谷子PT2基因保守序列的克隆與分析
2018-06-19 11:49:28楊致榮高建華
山西農(nóng)業(yè)科學(xué) 2018年6期
關(guān)鍵詞:分析
李 燕,楊致榮,高建華
(1.山西農(nóng)業(yè)大學(xué)文理學(xué)院,山西 太谷 030801;2.山西農(nóng)業(yè)大學(xué)生命科學(xué)學(xué)院,山西 太谷 030801)
谷子(Setaria italica)又稱粟,是起源于我國的一種古老作物,也是我國過去幾千年來的主栽作物和中華民族的哺育作物。近年來,隨著人們生活水平的不斷提高及食品多樣化和營養(yǎng)化,小米及其制品備受青睞。提高谷子產(chǎn)量和品質(zhì)也成為目前研究的熱點。
對于水稻(Oryza sativa)等糧食作物而言,覆蓋地膜[1],噴施赤霉素[2],不同氮肥組合和施肥方式[3],提高籽粒大小、質(zhì)量,防止成熟后落粒,均是促進產(chǎn)量的有效手段。目前,已經(jīng)克隆出了多個籽粒性狀相關(guān)基因如 GL7[4],GW8[5],GW7[6]和 GW2[7]等;落粒相關(guān)基因,如 qSH1[8],SHAT1[9],SH4[10],PT2[11]和 SH5[12]等。其中,水稻PT2(Panicle Traits 2)基因編碼蛋白,即生長調(diào)節(jié)因子Os GRF4(Growth Regulating Factor 4,Os02g47280)是擬南芥At GRF的同源蛋白。研究表明,Os GRF4能夠調(diào)節(jié)細胞分裂素脫氫酶前體基因(CKX5和CKX1),導(dǎo)致細胞分裂素含量提高,進而調(diào)節(jié)籽粒大小、形狀等。當(dāng)Os GRF4表達受到抑制后,離層發(fā)育良好,從而增強了落粒性;反之,若Os GRF4高效表達,離層發(fā)育不完全,落粒性減弱[13]。PT2基因的表達受到miRNA的調(diào)控,第3個外顯子中有一段miR396結(jié)合序列。當(dāng)OsmiR396結(jié)合該位點時,PT2基因的表達被抑制;該結(jié)合位點的突變,導(dǎo)致OsmiR396無法結(jié)合,從而使PT2基因的表達量升高[14-15]。