拮抗植物病原真菌Streptomyces corchorusii AUH-1的分離與鑒定
2018-05-09 09:04:01章帥文吳志明李昆太
廣東農(nóng)業(yè)科學(xué) 2018年2期
關(guān)鍵詞:植物
章帥文,楊 勇,劉 群,吳志明,李昆太
(江西農(nóng)業(yè)大學(xué)生物科學(xué)與工程學(xué)院/江西省農(nóng)業(yè)微生物資源開發(fā)與利用工程實(shí)驗(yàn)室,江西 南昌 330045)
植物病害是造成農(nóng)作物生長(zhǎng)受阻、產(chǎn)量下降和品質(zhì)降低的主要原因之一。據(jù)統(tǒng)計(jì),全球每年因植物病害而造成的農(nóng)業(yè)產(chǎn)量下降比例就高達(dá)10%~16%[1]。植物病原菌主要包括細(xì)菌、病毒、真菌、線蟲等,而約3/4的植物病害是由植物病原真菌引起的[2]。植物病原真菌常借助侵染墊、附著胞和吸器等侵染結(jié)構(gòu)完成對(duì)寄主的侵染,并在寄主組織中大量生長(zhǎng),從而誘發(fā)植物真菌病害的發(fā)生[3-4]。例如,由稻瘟病菌(Magnaporthe grisea)和水稻紋枯病菌(Rhizoctonia solani)引發(fā)的水稻稻瘟病以及紋枯病,已成為全球性的水稻主要病害[5-6];尖孢鐮刀菌(Fusarium oxysporum)是一種最為常見(jiàn)的世界性的土傳病原真菌,可引起葫蘆科、香蕉、瓜類、豆科、茄類、棉花等100多種植物發(fā)生枯萎病[7-9];此外,由炭疽病菌(Colletotrichum)引發(fā)的植物炭疽病,也是造成花果和蔬菜等植物重大損失的一類常見(jiàn)真菌病害[10]。
利用拮抗微生物或其代謝產(chǎn)物進(jìn)行植物病害生防,具有環(huán)保、安全、無(wú)殘留等優(yōu)點(diǎn),且不易使病原菌產(chǎn)生抗藥性,而且很多拮抗微生物都具有對(duì)作物的促生作用,因此微生物生防顯示出越來(lái)越廣闊的應(yīng)用前景,成為目前研究開發(fā)的熱點(diǎn)。
黃麻鏈霉菌(Streptomyces corchorusii)是鏈霉菌屬中的一種,對(duì)黃瓜枯萎病[11]、草莓枯萎病[12]、草莓炭疽病[13]等多種植物病害均具有一定的抑制作用。本研究以西瓜枯萎病菌為指示菌,從土壤中分離篩選到1株對(duì)植物病原真菌具有良好拮抗作用的黃麻鏈霉菌AUH-1,采用生理生化方法和16S rDNA序列分析對(duì)其鑒定,以期為該拮抗菌后期的活性物質(zhì)檢測(cè)與生物防治應(yīng)用提供基礎(chǔ)。
1 材料與方法
1.1 試驗(yàn)材料
供試土樣:從江西南昌、贛州、鷹潭、萍鄉(xiāng)、福建武夷山、河北石家莊等地用五點(diǎn)取樣法采集不同類型土樣26個(gè)。
供試菌株:意大利青霉(Penicillium italicum)、西瓜枯萎病菌(Fusarrium oxysporum f.sp.niveum)、水稻紋枯病菌(Rhizoctonia solani)、大腸桿菌(Escherichia coli)、金黃色葡萄球菌(Staphylococcus aureus)、釀酒酵母(Saccharomyces cerevisiae)、枯草芽孢桿菌(Bacillus subtilis),均由本實(shí)驗(yàn)室保藏。
