培養箱中培養10 h,將200μl的FixDenant溶液加入,室溫下孵育25 min,于2 000 r/min離心10 min,棄上清,將100μl的抗BrdU-POD工作液加入,室溫下孵育100 min,于2 000 r/min離心10 min,棄上清,緩沖液反復沖洗3次,棄上清,將100μl的底物溶液加入,室溫下孵育25 min,將25μl的H2SO4溶液加入,于400 r/min離心2 min,于20 min內利用酶標儀取450 nm波長對吸光度(A)值進行檢測。
1.2.6Western blot檢測各組細胞中Chemerin、ERK1/2、p-ERK1/2、JNK和p-JNK蛋白表達取各組傳代培養48 h細胞,加入細胞裂解液進行裂解,用總蛋白提取試劑盒對總蛋白進行提取,用BCA蛋白檢測試劑盒對蛋白濃度進行檢測。取30μg蛋白樣品,進行SDS-PAGE電泳,電轉移至PVDF膜上,用5%脫脂奶粉封閉120 min,分別將小鼠抗大鼠Chemerin單克隆抗體、兔抗鼠細胞外調節蛋白激酶1/2(ERK1/2)單克隆抗體、小鼠抗大鼠p-ERK1/2多克隆抗體、鼠抗JNK單克隆抗體和鼠抗p-JNK單克隆抗體(稀釋比例:1∶800、1∶1 200、1∶1 000、1∶500和1∶800)加入,4℃下過夜孵育,用TBST洗滌3次,將二抗加入,室溫下孵育60 min,TBST洗滌3次,用ECL發光試劑盒避光反應30 min,利用Image J圖像分析軟件對獲得的條帶進行分析,獲得各組細胞中 Chemerin、ERK1/2、p-ERK1/2、JNK 和 p-JNK蛋白相對表達量。
1.3 統計學方法
利用SPSS21.0軟件對數據進行分析,計量資料采用均數±標準差(±s)表示,多組間比較采用單因素方差分析,組間兩兩比較采用LSD-t檢驗,P<0.05為差異有統計學意義。
2 結果
2.1 小鼠ASMC培養及慢病毒感染檢測結果
顯微鏡下觀察,小鼠ASMC呈不規則形、三角形或長梭形,體積較小,見圖1A;沉默組和對照序列組小鼠ASMC分別轉染Chemerin基因感染序列和陰性對照序列后,培養48 h,熒光顯像觀察可見綠色熒光達90%以上,見圖1B,提示ASMC細胞慢病毒感染成功。
2.2 各組細胞中Chemerin基因表達比較
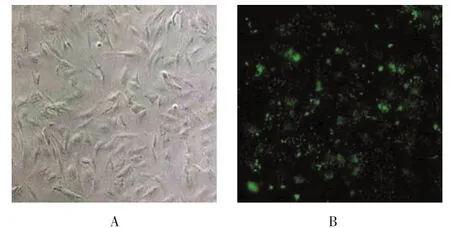
圖1 小鼠ASMC培養及慢病毒感染檢測結果
qRT-PCR檢測各組細胞中Chemerin表達水平,經單因素方差分析,差異有統計學意義(F=172.943,P=0.000),各組Chemerin表達水平有差異;進一步兩兩比較顯示,與Sham組相比,對照序列組和PDGF組Chemerin mRNA相對表達量均增高(t=31.173和29.351,均P=0.000),Chemerin沉默組Chemerin mRNA相對表達量降低(t=16.817,P=0.000)。與對照序列組和PDGF組相比,Chemerin沉默組Chemerin mRNA相對表達量均降低(t=45.183和42.953,均P=0.000),見圖 2。