另外,作為一種調(diào)節(jié)因子,Os GRF4蛋白具有與核酸和其他蛋白質(zhì)(比如,GRF-Interacting Factors,GIF)互作的結(jié)構(gòu)域——WRC和QLQ。其中,WRC幫助Os GRF4蛋白與核酸互作,QLQ協(xié)助Os GRF4與其他蛋白互作[14-15]。另外,水稻Os04g51190基因編碼序列與Os GRF4氨基酸序列同源性約70%,但目前未發(fā)現(xiàn)其與籽粒性狀或落粒相關(guān)。
在豫谷1號的基因組數(shù)據(jù)(Setaria italica v2.2)中,有2個PT2基因的同源基因(Seita.1G287100.1和Seita.7G224500.1)。初步分析發(fā)現(xiàn),這2個基因均含有相應(yīng)的miR396結(jié)合序列,且其編碼多肽也具有WRC和QLQ結(jié)構(gòu)域。狗尾草(Setaria viridis)是谷子的近緣野生種,但二者在籽粒大小和落粒性等方面存在顯著差異。
為了研究PT2基因與谷子籽粒性狀及其落粒性的相關(guān)性,本研究首先分析了幾種植物PT2基因的親緣關(guān)系;預(yù)測了豫谷1號PT2基因啟動子序列中的相關(guān)元件,并驗證了PT2基因在豫谷1號不同部位的表達水平;篩選并比較了43種谷子和3種狗尾草中該基因的保守區(qū)域序列,旨在為后期篩選谷子落粒相關(guān)基因奠定了基礎(chǔ)。
1 材料和方法
1.1 材料
本研究所用到的谷子和狗尾草材料列于表1。所有材料均種植于山西農(nóng)業(yè)大學(xué)試驗田。
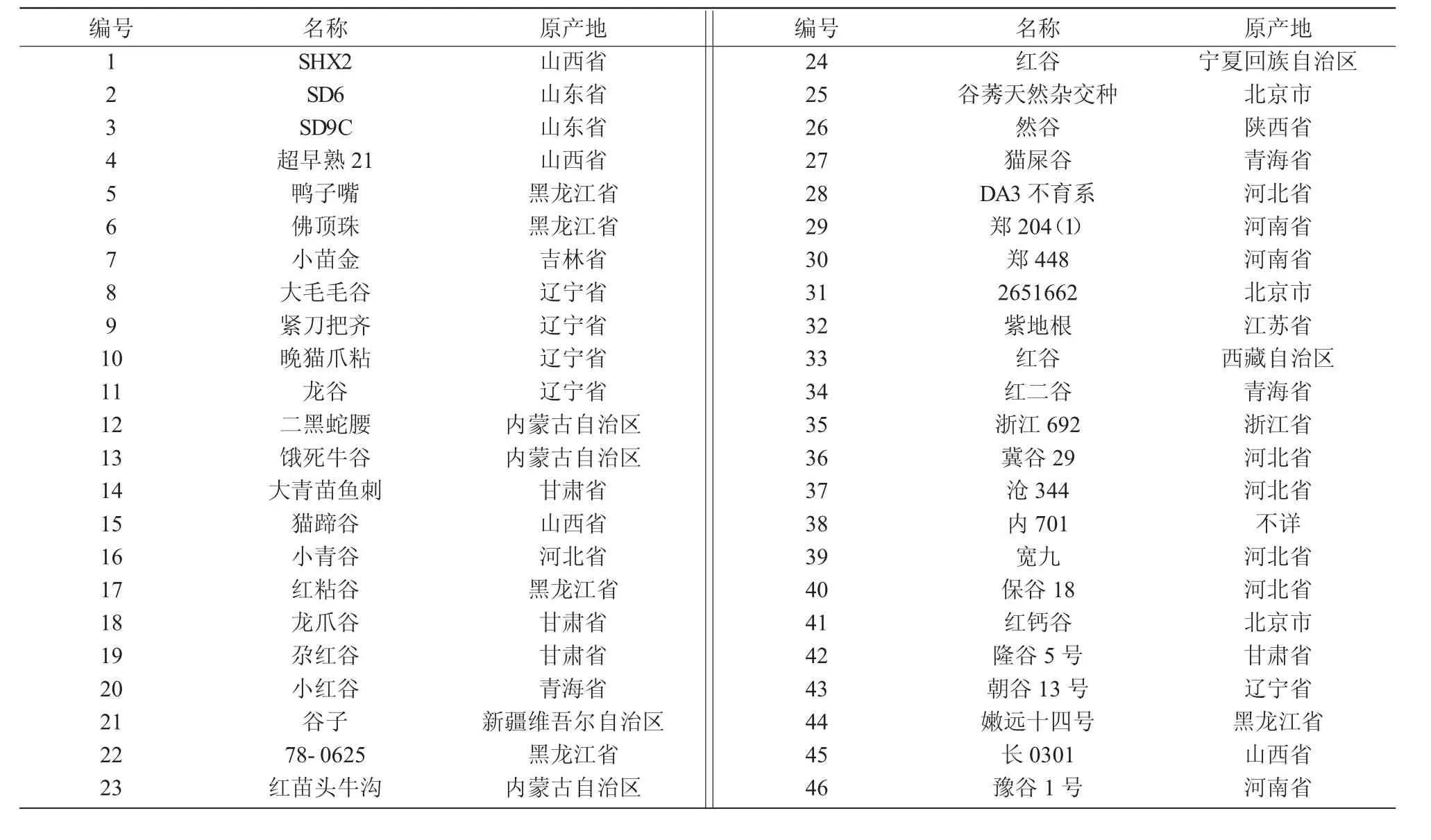
表1 研究所用到的谷子和狗尾草種質(zhì)資源
1.2 方法
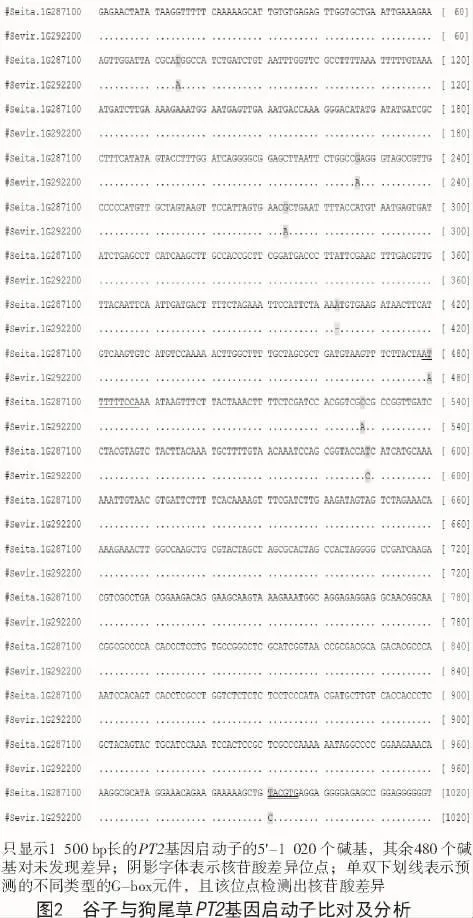
1.2.1 PT2基因及其啟動子的生物信息學(xué)分析從 phytozome網(wǎng)站(https://phytozome.jgi.doe.gov/)下載水稻PT2基因序列(LOC_Os02g47280),并下載擬南芥、水稻、谷子、狗尾草、高粱和玉米中與之同源性最高的基因序列。對這些基因編碼的氨基酸序列進行同源性分析,并利用MEGA 7.0分析其親緣關(guān)系。下載Seita.1G287100基因起始密碼子上游1 500 bp的序列,在 PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)進行啟動子在線預(yù)測。然后,對谷子和狗尾草PT2基因的啟動子進行序列比對。