培養(yǎng)基:高氏一號(hào)培養(yǎng)基,PDA培養(yǎng)基,硫化氫(H2S)產(chǎn)生培養(yǎng)基(柴斯鈉培養(yǎng)基),纖維素分解培養(yǎng)基,明膠液化培養(yǎng)基,淀粉水解測(cè)定培養(yǎng)基,牛奶液化測(cè)定培養(yǎng)基,硝酸鹽還原培養(yǎng)基[14-15]。
主要試劑:Ezup柱式細(xì)菌基因組DNA提取試劑盒,Taq DNA聚合酶;DNA marker,PCR通用引物等。
主要儀器:2720 Thermal cycler PCR儀,3730-XL測(cè)序儀,F(xiàn)R980凝膠成像系統(tǒng),HC-2518R 冷凍高速離心機(jī),DYY-5電泳儀,光學(xué)顯微鏡,超凈工作臺(tái),恒溫培養(yǎng)箱等。
1.2 試驗(yàn)方法
1.2.1 拮抗放線菌菌株的分離篩選 放線菌菌株的分離:為了獲得豐富的放線菌資源,本研究從江西南昌、贛州、鷹潭、萍鄉(xiāng)、武夷、河北等地采集了26個(gè)不同類型的土樣。分別稱取10 g土樣加入裝有90 mL無(wú)菌水的三角瓶中制成10-1的土壤懸浮液,以無(wú)菌水梯度稀釋至10-3、10-4、10-5;吸取0.2 mL各梯度稀釋液,分別涂布在加有50 mg/L放線菌酮和50 mg/L重鉻酸鉀的高氏一號(hào)培養(yǎng)基平板上,30℃培養(yǎng)7 d;挑取呈放線菌菌落特征的單菌落,轉(zhuǎn)接于高氏一號(hào)培養(yǎng)基進(jìn)一步分離純化和保存。
放線菌菌株純化:挑取30℃培養(yǎng)7 d后呈放線菌菌落特征的單菌落,轉(zhuǎn)接到高氏一號(hào)培養(yǎng)基中進(jìn)一步分離純化,于20%甘油溶液中保存,-20℃存放備用。
采用平板對(duì)峙培養(yǎng)法進(jìn)行拮抗菌的初篩[16]:將待測(cè)病原真菌菌餅菌(6 mm)面朝下接種至中心位置,用滅菌后的牙簽挑取相同大小的供試放線菌倒置接種于距中心30 mm處,每個(gè)處理3次重復(fù),以只接植物病原真菌作為對(duì)照,28℃恒溫培養(yǎng),待對(duì)照組植物病原真菌菌絲鋪滿整個(gè)平板時(shí),測(cè)定實(shí)驗(yàn)組的抑菌帶寬度(R2)和指示菌菌落直徑(R1),并根據(jù)(R1-R2)/R1的比值大小,篩選出抑菌活性強(qiáng)的放線菌菌株。
拮抗放線菌的復(fù)篩:采用管碟法(牛津杯)進(jìn)行拮抗放線菌的復(fù)篩。將大腸桿菌和金黃色葡萄球菌的菌懸液與冷卻至50℃的牛肉膏蛋白胨培養(yǎng)基混合均勻,每100 mL培養(yǎng)基加入10 mL菌懸液,制成帶菌平板。類似地,釀酒酵母菌、西瓜枯萎病菌、枯草芽孢桿菌等以PDA培養(yǎng)基制成帶菌平板。等平板凝固后,在平板上放經(jīng)過(guò)滅菌的牛津杯,各取200 mL經(jīng)復(fù)篩培養(yǎng)基(可溶性淀粉20.0 g/L,KNO31.0 g/L,NaCl 0.5 g/L,MgSO40.5 g/L,K2HPO40.5 g/L,F(xiàn)eSO4·7H2O 0.01 g/L,pH 7.2~7.4)發(fā)酵的無(wú)菌發(fā)酵液吸入其中。細(xì)菌在37℃培養(yǎng)2 d,真菌在30℃培養(yǎng)2~3 d。然后測(cè)定抑菌圈直徑,挑選出具廣譜抗菌特性的拮抗放線菌。