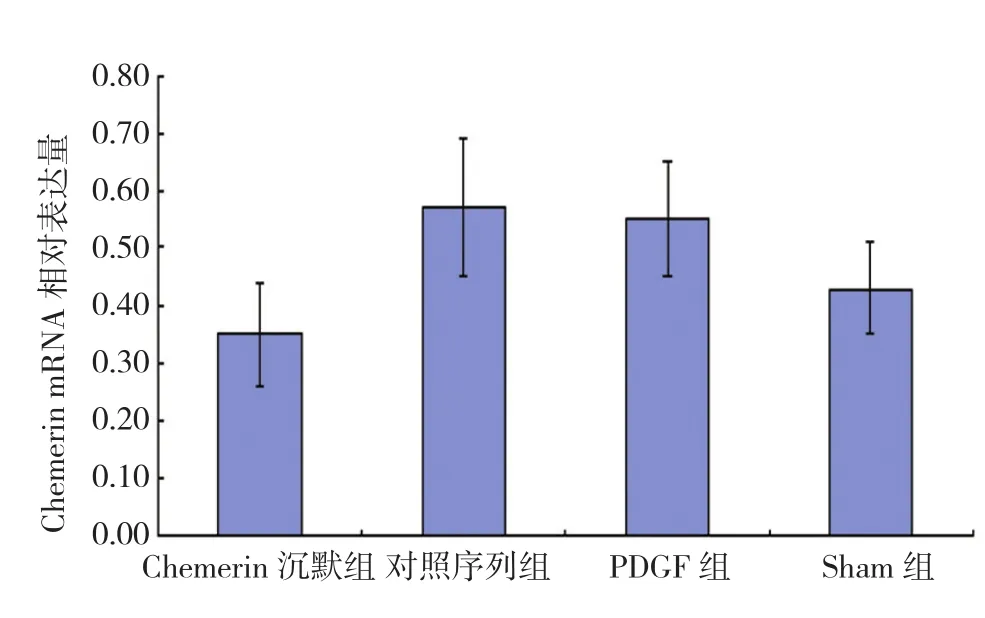
圖2 各組細胞中Chemerin基因表達
2.3 各組細胞增殖能力比較
細胞計數實驗顯示,Chemerin沉默組、對照序列組、PDGF組和Sham組細胞數分別為(34.27±3.08)×103個 /cm2、(58.62±3.71)×103個 /cm2、(61.35±4.16)×103個 /cm2和(46.14±2.95)×103個 /cm2,經單因素方差分析,差異有統計學意義(F=208.318,P=0.000),各組細胞數有差別;進一步兩兩比較顯示,與Sham組相比,對照序列組和PDGF組細胞數均增加(t=46.826和48.617,均P=0.000),而Chemerin沉默組細胞數降低(t=19.381,P=0.000);與對照序列組和PDGF組相比,Chemerin沉默組細胞數均降低(t=61.291和 64.064,均P=0.000)。
BrdU摻入法結果顯示,Chemerin沉默組、對照序列組、PDGF組和Sham組吸光度A值分別為(1.26±0.07)、(1.58±0.09)、(1.57±0.10)和(1.43±0.08),經單因素方差分析,差異有統計學意義(F=49.164,P=0.000),各組細胞吸光度A值有差別;進一步兩兩比較顯示,與Sham組相比,對照序列組和PDGF組吸光度A值均增加(t=10.517和10.483,均P=0.000),而Chemerin沉默組吸光度A值降低(t=13.067,P=0.000);與對照序列組和PDGF組相比,Chemerin沉默組吸光度A值均降低(t=16.862和16.798,均P=0.000)。
2.4 各組細胞中 Chemerin、ERK1/2、p-ERK1/2、JNK和p-JNK蛋白表達比較
Western blot檢測各組細胞中Chemerin、ERK1/2、p-ERK1/2、JNK和p-JNK蛋白表達水平,經單因素方差分析,差異有統計學意義(F=39.167、48.375、31.272、56.759和 40.391,均P=0.000),各組細胞中 Chemerin、ERK1/2、p-ERK1/2、JNK 和 p-JNK蛋白表達水平有差別;進一步兩兩比較顯示,與Sham組相比,對照序列組和PDGF組Chemerin、p-ERK1/2、p-JNK灰度均增高(t=26.842、27.167、20.572、19.883、11.057 和 