語言認知神經科學研究中的腦溝形態分析*
2018-06-04 02:40:29蘇萌萌
心理科學進展 2018年6期
崔 新 蘇萌萌 舒 華
(1北京師范大學認知神經科學與學習國家重點實驗室, 北京 100875)
(2首都師范大學初等教育學院, 北京 100048)
1 引言
神經影像學技術的發展為研究者提供了豐富的大腦形態學測度。例如, 基于體素的形態分析法(Voxel-based morphometry, VBM)通過量化灰質和白質, 尋找有助于疾病早期診斷的生物標記(Ashburner & Friston, 2000)。隨著技術的進一步發展, 研究者開始關注更加精細的大腦形態測度,越來越多的研究者將灰質體積細分為皮層厚度和皮層表面積, 進而探討不同維度的腦結構指標與認知能力的相互關聯(Panizzon et al., 2009)。
近年來, 研究者開始關注皮層表面的形態特征, 腦溝是其中最具代表性的一種。已有的理論假設和研究結果認為腦溝形態并非任意分布, 而是底層生理組織建構以及發展的宏觀表征(Mangin,Jouvent, & Cachia, 2010)。具體表現在, 首先, 腦溝形成與遺傳因素聯系緊密。腦溝和腦回在胎兒10至16孕周顯現, 妊娠晚期已經出現與成人類似的皮層表面形態(Armstrong, Schleicher, Omran,Curtis, & Zilles, 1995), 而異常的遺傳因素會導致腦溝形態的變化(Fahim et al., 2012)。其次, 腦溝形態可能反映了皮層下的纖維連接方式(van Essen,1997; Hilgetag & Barbas, 2005), 有研究報告了腦回指數(gyrification index)和皮層下白質纖維束的各向異性(fractional anisotropy, FA)存在相關(Melbourne et al., 2014)。此外, 腦溝形態并非完全由灰質等其他形態測度決定。Trefler等人(2016)發現, 一天的不同時間段(time-of-day, TOD), 個體的皮層表面積、皮層厚度和灰質密度等指標會發生改變, 而腦溝深度則不會受到TOD的影響, 并且腦溝空間分布在發展過程中也保持相對穩定(Cachia et al.,2016)。
腦溝形態的特點契合了語言認知研究對神經生物表型的需要。一方面, 腦溝形態發生發展時間早, 為探究語言能力的產生及語言功能的偏側化提供了顯著的形態學標志(Habas et al., 2012;Kersbergen et al., 2016); 另一方面, 語言能力與遺傳因素(Platt et al., 2013)、白質纖維(Dubois et al., 2016; Hagoort, 2014)存在密切關系, 腦溝形態同時受到遺傳因素和白質纖維的影響, 從而為探究語言能力的底層神經機制提供了皮層水平的宏觀特征。目前的研究, 從認知層面考察腦溝特征的研究還相對較少, 對語言認知及其發展性障礙(例如閱讀障礙)的研究就更為不足。然而, 語言認知及其發展性障礙的病因學研究是人們關注的焦點, 研究者試圖從遺傳、環境、腦機制等層面全方位探討語言認知能力的發展。腦溝, 這一與遺傳、白質纖維等密切相關的形態學特征, 能夠為深入理解語言能力的正常與異常發展提供新的視角。綜上所述, 本文將首先介紹腦溝形態分析的方法, 然后綜述腦溝形態與語言能力有關的重要發現并展望未來可能的研究方向。
2 腦溝形態分析的方法
2.1 皮層復雜程度的量化
最早嘗試量化皮層表面特征的指標是腦回指數(gyrification index, GI) (Zilles, Armstrong, Schleicher,& Kretschmann, 1988), 即二維腦成像圖片(slices)中大腦內部周長(腦溝內的輪廓線長度)與外周周長的比值。隨著圖像處理技術的發展, 基于vertex計算的三維腦回指數在孤獨癥(Kates, Ikuta, &Burnette, 2009)、精神分裂(Harris et al., 2007)、雙相障礙(Mirakhur et al., 2009)等精神疾病的研究中得到了應用。
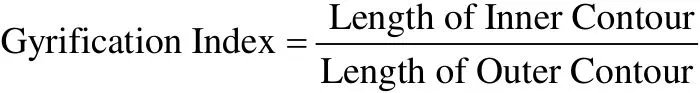
在腦回指數的基礎上, Cachia等人(2008)提出了腦溝指數(sulcal index, SI)的概念。腦溝指數包括總體腦溝指數(global sulcal index, g-SI)和局部腦溝指數(local sulcal index, l-SI)。總體腦溝指數指每個半球所有腦溝表面積和外部皮層表面積的比值; 局部腦溝指數指感興趣區域(region of interest,ROI)內所有腦溝表面積和外部皮層表面積的比值。腦溝指數沒有考慮灰質信號, 因此可以減少一些無關因素如藥物(Penttil? et al., 2008)對結果的影響。研究者發現異常的腦溝指數與幻聽型精神分裂(Cachia et al., 2008), 早發型精神分裂(Penttil? et al., 2008)有關。此外, 腦溝指數與老年人的注意、加工速度、記憶及語言等認知能力相關(Liu et al., 2011, 2013)。
盡管腦溝指數(SI)和腦回指數(GI)在計算方法上存在差異, 實際上均是對皮層復雜程度的量化, 并已經在神經疾病、認知能力的研究中積累了大量研究成果。但是, 復雜程度的分析仍不能確定腦溝形態的哪些方面發生了變化, 因此需要進一步量化目標腦溝的特征。
2.2 目標腦溝分析
Mangin等人(2004)提出新的算法:等位侵蝕技術(homotopic erosion technique)和裂隙探測器(crevasse detector), 自動識別T1加權像中的腦溝。兩種算法基于T1加權像, 將圖像分割為包含屬性鄰接圖(attributed relational graph, ARG)的簡單曲面(simple surfaces, SSs), 屬性鄰接圖中綜合了識別腦溝需要的所有信息。目標腦溝分析的主要步驟包括:預處理MR圖像; 分割大腦組織(包括腦脊液、灰質和白質); 在灰質與腦脊液界面分割腦溝并轉換為腦溝曲面; 最后識別并命名腦溝(Sun,Rivière, Poupon, Régis, & Mangin, 2007)。目前已有軟件包BrainVISA (http://brainvisa.info/) (Geffroy et al., 2011)能夠實現上述一系列處理過程(圖 1),處理之后可以得到定量的腦溝形態信息(表面積、長度、深度), 還可以定性分析腦溝的空間組織特征(相對位置和相對方向)。
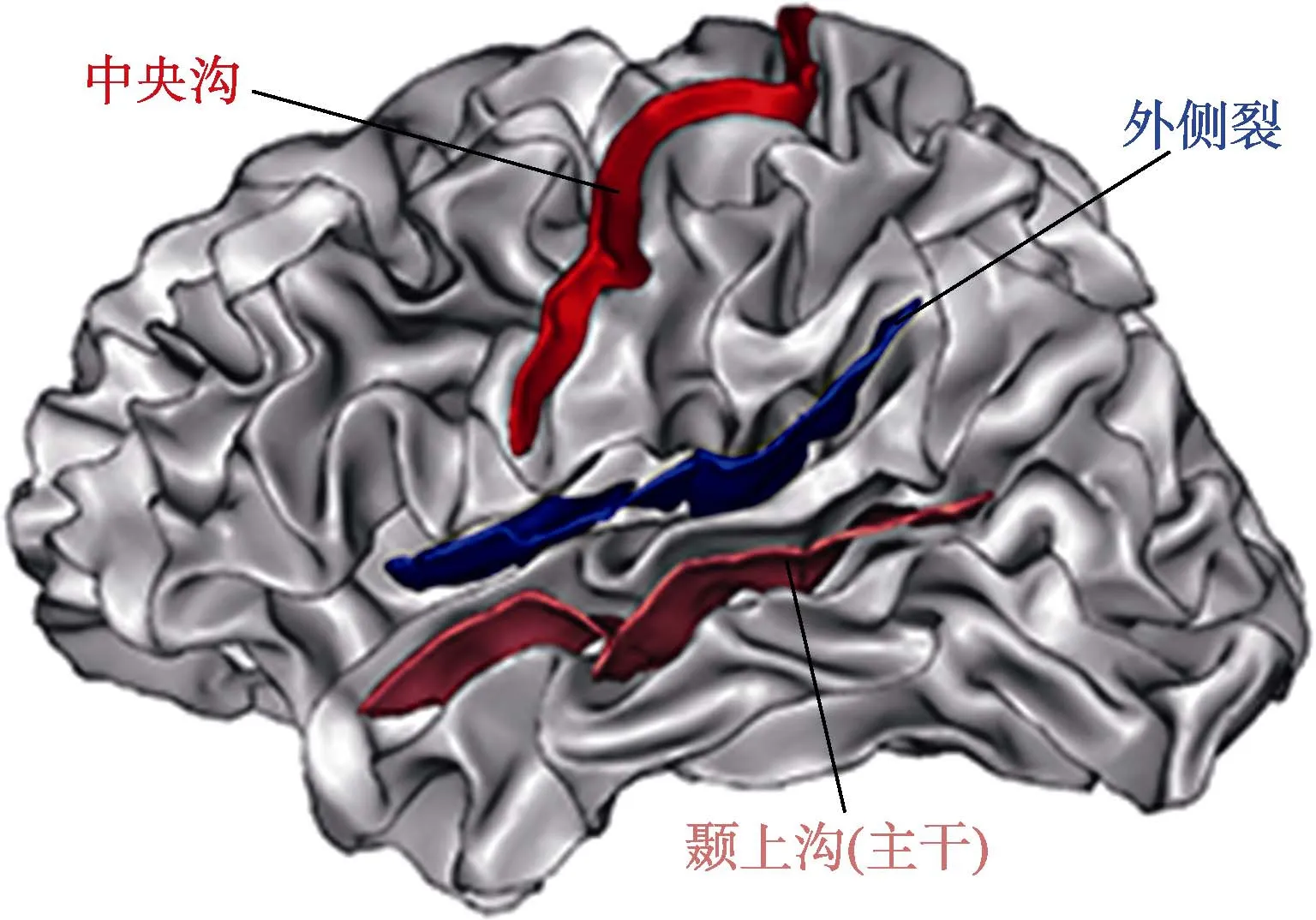