1.2.2 拮抗菌株的鑒定 菌絲形態(tài)學(xué)觀察:采用平皿插片法,將拮抗菌株接種于高氏一號(hào)固體培養(yǎng)基上,28℃培養(yǎng)7~14 d,取插片在光學(xué)顯微鏡下觀察基內(nèi)菌絲和氣生菌絲的形態(tài)。
生理生化特征:參照《鏈霉菌鑒定手冊(cè)》[15]中的方法,進(jìn)行拮抗菌的碳源利用、明膠液化、纖維素分解、淀粉水解、牛奶凝固與胨化、硝酸鹽還原、產(chǎn)H2S測(cè)定等試驗(yàn)。
分子生物學(xué)鑒定:使用Ezup柱式細(xì)菌基因組DNA抽提試劑盒提取拮抗菌株的基因組DNA,采用通用引物27F(5′-AGTTTGATCMT GGCTCAG-3′)和1492R(5′-GGTTACCTTG TTACGACTT-3′)進(jìn)行PCR擴(kuò)增拮抗菌株的16SrDNA。PCR反應(yīng)條件為:94℃預(yù)變性4 min;94℃變性 45 s、55℃退火 45 s、72℃延伸 1 min,30個(gè)循環(huán);最后72℃延伸10 min。待PCR反應(yīng)完成后,取1μL PCR產(chǎn)物進(jìn)行1%瓊脂糖凝膠電泳檢測(cè),確認(rèn)PCR擴(kuò)增片段。將測(cè)序得到的16SrDNA序列在NCBI網(wǎng)站進(jìn)行BLAST比對(duì),用MEGA 4.1軟件以Neighbor-Joining法構(gòu)建系統(tǒng)發(fā)育樹,確定拮抗菌株的親緣關(guān)系和分類地位。
采用Excel 2003和DPS 7.5軟件進(jìn)行數(shù)據(jù)統(tǒng)計(jì)和分析,采用Photoshop CS5進(jìn)行圖像處理。
2 結(jié)果與分析
2.1 拮抗放線菌的分離篩選
2.1.1 拮抗放線菌的初篩 以西瓜枯萎病病原菌為供試菌株,采用對(duì)峙法對(duì)26個(gè)不同類型的土樣中初篩得到15株對(duì)西瓜枯萎病菌具有較強(qiáng)拮抗作用的菌株,結(jié)果見(jiàn)表1。其中,由抑菌帶的大小和(R1-R2)/R1的比值大小可知,菌株AUH-1對(duì)西瓜枯萎病菌的抑制能力最強(qiáng),其抑菌帶寬度和(R1-R2)/R1比值分別達(dá)到29.46(±0.48)mm和36.15%,如圖1(封三)所示。

表1 拮抗放線菌的初篩結(jié)果
2.1.2 拮抗放線菌的復(fù)篩 采用管碟法測(cè)定拮抗菌株AUH-1對(duì)不同供試菌株的抑制效果,挖取2 cm×1 cm大小的菌塊接種至裝量為40 mL/250 mL三角瓶的液體發(fā)酵培養(yǎng)基(蔗糖30 g/L,玉米淀粉20 g/L,玉米漿20 g/L,黃豆餅粉 10 g/L,(NH4)SO41 g/L,KH2PO40.25 g/L,MnCl20.05 g/L,MgSO41 g/L,NaCl 0.5 g/L,pH 7.2~7.4)中發(fā)酵,180 r/min、28℃下?lián)u床培養(yǎng)96 h,發(fā)酵液無(wú)菌過(guò)濾,備用。