10.837,均P=0.000),而ERK1/2、JNK灰度均降低(t=29.382、29.168、32.672和32.405,均P=0.000);與Sham組相比,Chemerin沉默組Chemerin、ERK1/2、p-ERK1/2、JNK、p-JNK灰度均降低(t=13.286、12.195、7.235、11.836和10.672,P=0.000、0.000、0.001、0.000 和 0.000);與對照序列組和PDGF組相比,Chemerin沉默組Chemerin、p-ERK1/2、p-JNK灰度均降低(t=37.268、38.173、11.971、11.684、31.824和 32.174,均P=0.000),而ERK1/2、JNK 灰 度 均 增 加(t=13.082、13.574、13.582和14.015,均P=0.000),見圖3和附表。
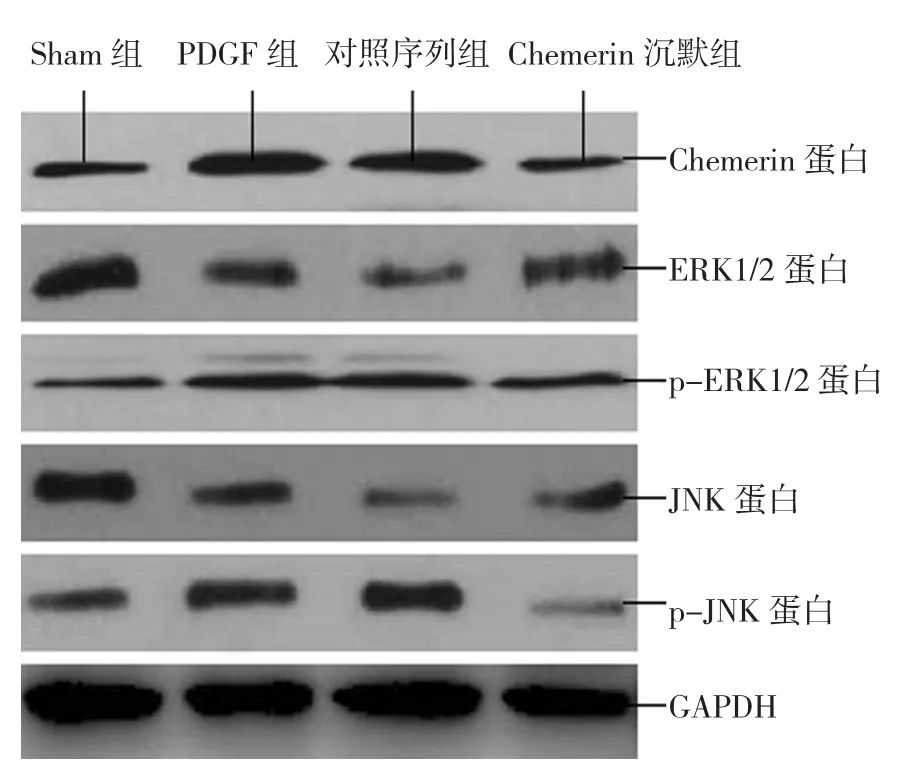
圖3 各組細胞中Chemerin、ERK1/2、p-ERK1/2、JNK和p-JNK蛋白表達
附表 各組細胞中Chemerin、ERK1/2、p-ERK1/2、JNK和p-JNK蛋白表達比較 (±s)
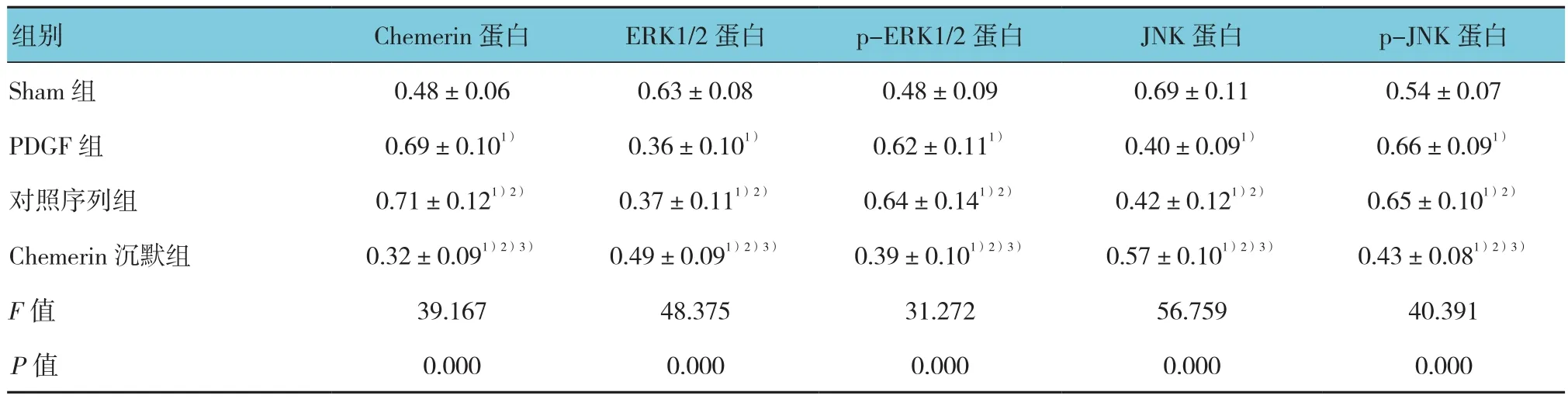
附表 各組細胞中Chemerin、ERK1/2、p-ERK1/2、JNK和p-JNK蛋白表達比較 (±s)
注:1)與Sham組相比,P <0.05;2)與PDGF組相比,P <0.05;3)與對照序列組相比,P <0.05
組別 Chemerin蛋白 ERK1/2蛋白 p-ERK1/2蛋白 JNK蛋白 p-JNK蛋白Sham 組 0.48±0.06 0.63±0.08 0.48±0.09 0.69±0.11 0.54±0.07 PDGF 組 0.69±0.101) 0.36±0.101) 0.62±0.111) 0.40±0.091) 0.66±0.091)對照序列組 0.71±0.121)2) 0.37±0.111)2) 0.64±0.141)2) 0.42±0.121)2) 0.65±0.101)2)Chemerin 沉默組 0.32±0.091)2)3) 0.49±0.091)2)3) 0.39±0.101)2)3) 0.57±0.101)2)3) 0.43±0.081)2)3)F值 39.167 48.375 31.272 56.759 40.391 P值 0.000 0.000 0.000 0.000 0.000
3 討論
冠狀動脈粥樣硬化作為冠心病和冠心病介入治療術后再狹窄發生的共同病理基礎,對患者生命健康造成嚴重影響[7]。該病發生機制較為復雜,目前尚未完全清楚。Chemerin作為新近發現的脂肪因子,廣泛存在于機體組織中,與肥胖相關疾病發生及糖脂代謝失衡密切相關[8]。有研究指出[9],冠心病患者血漿Chemerin水平升高,且與病情嚴重程度呈正相關。研究表明[10],Chemerin與存在于人血管內皮細胞上的特異性CMKLR1結合可誘導血管內皮細胞增殖及血管新生。而血管平滑肌細胞異常增殖在冠狀動脈粥樣硬化發生及血管重構中發揮關鍵性作用[11]。亦有研究指出[12],動脈粥樣斑塊增殖中Chemerin呈高表達。這些研究表明Chemerin可能參與了動脈粥樣硬化發生及進展過程,但具體機制尚不明確。