圖1 個體左半球的三條主要腦溝, 分別為中央溝(上)、外側裂(中)、顳上溝(下)。
目標腦溝形態的定量特征。腦溝曲面由外部邊界頂脊(top ridge), 內部邊界底脊(bottom ridge)和面(surface)組成, 其中“面”由相對的腦回“河床”(gyral banks)之間確定。頂脊和底脊的交叉點為面的終結點, 以頂脊和底脊包圍的面重構腦溝曲面。腦溝的表面積為腦溝曲面中“面”的面積。進一步地, 腦溝曲面的線性測度:長度和深度, 分別由溫度擴散模型(heat diffusion model)量化(Kochunov et al., 2010)。腦溝長度定義為沿著腦溝曲面排列的 100段等溫曲線長度的均值, 方向與腦溝底脊和頂脊共線; 腦溝深度的定義為沿腦溝曲面排列的 100段曲線的均值, 方向垂直于底脊和頂脊的連線。
目標腦溝的定性特征。腦溝空間組織(相對位置和相對方向等)分析通常包括中斷情況、腦溝之間是否相互連接, 腦溝末端的走向等。研究者要求多個評估者分別利用相同的編碼表評估每個被試的腦溝形態, 以評估的一致性程度確定腦溝的空間組織特征。以中斷為例, Borst等人(2016)讓3位研究者分別使用二分變量編碼(dichotomous variable code)評估兒童的顳枕溝(occipitotemporal sulcus,OTS)類型, 評估者之間的一致性達到 100%即可確定個體的顳枕溝類型為中斷或連續。目標腦溝的定性分析還應用于威廉姆斯綜合癥(Williams Syndrome, WS)患者的研究, 威廉姆斯綜合征患者右側顳平面顯著大于控制組, 其原因是患者右半球外側裂水平延伸而不是上升到頂葉(Eckert et al., 2006); 還有研究發現顳上溝(superior temporal sulcus, STS)的中斷數量存在左右半球的差異, 這種結構不對稱可能和語言偏側化相關(Ochiai et al., 2004)。
2.3 腦溝模式分析
腦回指數和腦溝指數能夠宏觀量化腦溝復雜程度, 目標腦溝分析旨在對特定腦溝形態進行描繪, 這兩種方法都無法量化多個腦溝的空間位置和排列關系。因此研究者提出了腦溝模式(sulcal pattern)的概念, 即腦溝的拓撲特征。
Lohmann, von Cramon 和 Colchester (2008)分析了腦溝深部的特點, 發現腦溝深部比淺層表現出較少的個體差異。每條腦溝沿其底部通常都能夠發現界限清楚的最大深度區域腦溝凹陷(sulcal pit)。Im等人(2010)發展出了基于腦溝凹陷建構腦溝模式的方法, 使用流域算法(watershed algorithm),將皮層分割為多個盆地(basin), 修剪較淺和較小的盆地之后, 每個盆地的最深處被標記為腦溝凹陷,每個腦溝凹陷在圖形表征中作為一個節點(node),如果腦溝盆地相鄰則使用無方向的邊(edge)相連,從而在選定的區域內得到腦溝圖形(sulcal graph)。腦溝圖形中主要包含節點的幾何特征(三維空間位置和腦溝盆地面積), 節點之間的幾何關系(反映腦溝之間的關系)以及節點的數量和節點間的路徑(圖形間的拓撲特征)。
研究者發現同卵雙生子的腦溝模式相似程度顯著高于樣本內隨機兩個個體間的腦溝模式(Im et al., 2011), 表明腦溝模式與遺傳因素之間存在密切聯系; 此外, 在多小腦回(Polymicrogyria,PMG)患者中, 完整語言能力患者和語言能力損傷患者相比, 頂葉腦溝模式與正常控制組更相似(Im, Pienaar et al., 2013); 還有研究報告了發展性閱讀障礙兒童和家族風險兒童與其控制組相比, 枕顳區域(parieto-temporal region)及頂顳區域(occipitotemporal region)腦溝模式出現異常(Im, Raschle,Smith, Ellen Grant, & Gaab, 2016)。
3 腦溝形態分析在語言認知領域的研究進展
3.1 腦溝形態的早期發展軌跡
兒童語言的早期發展為后期閱讀乃至社會認知的發展奠定了基礎(Catts, Fey, Ellis Weismer, &Bridges, 2014)。揭示語言相關腦結構的早期發展軌跡能夠幫助我們深入理解人類語言發展的底層神經機制。作為衡量大腦早期發育的重要指標,腦溝形態分析在早期語言發展中的應用受到研究者關注(Kersbergen et al., 2016)。
腦溝和腦回的形成能夠在有限空間內盡可能擴展皮層表面積(Striedter, Srinivasan, & Monuki,2015)。妊娠的3個月末期, 神經元開始最后的遷移并形成連接, 與此同時, 平滑的皮層表面開始形成復雜的腦溝和腦回。初級腦溝(primary sulci)很快延長和分支并形成二級(secondary sulci)和三級腦溝(tertiary sulci), 最終出生時皮層表面形成類似成人的復雜形態(Tallinen et al., 2016)。
研究者采用不同的方法描繪了18孕周到2歲時期腦溝形態的發展過程, 并在外側裂周區發現了顯著的皮層變化。Kasprian等人(2011)采集了197名宮內胎兒(18~37孕周)的MR圖像, 基于二維 MR圖像的冠狀面, 描繪每個胎兒大腦的輪廓線, 以輪廓線變異代表腦溝的復雜程度。結果發現超過2/3的胎兒表現出更大的左側顳葉, 94%的胎兒右側顳上溝(23孕周出現)比左側顳上溝(25孕周出現)出現的更早, 并且右側深于左側。Dubois,Benders和Borradori-Tolsa等人(2008)從35名早產兒(26~36孕周)MR圖像中重構了皮層(surface),并定義腦溝指數用于量化分析個體腦溝形態的復雜程度, 研究者發現額葉、顳葉等不同區域的腦溝指數存在差異, 而右半球比左半球更早的表現出復雜的腦溝形態。Habas等人(2012)基于皮層局部曲率, 描繪了宮內胎兒的腦溝形態發展。結果表明主要腦溝的出現時間存在差異, 外側裂(Sylvian fissure)最早出現曲率變化; 顳上溝在24周左右顯現, 而右側顳上溝比左側提早1周出現。出生后2年, 大腦經歷了發展最快的階段, 體積和表面積迅速增加(Gilmore et al., 2007)。Li等人(2014)追蹤了出生到 2歲嬰兒的腦溝形態變化, 發現從出生到 2歲顳上溝和頂枕溝右側深度大于左側, 而顳平面的大小和深度左側大于右側的現象。此外,研究者還發現顳上溝和顳平面在成人和新生兒群體間表現出類似的偏側化模式(Hill et al., 2010)。