1.2.2 總RNA提取與PT2基因的檢測 使用TaKaRa總RNA提取試劑盒(MiniBEST Plant RNA Extraction Kit,貨號 9769S),按照說明書提取豫谷1號苗和穗的總RNA。然后使用PrimeScript RT reagent Kit with gDNA Eraser試劑盒(TaKaRa,貨號RR047A)合成 cDNA。使用引物 579F(5′-CAGCGG CTTCCAGAACCACT)和 1179R(5′-GTTCGGCGAC TGAGAGCTTGCCA)檢測Seita.1G287100.1基因的表達。
1.2.3 植物基因組DNA的提取 谷子和狗尾草基因組DNA提取使用CTAB法:CTAB提取液(200 mmol/L Tris-Cl,250 mmol/L NaCl,25 mmol/L EDTA和0.5%SDS,pH值7.5)65℃預(yù)熱;在1.5 mL離心管中加入50mg幼嫩葉片,并加入500μLCTAB提取液,用高通量DNA研磨機(寧波新芝生物科技股份有限公司,型號SCIENTZ-48)充分研磨;放入65℃水浴溫浴1 h,期間每隔5 min顛倒混勻;加入500 μL氯仿 /異戊醇(體積比 24∶1),振蕩混勻,室溫靜置10 min;12 000 r/min離心10 min,取上清至新離心管;加入等體積氯仿/異戊醇(體積比24∶1),混勻后12 000 r/min離心10 min;取上清至新離心管,并加入0.7倍體積預(yù)冷的異丙醇,混勻后放入-20℃1 h;取出后12 000 r/min離心10 min,棄上清,并用70%乙醇清洗沉淀2次;室溫晾干,加入30 μL ddH2O溶解DNA,最后-20℃保存?zhèn)溆谩?/p>