將大腸桿菌和金黃色葡萄球菌的菌懸液與冷卻至50℃的牛肉膏蛋白胨培養(yǎng)基混合均勻,每100 mL培養(yǎng)基加入10 mL菌懸液,制成帶菌平板。類似地,酵母菌、枯草芽孢桿菌、西瓜枯萎病菌、意大利青霉菌和水稻紋枯病菌等以PDA培養(yǎng)基制成帶菌平板。平板凝固后,在平板上放置經(jīng)過(guò)滅菌的牛津杯,各取200 μL無(wú)菌發(fā)酵液吸入其中。細(xì)菌在37℃培養(yǎng)2 d,真菌在30℃培養(yǎng)2~3 d。然后用十字交叉法測(cè)抑菌圈直徑。結(jié)果見(jiàn)表2。
2.2 菌株AUH-1的鑒定
2.2.1 菌株AUH-1的菌落特征 從圖2(封三)可以看出,菌株AUH-1的菌落正常、黃色、幾乎不產(chǎn)孢子,長(zhǎng)于基內(nèi),具有明顯的放線菌菌落特征。為了進(jìn)一步鑒定該菌株的種屬情況,對(duì)其開展了菌體形態(tài)學(xué)觀察、生理生化特征以及16S rDNA序列分析。

表2 菌株AUH-1發(fā)酵液對(duì)供試菌株的抑制效果
2.2.2 菌株AUH-1的菌絲形態(tài)特征 菌株AUH-1在PDA培養(yǎng)基上培養(yǎng)7 d后,在光學(xué)顯微鏡下觀察到的菌絲體形態(tài)如圖3(封三)所示。菌株AUH-1在PDA培養(yǎng)基上生長(zhǎng)發(fā)育良好,基內(nèi)菌絲和氣生菌絲較為豐富,少量氣生菌絲分化形成孢子。
2.2.3 菌株AUH-1的生理生化特征 表3為菌株AUH-1的碳源利用情況和生理生化特征。從表3可以看出,菌株AUH-1可以利用D-葡萄糖、D-甘露醇、蔗糖、麥芽糖、肌醇、D-果糖、和淀粉,不能利用D-木糖、山梨醇、甘油和阿拉伯糖;菌株AUH-1不能水解淀粉和分解纖維素,能使明膠液化以及牛奶凝固與胨化,能產(chǎn)硫化氫和還原硝酸鹽。

表3 菌株AUH-1的生理生化特征
2.2.4 菌株AUH-1的分子生物學(xué)鑒定 菌株AUH-1的基因組DNA經(jīng)PCR擴(kuò)增后得到1條約為1.4 kb的特征帶,經(jīng)測(cè)序其16S rDNA的序列長(zhǎng)度為1 491 bp。將該序列與NCBI數(shù)據(jù)庫(kù)中相應(yīng)的16S rDNA序列進(jìn)行Blast比對(duì),采用Neighbor-Joining方法構(gòu)建菌株AUH-1的系統(tǒng)發(fā)育樹(圖4),分析結(jié)果顯示菌株AUH-1與Streptomyces corchorusii的系列同源性達(dá)到99%。
3 結(jié)論與討論
微生物防治主要是利用有益的微生物,通過(guò)生物堿間的競(jìng)爭(zhēng)、抗生、寄生、溶菌及誘導(dǎo)抗性等作用,抑制病原物的存活[17]。目前,用于植物病害生防的微生物種類繁多,主要有真菌、細(xì)菌和放線菌等[18]。例如,作為植物根際促生菌中最大的細(xì)菌種群,假單胞桿菌(Pseudomonas spp.),尤其是熒光假單胞菌(Pseudomonas fluorscens),能有效地定殖在植物根際,控制植物病害、促進(jìn)植物生長(zhǎng),是植物病害生防研究中的一個(gè)重要類群[19]。