嬰兒大腦是目前唯一能夠精通語言的“機器”(Dehaene-Lambertz & Spelke, 2015), 早期腦溝形態的發展軌跡與語言能力的關系主要表現在外側裂周區(peri-sylvian region)的形態改變。事實上,30孕周左右的早產兒已經具有區分音節的能力(Mahmoudzadeh, Wallois, Kongolo, Goudjil, &Dehaene-Lambertz, 2017), 有研究發現早產兒等價足月出生時期(term-equivalent age, TEA)的外側裂形態與其 2歲時的接受性語言有關(Kersbergen et al., 2016), 這些證據表明胎兒時期外側裂周區的腦溝形態變化可能與嬰兒的語言獲得和發展存在密切聯系。此外, 外側裂周區中, 顳葉腦溝形態變異較大(Leroy et al., 2011), 并且顳上溝在不同研究中報告了一致的發展進程, 這種結構變化可能支持了認知功能的發展。從出生后的語言功能來說, 嬰兒和成人的顳葉皮層對于語音刺激表現出相似的功能激活(Dehaene-Lambertz, 2017; Shultz,Vouloumanos, Bennett, & Pelphrey, 2014), 因此顳葉尤其是顳上溝的早期形態變化可能對于語言能力有重要作用。
3.2 腦溝形態與語言偏側化
人類最明顯的大腦功能偏側化為語言功能的偏側化(Sun & Walsh, 2006), 一直以來, 研究者試圖找到人類大腦獨特的結構特征, 以解釋人類的認知成就。顳平面(planum temporale, PT)和言語功能密切相關, 有研究報告了顳平面表面積的偏側化現象(Lyttelton et al., 2009), 但并非所有研究都發現了這種偏側化特征(Dorsaint-Pierre et al., 2006;Keller, Crow, Foundas, Amunts, & Roberts, 2009)并且靈長類動物中也報告了相似的現象(Hopkins &Nir, 2010)。隨著量化方法的提出, 研究者嘗試尋找人類獨特的腦溝偏側化特征。
語言偏側化的腦溝形態研究集中在腦溝凹陷上。腦溝凹陷是腦溝局部最深處, 被認為是大腦中形態變化的起始點, 并且其分布在個體間相對穩定(Lohmann, von Cramon, & Colchester, 2008;Régis et al., 2005)。Im 等人(2010)從 148 名成年中提取了腦溝凹陷, 分別在左右半球發現了48個和47個腦溝凹陷團塊(clusters), 腦溝凹陷的密度和空間分布表現出較小的個體差異, 并在顳上溝表現出數量和空間分布的半球間偏側化現象。Meng,Li, Lin, Gilmore和Shen (2014) 進一步追蹤了0~2歲嬰兒的腦溝凹陷發展, 發現了類似成人的腦溝凹陷分布, 并且這種分布在出生后兩年內保持相對穩定。研究者還發現, 嬰兒和成人顳上溝的腦溝凹陷分布具有一致的偏側化模式(Auzias, Brun,Deruelle, & Coulon, 2015; Im et al., 2010)。Leroy等人(2015)通過分析不同年齡人類和黑猩猩的 MR圖像, 進一步確定了人腦特有的左右不對稱區域(右側顯著深于左側), 將其命名為顳上不對稱凹陷(superior temporal asymmetrical pit, STAP)。
顳上溝深度的偏側化可能為語言能力的偏側化提供支持。首先, 顳上溝本身對語言能力有著重要作用, 左半球顳上溝特別地對言語刺激敏感(DeWitt & Rauschecker, 2012), 而發展進程上, 左側顳上溝比右側更晚出現, 這種左側發育延遲,可能為處理語言和運動刺激提供了更充分的結構可塑性(Dubois, Benders, Cachia et al., 2008)。不僅如此, 皮層下白質連接可能是腦溝形成的原因之一(Xu et al., 2010), 而顳葉皮層下存在復雜的白質纖維連接, 其中之一的弓形束(arcuate fasciculus,AF)與顳上不對稱凹陷在發展中表現出相似的特征:第一, 人類的弓形束和顳上不對稱凹陷遠大于黑猩猩; 第二, 兩者在生命早期均出現了偏側化現象(Dubois et al., 2009)。因此顳上不對稱凹陷可能與皮層下的白質纖維密度以及偏側化發展有關。鑒于弓形束在語音加工(Vandermosten, Boets,Wouters, & Ghesquière, 2012)和詞匯閱讀(Gullick& Booth, 2015)中的重要作用, 顳上不對稱凹陷很可能和語言認知能力密切相關。盡管顳上不對稱凹陷的功能尚不明確, 但其處于言語腹側通路音-義對應的核心區域(Liebenthal et al., 2010; Striem-Amit, Hertz, & Amedi, 2011), 并且黑猩猩中未發現這一結構, 因此研究者推測與顳平面相比, 顳上不對稱凹陷可能是更加人類特異的偏側化結構。
腦溝凹陷表現出了顯著偏側化現象和高遺傳度(Takerkart, Auzias, Brun, & Coulon, 2017), 為進一步探索大腦結構偏側化與語言功能偏側化之間的關系提供新的形態學測度。同時, 腦溝凹陷也是可靠的解剖標志(anatomical landmark) (Li, Guo,Nie, & Liu, 2010), 個體MR圖像中提取的腦溝凹陷在不同掃描儀、掃描序列和皮層重構工具之間具有很高的穩定性(Im, Lee et al., 2013)。因此, 研究者能夠基于腦溝凹陷建立腦溝模式, 進一步分析腦溝拓撲特征與認知能力之間的關系。
3.3 腦溝形態與發展性閱讀障礙
閱讀是語言認知研究最重要的方面之一, 閱讀能力是兒童多項能力發展的基石。發展性閱讀障礙(Developmental Dyslexia, DD)是學齡期兒童常見的一種學習障礙, 通常為智力、感知能力以及動機正常并且接受了充分教育的個體, 在準確流利識別字詞或拼寫時出現困難(Peterson & Pennington,2012)。發展性閱讀障礙的病因學基礎仍處于爭論之中, 有研究認為風險基因和異常的神經發育導致發展性閱讀障礙的發生(Skeide et al., 2016), 而另一些研究發現發展性閱讀障礙大腦的功能和結構異常來源于缺乏閱讀經驗(Clark et al., 2014; Olulade,Napoliello, & Eden, 2013)。
為了解決這一爭論, 一種方法是加入新對照組(Xia, Hoeft, Zhang, & Shu, 2016), 通過比較閱讀障礙組、年齡控制組和閱讀水平控制組來解釋DD腦結構異常的原因; 另一種方法是招募學前兒童(Raschle et al., 2017; Raschle, Stering, Meissner, &Gaab, 2014; Vandermosten et al., 2015; Wang et al.,2017)。學前兒童尚未接受系統的閱讀訓練, 通過比較閱讀障礙家族風險兒童和年齡控制組, 能夠發現相對獨立于閱讀經驗的大腦功能和結構異常。Raschle, Chang和Gaab (2011), Raschle等人(2017)報告了學前DD風險兒童, 枕顳區域和顳頂區域灰質體積的下降, 這表明 DD的大腦結構異常可能在學習閱讀之前已經存在。