本研究利用RNA干擾技術,構建Chemerin基因缺陷慢病毒載體,并轉染小鼠ASMC,結果顯示,小鼠ASMC被成功分離培養,轉染培養48 h后,熒光顯微鏡觀察可見綠色熒光達90%以上,提示ASMC細胞慢病毒感染成功。本研究利用PDGF-BB對小鼠ASMC進行誘導,結果顯示,PDGF-BB可促進小鼠ASMC增殖。本研究結果顯示,Chemerin沉默組細胞中Chemerin基因和蛋白相對表達量均低于對照序列組、PDGF組和Sham組,進一步說明Chemerin沉默組細胞中Chemerin基因被抑制。細胞計數實驗和BrdU摻入法結果均顯示,Chemerin沉默組細胞數和吸光度A值均降低,表明Chemerin與小鼠ASMC增殖密切相關,抑制Chemerin基因表達可抑制ASMC增殖能力。
研究表明[13],絲裂原活化蛋白激酶(MAPK)信號通路在細胞增殖、凋亡、分化、遷移等多種細胞生物學過程中發揮重要作用。MAPK信號活化可促進自發性高血壓大鼠血管平滑肌細胞增殖[14],阻斷MAPK信號通路可抑制動脈損傷后血管平滑肌細胞增殖[15]。ERK1/2和JNK通路是MAPK信號通路中重要組成部分,與細胞增殖、分化和凋亡過程密切相關[16]。本研究顯示,Chemerin沉默組細胞中ERK1/2和JNK蛋白相對表達量均高于對照序列組和PDGF組,而低于Sham組,Chemerin沉默組細胞中p-ERK1/2和p-JNK蛋白相對表達量均低于對照序列組、PDGF組和Sham組,而對照序列組和PDGF組均高于Sham組,說明Chemerin沉默組細胞中ERK1/2和JNK信號通路被抑制,提示沉默Chemerin基因阻止小鼠ASMC增殖可能與抑制ERK1/2和JNK信號通路有關。
綜上所述,特異性沉默Chemerin基因可阻止ASMC的增殖作用,其機制可能與抑制ERK1/2和JNK信號通路有關,為冠狀動脈粥樣硬化發病機制研究提供了一定理論基礎,同時為該病基因防治提供了潛在靶點。
參 考 文 獻:
[1]WONG M C, ZHANG D X, WANG H H. Rapid emergence of atherosclerosis in Asia: a systematic review of coronary atherosclerotic heart disease epidemiology and implications for prevention and control strategies[J]. Curr Opin Lipidol, 2015,26(4): 257-269.