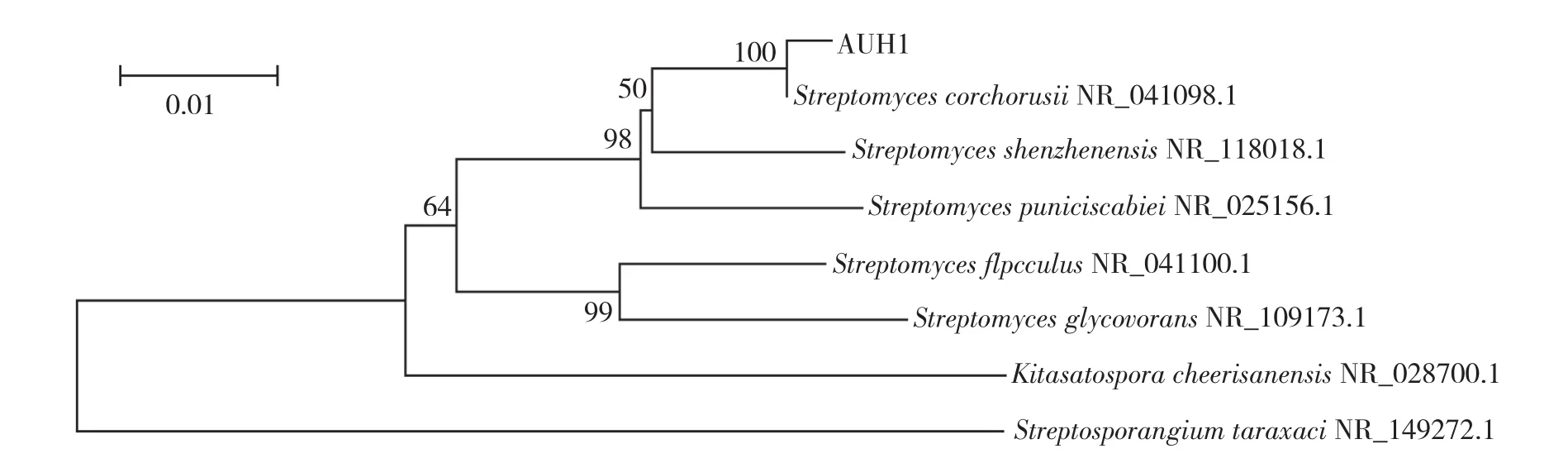
圖4 基于16S rDNA序列構(gòu)建的AUH-1菌株系統(tǒng)發(fā)育樹
放線菌是一類廣泛存在于土壤、植物體、淡水和海洋中的生防菌[20],種類繁多,具有復(fù)雜的次級(jí)代謝系統(tǒng),能產(chǎn)生諸多結(jié)構(gòu)新穎、生物活性顯著的次級(jí)代謝產(chǎn)物,是微生物農(nóng)藥的重要來(lái)源[21],在植物病害防治中具有舉足輕重的作用[22]。目前從自然界中發(fā)現(xiàn)上萬(wàn)種天然抗生素中,近70%是由放線菌產(chǎn)生的,其中鏈霉菌產(chǎn)生的抗生素占總數(shù)的52%[23-25]。鏈霉菌是放線菌門中種類最多的一個(gè)屬,是各種抗生素的主要產(chǎn)生來(lái)源[26-27]。鏈霉菌產(chǎn)生的抗生素類物質(zhì),能夠特異作用于某些病原菌,降低其生長(zhǎng)和繁殖速度,從而降低病害發(fā)作的頻率[28]。其中,由鏈霉菌產(chǎn)生的滅瘟素(Blasticidin S)、春日霉素(Kasugamycin)、多氧霉素(Polyoxins)、井岡霉素(Validamycin A)等,更是成功地商業(yè)化運(yùn)用于水稻、蔬菜和果類等的真菌病害防治中[29]。
本研究通過(guò)分離篩選得到1株拮抗鏈霉菌AUH-1,初步鑒定為黃麻鏈霉菌(Streptomyces corchorusii)。黃麻鏈霉菌作為鏈霉菌屬的一員,然目前國(guó)內(nèi)外對(duì)該菌的報(bào)道極少,特別是對(duì)其所產(chǎn)活性物質(zhì)結(jié)構(gòu)及抑菌機(jī)理等方面的研究更是少之甚少。燕照玲等[11]在開展植物病原真菌拮抗菌的分離篩選過(guò)程中,也獲得了1株對(duì)黃瓜枯萎病菌具有高效拮抗作用的Streptomyces corchorusii,但未有關(guān)于其活性物質(zhì)方面的研究。根據(jù)國(guó)外現(xiàn)有研究表明,Streptomyces corchorusii系蒽醌類(anthraquinone)抗腫瘤抗生素的主要產(chǎn)生菌[30],如拒霉素(resistomycin)和特曲霉素D(tetracenomycin D)[31]。值得注意的是,本研究是以植物病害生物防治為目的,分離篩選到1株植物病原真菌拮抗菌Streptomyces corchorusii AUH-1,該菌是否也會(huì)產(chǎn)生拒霉素和特曲霉素D等抗腫瘤抗生素,有待進(jìn)一步對(duì)其活性組分進(jìn)行結(jié)構(gòu)鑒定。另外,盡管菌株AUH-1具備的廣譜性拮抗植物病原真菌特點(diǎn)為其在不同類型植物病害上的生防應(yīng)用奠定了基礎(chǔ),但其抑菌機(jī)理以及在田間的防病效果,有待進(jìn)一步研究。
參考文獻(xiàn):
[1]Chakraborty S,Newton AC.Climate change,palnt diseases and food security:an over review[J].Plant Pathology,2011,60:2-14.