最新一項研究中, Im等人(2016)對DD兒童及其年齡控制組和學前 DD風險兒童及其年齡控制組的腦溝模式進行了比較。研究者基于枕顳和頂顳區域的腦溝凹陷建立腦溝圖形, 每個正常被試與其他正常被試之間計算一個平均相似度每個 DD和所有正常被試計算一個平均相似度, 通過比較ST′和ST′,D可以得到DD與正常控制組的組間差異(ST′vs.ST′,D)。同理可以比較風險兒童和非風險兒童之間的相似度結果表明DD兒童與正常兒童相比以及學前風險兒童和非風險兒童相比, 均表現出腦溝模式的顯著差異; 腦溝模式的跨年齡比較中, 非風險兒童和學后正常兒童表現出了較高一致性, 而學前風險兒童和 DD表現出了較高的一致性。腦溝相似度和行為成績的相關表明, DD腦溝模式與控制組相似度越高, 其詞匯和語言加工測驗的成績越好。
鑒于腦溝模式出生時已經基本形成, 閱讀障礙兒童、風險兒童和其對應控制組的腦溝模式差異可能反映了早期大腦結構存在的異常, 甚至可能追溯到胎兒發育時期。研究者推測異常的腦溝模式可能受到異常遺傳因素和皮層下白質連接的影響。首先, DD風險基因(如DCDC2, KIAA0319)等對于大腦發展尤其是神經元遷移有著重要作用(Peterson & Pennington, 2012), 神經元的遷移是腦溝形成的影響因素之一(Striedter et al., 2015),發育早期風險遺傳因素可能會通過影響神經元遷移, 進而導致腦溝模式的異常。其次, 皮層下白質纖維束的密度和張力可能驅動了腦溝形成(Zilles,Palomero-Gallagher, & Amunts, 2013)。枕顳皮層的視覺詞匯加工區(visual word form area, VWFA)對于閱讀有著重要作用(Bouhali et al., 2014; Dehaene,Cohen, Morais, & Kolinsky, 2015), 而有證據表明閱讀學習之前的白質纖維連接支持了VWFA的產生(Saygin et al., 2016), 因此, 學前風險兒童枕顳腦溝模式的差異可能反映了發展早期白質連接的異常。近期Williams, Juranek, Cirino和Fletcher(2017)報告了 DD左側枕顳區域腦回指數的增加,研究者推測增加的皮層復雜程度可能反映了皮層局部信息交換能力的提升, 但同時以長距離連接(如下額枕束等)減弱為代價, 從而影響了DD的閱讀能力。
正常群體的研究同樣在枕顳區域發現了腦溝形態與閱讀能力之間存在關聯。左側枕顳溝的空間分布和閱讀準確性相關(Cachia et al., 2017), 而腦溝的空間分布被認為在出生前已經形成并且較少受到出生后神經可塑性的影響(Cachia et al.,2016), 研究者推測左側枕顳溝和閱讀能力的關系可能反映了皮層細胞構筑(Cytoarchitecture)和皮層下白質纖維連接對于閱讀能力的影響(Dubois et al., 2016; Weiner et al., 2017)。
總之, 異常群體和正常群體的腦溝形態研究為探討早期大腦發育與語言認知能力之間的關系提供了初步證據。同時, 腦溝的發生發展機制與編碼蛋白質的基因、微核糖核酸(microRNA)等因素密切聯系(Sun & Hevner, 2014), 為探討語言發展及障礙的遺傳學機制提供了新的中介表型。
4 總結與展望
Klee和Stokes (2011)提出的語言發展模型中包含了遺傳因素、環境刺激、神經生物基礎、認知機制等多種成分和過程, 語言的發展并非單一因素決定而受到多種過程的影響。腦溝形態為研究者提供了更加精細的神經生物學指標, 已有研究表明其在語言認知研究中的應用潛力:首先, 早期腦溝形態變化顯著, 能夠作為發育早期的神經影像學指標, 探討語言認知能力與大腦結構發展及遺傳因素的關系(Sun et al., 2005); 其次, 腦溝凹陷的偏側化可能提供了語言功能偏側化的結構基礎, 支持了語言認知理論的模型; 最后, 腦溝模式及腦溝的空間分布較少受到出生后神經可塑性的影響, 為揭示發展性閱讀障礙等認知缺陷的病因學基礎提供了更接近遺傳因素的形態學測度。
事實上, 腦溝形態分析在神經和精神疾病中已經有了廣泛的應用, 如孤獨癥(Libero, DeRamus,Deshpande, & Kana, 2014; Wallace et al., 2013), 威廉姆斯綜合征(Fahim et al., 2012), 強迫癥(Shim et al., 2009), 口吃(Cykowski et al., 2008), 阿爾茲海默癥(Hamelin et al., 2015)等。這些腦溝形態異常通常與遺傳和神經發育的擾動有關, 反映了疾病早期的病理學特征, 為探討神經及精神疾病的病因學基礎, 關聯認知損傷與大腦結構異常以及預測疾病發生發展提供了早期生物標志。
相比較而言, 腦溝形態在語言認知領域內的應用正處于起步階段, 仍有很多問題有待進一步研究。首先, 腦溝形態與遺傳因素的關系。相關問題包括, 語言相關的遺傳因素影響了哪些腦溝形態特征, 及早期腦溝形態的偏側化是否與這些基因表達有關等。第二, 腦溝形態與語言等認知能力之間的關系。腦溝本身并不包含神經元, 也不承擔特定認知功能, 因此, 研究者面臨相繼的兩個問題, 首先, 腦溝形態是否與語言認知能力相關; 其次, 假設相關存在, 腦溝形態反映了何種生理過程進而影響語言認知能力。這些問題需要研究者基于腦溝的發生發展機制及細胞構筑等研究謹慎推測。第三, 腦溝形態與后天發展和環境。大腦在出生后仍不斷發展, 兒童及青少年時期是個體認知能力發展的關鍵時期, 這一時期語言發展和腦溝形態發展軌跡之間存在何種關系尚不明確, 而成熟過程和后天環境因素如何塑造腦溝形態仍有待研究。闡明腦溝形態與遺傳、認知和環境三者的相互作用, 有助于研究者深入了解語言發展認知神經機制。此外, 語言認知研究應考慮到文化差異的影響, 來自不同國家地區的大樣本研究有助于發現語言能力和腦溝形態的關聯,抑或探討不同文化背景下群體之間可能存在的差異。最后, 腦溝形態分析方法的優化能夠提供更加豐富神經影像指標, 為揭示發展性閱讀障礙的病因學基礎, 探索遺傳因素與語言發展及語言障礙之間的關系, 并進一步為建立發展性閱讀障礙等語言障礙的早期診斷模型提供支持。