1.2.4 PT2基因外顯子的擴增及序列分析 根據(jù)數(shù)據(jù)庫中Seita.1G287100基因序列,設(shè)計2對特異性引物,用于分別擴增不同谷子或狗尾草中PT2基因的前3個外顯子序列。引物PT21-E12-F(5′-ACAAAGCGGGCAATAAAGGC)和PT21-E12-R(5′-ACACGGCAAGTATTCGGGAG)用于擴增外顯子1和 2。引物 PT21-E3-F(5′-GTAGCTGCTCCAC TGTTCGC)和 PT21-E3-R(5′-AGTTATCTGCGTGA CCATCTCTG)用于擴增外顯子3。PCR體系為:Takara Ex Taq premix (RR902A)10 μL,DNA 模板0.5μL,上下游引物各 0.5 μL,ddH2O補齊至 20 μL。PCR反應(yīng)條件為:94℃預(yù)變性3 min;94℃,30 s,60 ℃,30 s,72℃,60 s,共 35 個循環(huán);最后 72 ℃延伸5 min。PCR產(chǎn)物使用1%的瓊脂糖凝膠電泳檢測,并克隆測序。使用MEGA 7.0對不同谷子和狗尾草的PT2基因序列進行同源性和單核苷酸多態(tài)性分析。
2 結(jié)果與分析
2.1 谷子PT2候選基因的篩選與分析

將數(shù)據(jù)庫中谷子、狗尾草以及其他植物的PT2候選基因編碼的蛋白質(zhì)進行比對,并構(gòu)建系統(tǒng)發(fā)育樹。分析發(fā)現(xiàn),已經(jīng)報道的水稻Os GRF4蛋白(Os02g47280基因編碼)[10-12]與谷子 Seita.1G287100、狗尾草Sevir.1G292200編碼的蛋白質(zhì)序列同源性較高(圖1-A)。其中,谷子Seita.1G287100和狗尾草Sevir.1G292200編碼的蛋白序列完全一致。該結(jié)果說明這2個基因分別是谷子和狗尾草的PT2基因。進一步分析3種植物的PT2蛋白質(zhì)序列發(fā)現(xiàn),QLQ結(jié)構(gòu)域序列完全一致,而WRC結(jié)構(gòu)域也僅存在一個氨基酸差異(圖1-B)。該結(jié)果說明該生長調(diào)節(jié)因子中參與核酸或蛋白互作的2個重要位點高度保守,保障其正常執(zhí)行功能。
2.2 谷子PT2基因啟動子元件分析