[2]蔡思琦,韓佳延,金黎明.海洋真菌ZJ353對(duì)植物病原菌的抑菌譜研究[J],山東化工,2016,45(17):25-26.
[3]Berendsen R L,Pieterse C M,Bakker P A.The rhizosphere microbiome and plant health[J].Trends in Plant Science,2012,17(8):478-86.
[4]Doornbos R F,Loon L C V,Bakker P A H M.Impact of root exudates and plant defense signaling on bacterial communities in the rhizosphere.[J].Agronomy for Sustainable Development,2012,32(1):227-243.
[5]Xiong Z Q,Tu X R,Wei S J,et al.In vitro antifungal activity of antifungalmycin 702,a new polyene macrolide antibiotic,against the rice blast fungus Magnaporthe grisea[J].Biotechnology Letters,2013,35(9):1475.
[6]Harikrishnan H,Shanmugaiah V,Balasubramanian N,et al.Antagonistic potential of native strain Streptomyces aurantiogriseus,VSMGT1014 against sheath blight of rice disease[J].World Journal of Microbiology & Biotechnology,2014,30(12):3149-3161.
[7]Summerell B A,Laurence M H,Liew E C Y,et al.Biogeography and phylogeography of Fusarium:a review[J].Fungal Diversity,2010,44(1):3-13.
[8]Zhao S,Liu D Y,Ling N,et al.Bio-organic fertilizer application significantly reduces the Fusarium oxysporum population and alters the composition of fungi communities of watermelon Fusarium wilt rhizosphere soil[J].Biology &Fertility of Soils,2014,50(5):765-774.
[9]Li C H,Zhao M W,Tang C M,et al.Population dynamics and identification of endophytic bacteria antagonistic toward plant-pathogenic fungi in cotton root[J].Microbial Ecology,2010,59(2):344-356.
[10]Araújo L,Gon?alves A E,Stadnik M J.Ulvan effect on conidial germination and appressoria formation of Colletotrichum gloeosporioides[J].Phytoparasitica,2014,42(5):631-640.
[11]燕照玲,孫虎,施艷,等.黃瓜枯萎病拮抗放線菌S24的分離、鑒定及發(fā)酵條件優(yōu)化[J].河南農(nóng)業(yè)科學(xué),2014,43(9):88-92.
[12]陳宏州,莊義慶,楊敬輝.黃麻鏈霉菌 NF0919菌株對(duì)草莓枯萎病的生防活性初探[J].江西農(nóng)業(yè)學(xué)報(bào),2014(11):54-57.
[13]楊敬輝,吉沐祥,陳宏州,等.黃麻鏈霉菌NF0919菌株發(fā)酵培養(yǎng)基的優(yōu)化[J].江西農(nóng)業(yè)學(xué)報(bào),2012,24(10):136-139.
[14]Shirling EB,Gottlieb D.Methods for characterization of Streptom-yces species[J].Int Journal of Syst Bacteriol,1966,16(3):313-340.