Armstrong, E., Schleicher, A., Omran, H., Curtis, M., & Zilles,K. (1995). The ontogeny of human gyrification.Cerebral Cortex, 5(1), 56–63.
Ashburner, J., & Friston, K. J. (2000). Voxel-based morphometry—the methods.NeuroImage, 11(6), 805–821.
Auzias, G., Brun, L., Deruelle, C., & Coulon, O. (2015). Deep sulcal landmarks: Algorithmic and conceptual improvements in the definition and extraction of sulcal pits.NeuroImage,111, 12–25.
Borst, G., Cachia, A., Tissier, C., Ahr, E., Simon, G., &Houdé, O. (2016). Early cerebral constraints on reading skills in school-age children: An MRI study.Mind, Brain,and Education, 10(1), 47–54.
Bouhali, F., de Schotten, M. T., Pinel, P., Poupon, C., Mangin, J.F., Dehaene, S., & Cohen, L. (2014). Anatomical connections of the visual word form area.Journal of Neuroscience, 34(46), 15402–15414.
Cachia, A., Borst, G., Tissier, C., Fisher, C., Plaze, M., Gay,O., ... Raznahan, A. (2016). Longitudinal stability of the folding pattern of the anterior cingulate cortex during development.Developmental Cognitive Neuroscience, 19,122–127.
Cachia, A., Paillère-Martinot, M. L., Galinowski, A., Januel,D., de Beaurepaire, R., Bellivier, F., ... Martinot, J. L.(2008). Cortical folding abnormalities in schizophrenia patients with resistant auditory hallucinations.NeuroImage,39(3), 927–935.
Cachia, A., Roell, M., Mangin, J. F., Sun, Z. Y., Jobert, A.,Braga, L., ... Borst, G. (2017). How interindividual differences in brain anatomy shape reading accuracy.Brain Structure and Function, 1–12
Catts, H. W., Fey, M. E., Ellis Weismer, S., & Bridges, M. S.(2014). The relationship between language and reading abilities. In J. B. Tomblin & M. A. Nippold (Eds.),Understanding individual differences in language development across the school years(pp. 144–165). New York: Psychology Press.
Clark, K. A., Helland, T., Specht, K., Narr, K. L., Manis, F.R., Toga, A. W., & Hugdahl, K. (2014). Neuroanatomical precursors of dyslexia identified from pre-reading through to age 11.Brain, 137(12), 3136–3141.
Cykowski, M. D., Kochunov, P. V., Ingham, R. J., Ingham, J.C., Mangin, J. F., Rivière, D., ... Fox, P. T. (2008).Perisylvian sulcal morphology and cerebral asymmetry patterns in adults who stutter.Cerebral Cortex, 18(3),571–583.
Dehaene, S., Cohen, L., Morais, J., & Kolinsky, R. (2015).Illiterate to literate: Behavioural and cerebral changes induced by reading acquisition.Nature Review Neuroscience,16(4), 234–244.
Dehaene-Lambertz, G. (2017). The human infant brain: A neural architecture able to learn language.Psychonomic Bulletin and Review, 24(1), 48–55.
Dehaene-Lambertz, G., & Spelke, E. S. (2015). The infancy of the human brain.Neuron, 88(1), 93–109.
DeWitt, I., & Rauschecker, J. P. (2012). Phoneme and word recognition in the auditory ventral stream.Proceedings of the National Academy of Sciences of the United States of America, 109(8), E505–E514.