將豫谷1號的PT2基因(Seita.1G287100)上游1 500 bp的序列進行在線分析,結(jié)果表明,該基因啟動子較為復(fù)雜。除了常見的TATA-box,CAAT-box等核心啟動子元件外,還有較多的響應(yīng)元件,比如,光 應(yīng) 答 響 應(yīng) 元 件 ACE,AE-box,BOXI,G-BOX,G-box,GT1-motif,I-box,L-box,sp1,TCT-motif;脫落酸信號轉(zhuǎn)導(dǎo)下游響應(yīng)元件ABRE和motif-Iib;茉莉酸甲酯 (MeJA) 響應(yīng)元件 CGTCA-motif和TGACG-motif;赤霉素響應(yīng)元件 GARE-motif;缺氧特異性誘導(dǎo)響應(yīng)元件GC-motif;參與防御和應(yīng)激反應(yīng)響應(yīng)元件TC-rich-repeats等(表2)。另外,還有多種其他調(diào)控位點,比如調(diào)控轉(zhuǎn)錄水平的5′UTR Py-rich stretch,MYBHv1 結(jié)合位點 CCAAT-box,胚乳表達所需skn-1 motif元件,玉米醇溶蛋白代謝調(diào)控響應(yīng)元件O2-site,參與分生組織表達響應(yīng)元件CAT-box等。該預(yù)測結(jié)果說明PT2基因參與多種環(huán)境響應(yīng)及代謝調(diào)節(jié),是重要的調(diào)節(jié)因子。另外,對谷子和狗尾草PT2基因的啟動子序列進行對比發(fā)現(xiàn),二者只有8個核苷酸位點有差異(圖2)。其中,只有2個位點分別位于2個G-box元件中。該結(jié)果暗示在2種植物中,PT2基因的調(diào)控基本一致。
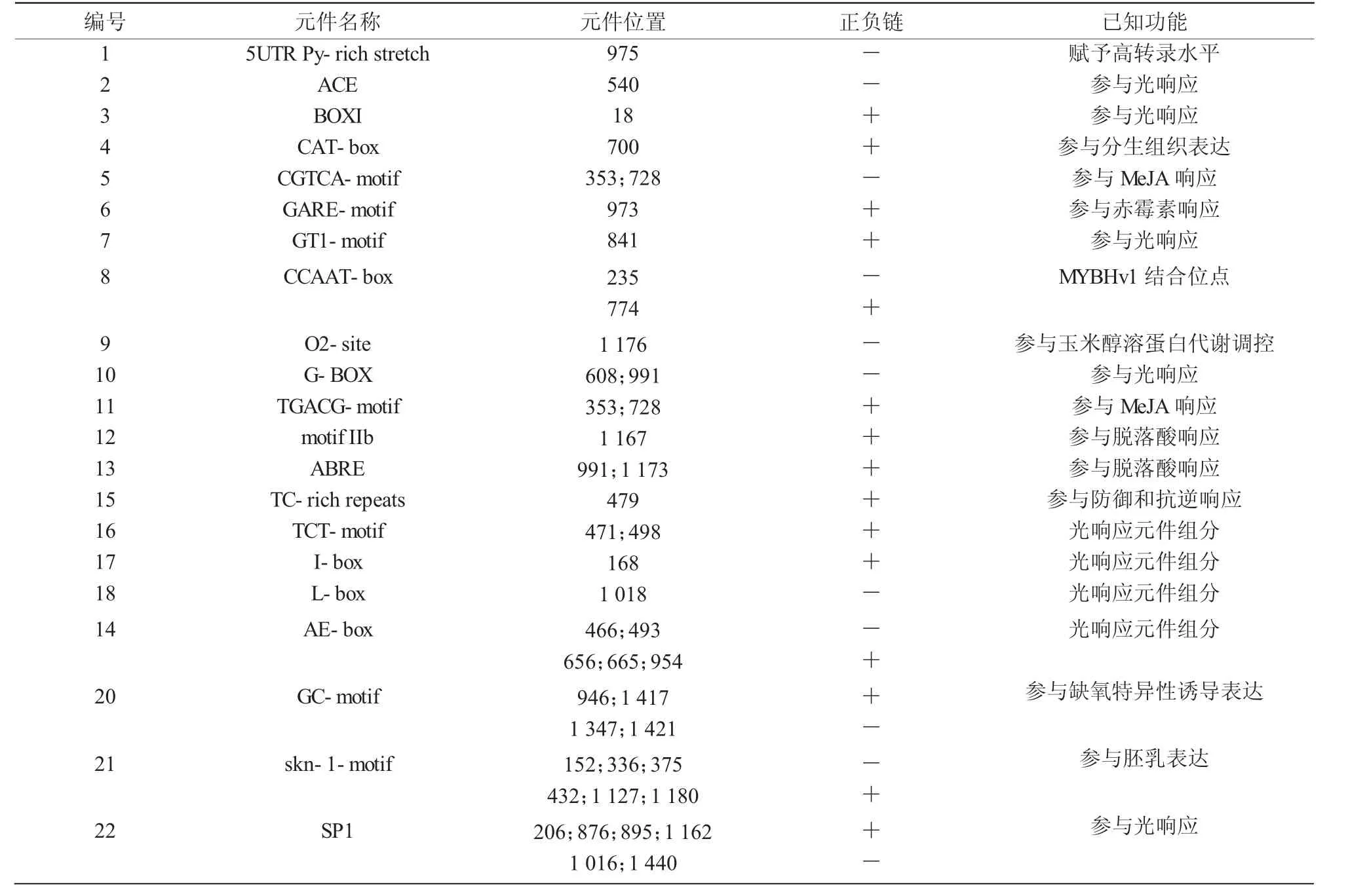
表2 PT2啟動子元件分析
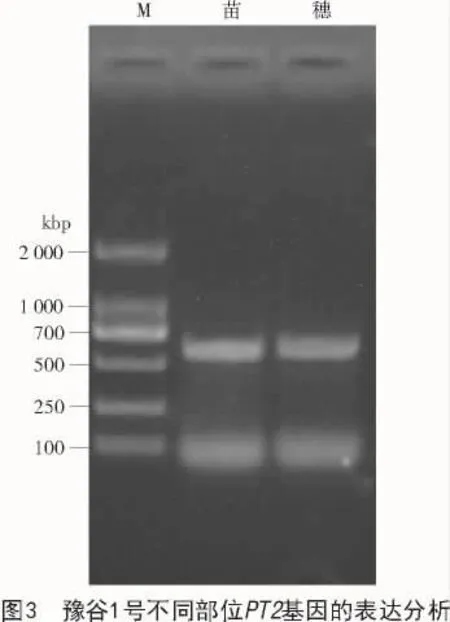
2.3 谷子PT2家族基因表達部位分析
提取豫谷1號苗和穗的總RNA,并將其反轉(zhuǎn)錄為cDNA。以cDNA為模板,使用引物579F和1179R,檢測該基因在不同部位的轉(zhuǎn)錄情況,結(jié)果表明,該基因在豫谷1號苗、穗部位均有表達(圖3)。說明PT2基因可能參與多種代謝調(diào)節(jié)和環(huán)境響應(yīng)。
2.4 PT2基因保守性分析
為分析PT2基因在不同品系中的保守性,將43種谷子和3種狗尾草的PT2基因前3個外顯子(包括OsmiR396結(jié)合位點、WRC和QLQ的編碼區(qū)域)進行了序列測定。