[15]中國(guó)科學(xué)院微生物研究所放線菌分類組.鏈霉菌鑒定手冊(cè)[M].北京:科學(xué)出版社,1975.
[16]傅本重,朱潔倩,郭新梅,等.核桃內(nèi)生菌XHE7對(duì)病原真菌的拮抗作用及種子發(fā)芽的影響[J].江西農(nóng)業(yè)大學(xué)學(xué)報(bào),2017,39(2):272-277.
[17]張麗,孫書娥.利用微生物防治植物病害研究進(jìn)展[J].農(nóng)藥研究與應(yīng)用,2010(6):10-13.
[18]Weller D M,Raaijmakers J M,Gardener B B M,et al.Microbial populations responsible for specific soil suppressiveness to plant pathogens 1[J].Annual Review of Phytopathology,2002,40(1):309.
[19]Sivasakthi S,Usharani G,Saranraj P.Biocontrol potentiality of plant growth promoting bacteria(PGPR)-Pseudomonas fluorescens and Bacillus subtilis:A review[J].African Journal of Agricultural Research,2014,9(16):1265-1277.
[20]安德榮.生物制藥的原理及方法 Ⅰ.抗生素的制備[J].中國(guó)生物工程雜志,2002(6):17.
[21]孔望君,蔣會(huì)芳,冉火苗,等.兩株土壤放線菌ZSM-1和ZSM-2的分離與鑒定[J].食品科學(xué),2016,37(11):114-119.
[22]彭衛(wèi)福,吳志明,陳未,等.拮抗多種植物病原真菌Streptomyces triostinicus C2的分離與鑒定[J].生物技術(shù)通報(bào),2016,32(7):106-111.
[23]Ayari A,Morakchi H,Djamila K G.Identification and antifungal activity of Streptomyces sp.S72 isolated from Lake Oubeira sediments in North- East of Algeria[J].African Journal of Biotechnology,2012,11(11):305-311.
[24]Liu X,Bolla K,Ashforth E J,et al.Systematicsguided bioprospecting for bioactive microbial natural products[J].Antonie Van Leeuwenhoek,2012,101(1):55-66.
[25]Couillerot O,Loqman S,Toribio A,et al.Purification of antibiotics from the biocontrol agent Streptomyces anulatus S37 by centrifugal partition chromatography[J].Journal of Chromatography B Analytical Technologies in the Biomedical & Life Sciences,2014,944:30-34.
[26]Kumari K S,Siddaiah V.Taxonomy,Identification and Biological Activities of a Novel Isolate of Streptomyces albus[J].Journal of Pharmacy Research,2011(12):4678-4680.
[27]Atta H M.Biochemical studies on antibiotic production from Streptomyces,sp.:Taxonomy,fermentation,isolation and biological properties[J].Journal of Saudi Chemical Society,2015,19(1):12-22.
[28]Song Y M,Dong X M,Cai Q M,et al.Isolation,Identification and Optimization of Fermentation Conditions of Streptomyces termitum Strain with Antimicrobial Activity against Xanthomonas oryzae pv.oryzae[J].Journal of Microbiology,2011,31(4):52-57.
[29]Kim B S,Hwang B K.Microbial fungicides in the control of plant diseases[J].Journal of Phytopathology,2007,155(12):641-653.
[30]Solanki R,Khanna M,Lal R.Bioactive compounds from marine actinomycetes[J].Indian Journal of Microbiology,2008,48(4):410-431.
[31]Adinarayana G,Venkateshan M R,Bapiraju V V S N K,et al.Cytotoxic compounds from the marine actinobacterium Streptomyces corchorusii,AUBN 1 /7 1[J].Russian Journal of Bioorganic Chemistry,2006,32(3):295-300.
猜你喜歡
少兒科學(xué)周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學(xué)閱讀指南·低年級(jí)版(2021年3期)2021-03-19 06:12:40
小天使·二年級(jí)語(yǔ)數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級(jí)語(yǔ)數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學(xué)苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