Dorsaint-Pierre, R., Penhune, V. B., Watkins, K. E., Neelin,P., Lerch, J. P., Bouffard, M., & Zatorre, R. J. (2006).Asymmetries of the planum temporale and Heschl's gyrus:Relationship to language lateralization.Brain, 129(5),1164–1176.
Dubois, J., Benders, M., Borradori-Tolsa, C., Cachia, A.,Lazeyras, F., Ha-Vinh Leuchter, R., ... Hüppi, P. S. (2008).Primary cortical folding in the human newborn: An early marker of later functional development.Brain, 131(8),2028–2041.
Dubois, J., Benders, M., Cachia, A., Lazeyras, F., Ha-Vinh Leuchter, R., Sizonenko, S. V., ... Hüppi, P. S. (2008).Mapping the early cortical folding process in the preterm newborn brain.Cerebral Cortex, 18(6), 1444–1454.
Dubois, J., Hertz-Pannier, L., Cachia, A., Mangin, J. F., Le Bihan, D., & Dehaene-Lambertz, G. (2009). Structural asymmetries in the infant language and sensori-motor networks.Cerebral Cortex, 19(2), 414–423.
Dubois, J., Poupon, C., Thirion, B., Simonnet, H., Kulikova,S., Leroy, F., ... Dehaene-Lambertz, G. (2016). Exploring the early organization and maturation of linguistic pathways in the human infant brain.Cerebral Cortex, 26(5), 2283–2298.
Eckert, M. A., Galaburda, A. M., Karchemskiy, A., Liang, A.,Thompson, P., Dutton, R. A., ... Reiss, A. L. (2006).Anomalous sylvian fissure morphology in Williams syndrome.NeuroImage, 33(1), 39–45.
Fahim, C., Yoon, U., Nashaat, N. H., Khalil, A. K.,El-Belbesy, M., Mancini-Marie, A., ... Meguid, N. (2012).Williams syndrome: A relationship between genetics,brain morphology and behaviour.Journal of Intellectual Disability Research, 56(9), 879–894.
Geffroy, D., Rivière, D., Denghien, I., Souedet, N., Laguitton,S., & Cointepas, Y. (2011).BrainVISA: A complete software platform for neuroimaging.Paper presented at the Python in neuroscience workshop.
Gilmore, J. H., Lin, W. L., Prastawa, M. W., Looney, C. B.,Vetsa, Y. S. K., Knickmeyer, R. C., ... Gerig, G. (2007).Regional gray matter growth, sexual dimorphism, and cerebral asymmetry in the neonatal Brain.Journal of Neuroscience, 27(6), 1255–1260.
Gullick, M. M., & Booth, J. R. (2015). The direct segment of the arcuate fasciculus is predictive of longitudinal reading change.Developmental Cognitive Neuroscience, 13, 68–74.
Habas, P. A., Scott, J. A., Roosta, A., Rajagopalan, V., Kim,K., Rousseau, F., ... Studholme, C. (2012). Early folding patterns and asymmetries of the normal human brain detected from in utero MRI.Cerebral Cortex,22(1), 13–25.
Hagoort, P. (2014). Nodes and networks in the neural architecture for language: Broca's region and beyond.Current Opinion in Neurobiology, 28, 136–141.
Hamelin, L., Bertoux, M., Bottlaender, M., Corne, H.,Lagarde, J., Hahn, V., ... Sarazin, M. (2015). Sulcal morphology as a new imaging marker for the diagnosis of early onset Alzheimer's disease.Neurobiology of Aging,36(11), 2932–2939.
Harris, J. M., Moorhead, T. W. J., Miller, P., McIntosh, A. M.,Bonnici, H. M., Owens, D. G. C., ... Lawrie, S. M. (2007).Increased prefrontal gyrification in a large high-risk cohort characterizes those who develop schizophrenia and reflects abnormal prefrontal development.Biological Psychiatry,62(7), 722–729.
Hilgetag, C. C., & Barbas, H. (2005). Developmental mechanics of the primate cerebral cortex.Anatomy and Embryology,210(5-6), 411–417.
Hill, J., Dierker, D., Neil, J., Inder, T., Knutsen, A., Harwell,J., ... van Essen, D. (2010). A surface-based analysis of hemispheric asymmetries and folding of cerebral cortex in term-born human infants.Journal of Neuroscience, 30(6),2268–2276.
Hopkins, W. D., & Nir, T. M. (2010). Planum temporale surface area and grey matter asymmetries in chimpanzees(Pan troglodytes): The effect of handedness and comparison with findings in humans.Behavioural Brain Research,208(2), 436–443.
Im, K., Jo, H. J., Mangin, J. F., Evans, A. C., Kim, S. I., &Lee, J. M. (2010). Spatial distribution of deep sulcal landmarks and hemispherical asymmetry on the cortical surface.Cerebral Cortex, 20(3), 602–611.
Im, K., Lee, J. M., Jeon, S., Kim, J. H., Seo, S. W., Na, D. L.,& Grant, P. E. (2013). Reliable identification of deep sulcal pits: The effects of scan session, scanner, and surface extraction tool.PLoS One,8(1), e53678.
Im, K., Pienaar, R., Lee, J. M., Seong, J. K., Choi, Y. Y., Lee,K. H., & Grant, P. E. (2011). Quantitative comparison and analysis of sulcal patterns using sulcal graph matching: A twin study.NeuroImage, 57(3), 1077–1086.
Im, K., Lee, J. M., Jeon, S., Kim, J. H., Seo, S. W., Na, D. L.,& Grant, P. E. (2013). Reliable identification of deep sulcal pits: the effects of scan session, scanner, and surface extraction tool.PloS One, 8(1),e53678.
Im, K., Pienaar, R., Paldino, M. J., Gaab, N., Galaburda, A.M., & Grant, P. E. (2013). Quantification and discrimination of abnormal sulcal patterns in polymicrogyria.Cerebral Cortex, 23(12), 3007–3015.
Im, K., Raschle, N. M., Smith, S. A., Ellen Grant, P., & Gaab, N.(2016). Atypical sulcal pattern in children with developmental dyslexia and at-risk kindergarteners.Cerebral Cortex,26(3), 1138–1148.
Kasprian, G., Langs, G., Brugger, P. C., Bittner, M., Weber,M., Arantes, M., & Prayer, D. (2011). The prenatal origin of hemispheric asymmetry: An in utero neuroimaging study.Cerebral Cortex, 21(5), 1076–1083.
Kates, W. R., Ikuta, I., & Burnette, C. P. (2009). Gyrification patterns in monozygotic twin pairs varying in discordance for autism.Autism Research, 2(5), 267–278.
Keller, S. S., Crow, T., Foundas, A., Amunts, K., & Roberts,N. (2009). Broca’s area: Nomenclature, anatomy, typology and asymmetry.Brain and Language, 109(1), 29–48.
Kersbergen, K. J., Leroy, F., I?gum, I., Groenendaal, F., de Vries, L. S., Claessens, N. H. P., ... Benders, M. J. N. L.(2016). Relation between clinical risk factors, early cortical changes, and neurodevelopmental outcome in preterm infants.NeuroImage, 142, 301–310.