序列比對發(fā)現(xiàn),其核酸序列高度保守,共發(fā)現(xiàn)4處SNP(圖4),但是只有一處導(dǎo)致氨基酸突變(T190A)。該突變位點(T190A)在谷子和狗尾草中均被檢測到,說明其與谷子和狗尾草之間籽粒性狀、落粒性的差異無關(guān)。

3 討論
對于水稻、小麥、谷子等禾本科作物而言,其籽粒大小、形狀、數(shù)量以及落粒等性狀是決定其產(chǎn)量高低的重要因素。研究表明,水稻PT2基因編碼的Os GRF4蛋白能夠調(diào)控籽粒形狀、大小以及落粒性等多種性狀。為研究該基因在谷子籽粒性狀調(diào)控中的作用,本試驗通過生物信息學(xué)的方法,確定了谷子和狗尾草中的PT2候選基因。通過對谷子PT2基因啟動子中各種相應(yīng)元件的分析發(fā)現(xiàn),該基因的啟動子含有多種環(huán)境(如光照、干旱)和激素(如水楊酸、茉莉酸)等響應(yīng)元件,暗示其參與多種代謝調(diào)節(jié)。RNA轉(zhuǎn)錄檢測結(jié)果也顯示,其在植株不同部位均有表達,進一步確定該基因具有多重功能。但是,本研究并未發(fā)現(xiàn)該基因其他形式的轉(zhuǎn)錄本。
值得注意的是,谷子與狗尾草的PT2基因編碼蛋白序列完全一致,僅在個別品系中發(fā)現(xiàn)一個氨基酸的點突變。該點突變在谷子和狗尾草中均存在。該基因啟動子序列也基本吻合,僅存在幾個核苷酸的差異。其中,2個差異分別位于2個G-box元件中。這2個差異是否影響該基因?qū)δ承┮蛩氐捻憫?yīng),尚未確定。然而,谷子和狗尾草籽粒性狀差異顯著,比如籽粒形狀明顯不同,落粒性完全相反。說明在2種植物體內(nèi),其他未知的基因調(diào)控了上述差異,其與水稻不同。
本研究通過生物信息學(xué)的方法確定了谷子和狗尾草的PT2基因,并且通過二者間的比對,初步判斷PT2基因與谷子籽粒大小、形狀以及落粒性等性狀無關(guān),結(jié)果為后續(xù)篩選相關(guān)基因奠定了堅實的基礎(chǔ)。
[1]世佳,任曉利,魏志敏.覆膜對夏播谷子生長發(fā)育與產(chǎn)量的影響機制及其相關(guān)性分析[J].華北農(nóng)學(xué)報,2016,31(Z):25-30.