[2]ORG E, MEHRABIAN M, LUSIS A J. Unraveling the environmental and genetic interactions in atherosclerosis: central role of the gut microbiota[J]. Atherosclerosis, 2015, 241(2): 387-399.
[3]ZHANG Y N, XIE B D, SUN L, et al. Phenotypic switching of vascular smooth muscle cells in the ‘normal region’ of aorta from atherosclerosis patients is regulated by miR-145[J]. J Cell Mol Med, 2016, 20(6): 1049-1061.
[4]ZABEL B A, KWITNIEWSKI M, BANAS M, et al. Chemerin regulation and role in host defense[J]. Am J Clin Exp Immunol,2014, 3(1): 1-19.
[5]吳孟水, 寧翠利. Chemerin的研究進展[J]. 醫學綜述, 2016,22(11): 2102-2106.
[6]KOSTOPOULOS C G, SPIROGLOU S G, VARAKIS J N,et al. Chemerin and CMKLR1 expression in human arteries and periadventitial fat: a possible role for local chemerin in atherosclerosis?[J]. BMC Cardiovasc Disord, 2014, 14(3): 56-61.
[7]王欣琛. 冠心病的危險因素及二級預防策略[J]. 中西醫結合心腦血管病雜志, 2016, 14(11): 1303-1304.
[8]韓曉菲, 田慧. 脂肪因子chemerin的研究進展[J]. 解放軍醫學雜志, 2016, 41(5): 430-436.
[9]AKSAN G, ?NCI S, NAR G, et al. Association of serum chemerin levels with the severity of coronary artery disease in patients with metabolic syndrome[J]. Int J Clin Exp Med, 2014, 7(12): 5461-5468.
[10]AKAMATA K, ASANO Y, TANIGUCHI T, et al. Increased expression of chemerin in endothelial cells due to Fli1 de fi ciency may contribute to the development of digital ulcers in systemic sclerosis[J]. Rheumatology (Oxford), 2015, 54(7): 1308-1316.
[11]YU X, LI Z, CHEN G, et al. MicroRNA-10b induces vascular muscle cell proliferation through Akt pathway by targeting TIP30[J]. Curr Vasc Pharmacol, 2015, 13(5): 679-686.
[12]GASBARRINO K, MANTZOROS C, GORGUI J, et al.Circulating chemerin is associated with carotid plaque instability,whereas resistin is related to cerebrovascular symptomatology[J].Arterioscler Thromb Vasc Biol, 2016, 36(8): 1670-1678.
[13]ZHANG Y, PIZZUTE T, PEI M. A review of crosstalk between MAPK and Wnt signals and its impact on cartilage regeneration[J]. Cell Tissue Res, 2014, 358(3): 633-649.
[14]BOU DAOU G, LI Y, ANAND-SRIVASTAVA M B. Enhanced expression of Giα proteins contributes to the hyperproliferation of vascular smooth muscle cells from spontaneously hypertensive rats via MAP kinase- and PI3 kinase-independent pathways[J].Can J Physiol Pharmacol, 2016, 94(1): 49-58.
[15]ZHANG M, WANG X, BAI B, et al. Oxymatrine protects against sepsis-induced myocardial injury via inhibition of the TNF-α/p38-MAPK/caspase-3 signaling pathway[J]. Mol Med Rep, 2016,14(1): 551-559.
[16]王兆林, 王蓋昊, 于曉輝. MAPK 信號通路在肝細胞癌發生發展中的作用[J]. 現代腫瘤醫學, 2016, 24(2): 326-330.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