Klee, T., & Stokes, S. F. (2011). Language development. InChild psychology and psychiatry(pp. 45–50). Chichester:John Wiley & Sons.
Kochunov, P., Glahn, D. C., Fox, P. T., Lancaster, J. L.,Saleem, K., Shelledy, W., ... Rogers, J. (2010). Genetics of primary cerebral gyrification: Heritability of length, depth and area of primary sulci in an extended pedigree of Papio baboons.NeuroImage, 53(3), 1126–1134.
Leroy, F., Cai, Q., Bogart, S. L., Dubois, J., Coulon, O.,Monzalvo, K., ... Dehaene-Lambertz, G. (2015). New human-specific brain landmark: The depth asymmetry of superior temporal sulcus.Proceedings of the National Academy of Sciences of the United States of America,112(4), 1208–1213.
Leroy, F., Glasel, H., Dubois, J., Hertz-Pannier, L., Thirion,B., Mangin, J. F., & Dehaene-Lambertz, G. (2011). Early maturation of the linguistic dorsal pathway in human infants.Journal of Neuroscience, 31(4), 1500–1506.
Li, G., Guo, L., Nie, J. X., & Liu, T. M. (2010). An automated pipeline for cortical sulcal fundi extraction.Medical Image Analysis, 14(3), 343–359.
Li, G., Wang, L., Shi, F., Lyall, A. E., Lin, W., Gilmore, J. H.,& Shen, D. (2014). Mapping longitudinal development of local cortical gyrification in infants from birth to 2 years of age.Journal of Neuroscience, 34(12), 4228–4238.
Libero, L. E., DeRamus, T. P., Deshpande, H. D., & Kana, R.K. (2014). Surface-based morphometry of the cortical architecture of autism spectrum disorders: Volume, thickness,area, and gyrification.Neuropsychologia, 62, 1–10.
Liebenthal, E., Desai, R., Ellingson, M. M., Ramachandran,B., Desai, A., & Binder, J. R. (2010). Specialization along the left superior temporal sulcus for auditory categorization.Cerebral Cortex, 20(12), 2958–2970.
Liu, T., Sachdev, P. S., Lipnicki, D. M., Jiang, J. Y., Geng, G.Q., Zhu, W. L., ... Wen, W. (2013). Limited relationships between two-year changes in sulcal morphology and other common neuroimaging indices in the elderly.NeuroImage,83, 12–17.
Liu, T., Wen, W., Zhu, W. L., Kochan, N. A., Trollor, J. N.,Reppermund, S., ... Sachdev, P. S. (2011). The relationship between cortical sulcal variability and cognitive performance in the elderly.NeuroImage, 56(3), 865–873.
Lohmann, G., von Cramon, D. Y., & Colchester, A. C. F.(2008). Deep sulcal landmarks provide an organizing framework for human cortical folding.Cerebral Cortex,18(6), 1415–1420.
Lyttelton, O. C., Karama, S., Ad-Dab'bagh, Y., Zatorre, R. J.,Carbonell, F., Worsley, K., & Evans, A. C. (2009). Positional and surface area asymmetry of the human cerebral cortex.NeuroImage, 46(4), 895–903.
Mahmoudzadeh, M., Wallois, F., Kongolo, G., Goudjil, S., &Dehaene-Lambertz, G. (2017). Functional maps at the onset of auditory inputs in very early preterm human neonates.Cerebral Cortex, 27(4), 2500–2512.
Mangin, J. F., Jouvent, E., & Cachia, A. (2010). In-vivo measurement of cortical morphology: Means and meanings.Current Opinion in Neurology, 23(4), 359–367.
Mangin, J. F., Rivière, D., Cachia, A., Duchesnay, E.,Cointepas, Y., Papadopoulos-Orfanos, D., ... Régis, J.(2004). A framework to study the cortical folding patterns.NeuroImage, 23, S129–S138.
Melbourne, A., Kendall, G. S., Cardoso, M. J., Gunny, R.,Robertson, N. J., Marlow, N., & Ourselin, S. (2014).Preterm birth affects the developmental synergy between cortical folding and cortical connectivity observed on multimodal MRI.NeuroImage, 89, 23–34.
Meng, Y., Li, G., Lin, W. L., Gilmore, J. H., & Shen, D. G.(2014). Spatial distribution and longitudinal development of deep cortical sulcal landmarks in infants.NeuroImage,100, 206–218.
Mirakhur, A., Moorhead, T. W. J., Stanfield, A. C., McKirdy,J., Sussmann, J. E. D., Hall, J., ... McIntosh, A. M. (2009).Changes in gyrification over 4 years in bipolar disorder and their association with the brain-derived neurotrophic factor valine 66 methionine variant.Biological Psychiatry,66(3), 293–297.
Ochiai, T., Grimault, S., Scavarda, D., Roch, G., Hori, T.,Riviere, D., ... Régis, J. (2004). Sulcal pattern and morphology of the superior temporal sulcus.NeuroImage,22(2), 706–719.
Olulade, O. A., Napoliello, E. M., & Eden, G. F. (2013).Abnormal visual motion processing is not a cause of dyslexia.Neuron, 79(1), 180–190.
Panizzon, M. S., Fennema-Notestine, C., Eyler, L. T.,Jernigan, T. L., Prom-Wormley, E., Neale, M., ... Kremen,W. S. (2009). Distinct genetic influences on cortical surface area and cortical thickness.Cerebral Cortex,19(11), 2728–2735.
Penttil?, J., Paillére-Martinot, M. L., Martinot, J. L., Mangin,J. F., Burke, L., Corrigall, R., ... Cachia, A. (2008). Global and temporal cortical folding in patients with early-onset schizophrenia.Journal of the American Academy of Child and Adolescent Psychiatry, 47(10), 1125–1132.
Peterson, R. L., & Pennington, B. F. (2012). Developmental dyslexia.The Lancet, 379(9830), 1997–2007.
Platt, M. P., Adler, W. T., Mehlhorn, A. J., Johnson, G. C.,Wright, K. A., Choi, R. T., ... Rosen, G. D. (2013).Embryonic disruption of the candidate dyslexia susceptibility gene homologKiaa0319-likeresults in neuronal migration disorders.Neuroscience, 248, 585–593.
Régis, J., Mangin, J. F., Ochiai, T., Frouin, V., Riviére, D.,Cachia, A., ... Samson, Y. (2005). “Sulcal Root” generic model: A hypothesis to overcome the variability of the human cortex folding patterns.Neurologia Medico-Chirurgica,45(1), 1–17.
Raschle, N. M., Becker, B. L. C., Smith, S., Fehlbaum, L. V.,Wang, Y. Y., & Gaab, N. (2017). Investigating the influences of language delay and/or familial risk for dyslexia on brain structure in 5-year-olds.Cerebral Cortex,27(1), 764–776.