[2]鄭林林,原向陽,邵冬紅.灌漿期噴施赤霉素對谷子農(nóng)藝性狀及產(chǎn)量的影[J].山西農(nóng)業(yè)科學(xué),2014,42(5):455-457.
[3]代小冬,常世豪,楊育峰.不同氮肥組合和施肥方式對谷子生長和產(chǎn)量的影響[J].河南農(nóng)業(yè)科學(xué),2014,43(11):13-16.
[4]WANG Y,XIONG G,HU J,et al.Copy number variation at the GL7 locus contributes to grain size diversity in rice[J].Nature Genetics,2015,47(8):944-948.
[5] WANG S,WU K,YUAN Q,et al.Control of grain size,shape and quality by OsSPL16 in rice[J].Nature Genetics,2012,44(8):950-954.
[6]WANG S,LI S,LIU Q,et al.The OsSPL16-GW7 regulatory module determines grain shape and simultaneously improves rice yield and grain quality[J].Nature Genetics,2015,47(8):949-954.
[7]BEDNAREK J,BOULAFLOUS A,GIROUSSE C,et al.Down-regulation of the TaGW2 gene by RNA interference results in decreased grain size and weight in wheat[J].Journal of Experimental Botany,2012,63(16):5945-5955.
[8] MAGWA R A,ZHAO H,YAO W,et al.Genomewide association analysis for awn length linked to the seed shattering gene qSH1 in rice[J].Journal of Genetics,2016,95(3):639-646.
[9]ZHOUY,LUD,LI C,et al.Genetic control of seed shatteringin rice by the APETALA2 transcription factor shattering abortion1[J].The Plant Cell,2012,24(3):1034-1048.
[10] INOUE C,HTUN TM,INOUE K,et al.Inhibition of abscission layer formation by an interaction of two seed-shattering loci,sh4 and qSH3,in rice[J].Genes&Genetic Systems,2015,90(1):1-9.
[11] HU J,WANG Y,F(xiàn)ANG Y,et al.A rare allele of GS2 enhances grain size and grain yield in rice[J].Molecular Plant,2015,8(10):1455-1465.
[12]YOON J,CHO LH,KIM SL,et al.The BEL1-type homeobox gene SH5 induces seed shattering by enhancing abscission-zone development and inhibiting lignin biosynthesis[J].The Plant Journal,2014,79(5):717-728.
[13] SUN P,ZHANG W,WANG Y,et al.OsGRF4 controls grain shape,panicle length and seed shattering in rice[J].Journal of Integrative Plant Biology,2016,58(10):836-847.
[14] LI S,GAO F,XIE K,et al.The OsmiR396c-OsGRF4-OsGIF1 regulatory module determines grain size and yield in rice[J].Plant Biotechnology Journal,2016,14(11):2134-2146.
[15] DUAN P,NI S,WANG J,et al.Regulation of OsGRF4 by OsmiR396 controls grain size and yield in rice[J].Nature Plant,2015,2(1):15203.
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設(shè)計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當(dāng)代經(jīng)濟研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫(yī)藥現(xiàn)代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學(xué)學(xué)報(社會科學(xué)版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