Raschle, N. M., Chang, M., & Gaab, N. (2011). Structural brain alterations associated with dyslexia predate reading onset.NeuroImage, 57(3), 742–749.
Raschle, N. M., Stering, P. L., Meissner, S. N., & Gaab, N.(2014). Altered neuronal response during rapid auditory processing and its relation to phonological processing in prereading children at familial risk for dyslexia.Cerebral Cortex, 24(9), 2489–2501.
Saygin, Z. M., Osher, D. E., Norton, E. S., Youssoufian, D.A., Beach, S. D., Feather, J., ... Kanwisher, N. (2016).Connectivity precedes function in the development of the visual word form area.Nature Neuroscience, 19(9),1250–1255.
Shim, G., Jung, W. H., Choi, J. S., Jung, M. H., Jang, J. H.,Park, J. Y., ... Kwon, J. S. (2009). Reduced cortical folding of the anterior cingulate cortex in obsessive–compulsive disorder.Journal of Psychiatry and Neuroscience, 34(6),443–449.
Shultz, S., Vouloumanos, A., Bennett, R. H., & Pelphrey, K.(2014). Neural specialization for speech in the first months of life.Developmental Science, 17(5), 766–774.
Skeide, M. A., Kraft, I., Müller, B., Schaadt, G., Neef, N. E.,Brauer, J., ... Friederici, A. D. (2016).NRSN1associated grey matter volume of the visual word form area reveals dyslexia before school.Brain, 139(10), 2792–2803.
Striedter, G. F., Srinivasan, S., & Monuki, E. S. (2015).Cortical folding: When, where, how, and why?.Annual Review of Neuroscience, 38, 291–307.
Striem-Amit, E., Hertz, U., & Amedi, A. (2011). Extensive cochleotopic mapping of human auditory cortical fields obtained with phase-encoding FMRI.PLoS One, 6(3),e17832.
Sun, T., & Hevner, R. F. (2014). Growth and folding of the mammalian cerebral cortex: From molecules to malformations.Nature Reviews Neuroscience, 15(4), 217–232.
Sun, T., Patoine, C., Abu-Khalil, A., Visvader, J., Sum, E.,Cherry, T. J., ... Walsh, C. A. (2005). Early asymmetry of gene transcription in embryonic human left and right cerebral cortex.Science, 308(5729), 1794–1798.
Sun, T., & Walsh, C. A. (2006). Molecular approaches to brain asymmetry and handedness.Nature Reviews Neuroscience,7(8), 655–662.
Sun, Z. Y., Rivière, D., Poupon, F., Régis, J., & Mangin, J.-F.(2007). Automatic inference of sulcus patterns using 3D moment invariants. In N. Ayache, S. Ourselin, & A. Maeder(Eds.),Medical image computing and computer-assisted intervention – MICCAI 2007(Vol. 4791, pp. 515–522).Berlin, Heidelberg: Springer.
Takerkart, S., Auzias, G., Brun, L., & Coulon, O. (2017).Structural graph-based morphometry: A multiscale searchlight framework based on sulcal pits.Medical Image Analysis,35, 32–45.
Tallinen, T., Chung, J. Y., Rousseau, F., Girard, N., Lefèvre,J., & Mahadevan, L. (2016). On the growth and form of cortical convolutions.Nature Physics, 12(6), 588–593.
Trefler, A., Sadeghi, N., Thomas, A. G., Pierpaoli, C., Baker,C. I., & Thomas, C. (2016). Impact of time-of-day on brain morphometric measures derived from T1-weighted magnetic resonance imaging.NeuroImage, 133, 41–52.
Vandermosten, M., Boets, B., Wouters, J., & Ghesquière, P.(2012). A qualitative and quantitative review of diffusion tensor imaging studies in reading and dyslexia.Neuroscience and Biobehavioral Reviews, 36(6), 1532–1552.
Vandermosten, M., Vanderauwera, J., Theys, C., De Vos, A.,Vanvooren, S., Sunaert, S., ... Ghesquière, P. (2015). A DTI tractography study in pre-readers at risk for dyslexia.Developmental Cognitive Neuroscience, 14, 8–15.
van Essen, D. C. (1997). A tension-based theory of morphogenesis and compact wiring in the central nervous system.Nature, 385(6614), 313–318.
Wallace, G. L., Robustelli, B., Dankner, N., Kenworthy, L.,Giedd, J. N., & Martin, A. (2013). Increased gyrification,but comparable surface area in adolescents with autism spectrum disorders.Brain, 136(6), 1956–1967.
Wang, Y. Y., Mauer, M. V., Raney, T., Peysakhovich, B.,Becker, B. L. C., Sliva, D. D., & Gaab, N. (2017).Development of tract-specific white matter pathways during early reading development in at-risk children and typical controls.Cerebral Cortex, 27(4), 2469–2485.
Weiner, K. S., Barnett, M. A., Lorenz, S., Caspers, J.,Stigliani, A., Amunts, K., ... Grill-Spector, K. (2017). The cytoarchitecture of domain-specific regions in human high-level visual cortex.Cerebral Cortex, 27(1), 146–161.Williams, V. J., Juranek, J., Cirino, P., & Fletcher, J. M.(2017). Cortical thickness and local gyrification in children with developmental dyslexia.Cerebral Cortex,doi: 10.1093/cercor/bhx001
Xia, Z. C., Hoeft, F., Zhang, L. J., & Shu, H. (2016).Neuroanatomical anomalies of dyslexia: Disambiguating the effects of disorder, performance, and maturation.Neuropsychologia, 81, 68–78.
Xu, G., Knutsen, A. K., Dikranian, K., Kroenke, C. D., Bayly,P. V., & Taber, L. A. (2010). Axons pull on the brain, but tension does not drive cortical folding.Journal of Biomechanical Engineering, 132(7), 071013.
Zilles, K., Armstrong, E., Schleicher, A., & Kretschmann,H.-J. (1988). The human pattern of gyrification in the cerebral cortex.Anatomy and Embryology, 179(2), 173–179.
Zilles, K., Palomero-Gallagher, N., & Amunts, K. (2013).Development of cortical folding during evolution and ontogeny.Trends in Neurosciences, 36(5), 275–284.
猜你喜歡
少兒美術·書法版(2021年11期)2021-10-20 06:23:28
少兒美術·書法版(2021年8期)2021-10-20 06:08:10
文苑(2020年4期)2020-05-30 12:35:30
小學生作文(中高年級適用)(2018年3期)2018-04-18 01:24:47
瘋狂英語·新策略(2017年8期)2017-05-31 08:13:46
華北電力大學學報(社會科學版)(2016年4期)2016-12-01 03:59:30
雜文選刊(2016年7期)2016-08-02 08:39:56
小天使·一年級語數英綜合(2016年6期)2016-05-14 12:21:05
少兒科學周刊·少年版(2015年4期)2015-07-07 21:11:17
語文知識(2014年10期)2014-02-28 22:00:56
