尾狀核?眶部?jī)?nèi)側(cè)前額葉的功能連接與反應(yīng)性攻擊的關(guān)系:基于靜息態(tài)功能磁共振研究
2018-06-22 01:49:14侯璐璐李長(zhǎng)燃王煥貞
心理學(xué)報(bào) 2018年6期
江 琦 侯璐璐,2 邱 江 李長(zhǎng)燃 王煥貞
(1西南大學(xué)心理學(xué)部, 心理健康教育中心, 重慶 400715) (2南京大學(xué)社會(huì)學(xué)院心理學(xué)系, 南京 210023)(3西南大學(xué)心理學(xué)部, 認(rèn)知與人格教育部重點(diǎn)實(shí)驗(yàn)室, 重慶 400715)
1 引言
反應(yīng)性攻擊(reactive aggression)是指在被激惹后產(chǎn)生的沖動(dòng)的、不假思索的指向另一個(gè)人并對(duì)其造成傷害的行為(Anderson & Bushman, 2002)。由于反應(yīng)性攻擊會(huì)造成很大的社會(huì)危害, 因而受到社會(huì)學(xué)、心理學(xué)等多學(xué)科的廣泛關(guān)注(Herpertz et al.,2017; McEwen & McEwen, 2017; Fite, Rubens,Preddy, Raine, & Pardini, 2014)。近年來(lái), 研究者試圖采用事件相關(guān)電位、腦成像等方法對(duì)反應(yīng)性攻擊的神經(jīng)機(jī)制進(jìn)行探討。已有對(duì)具有較高攻擊性的病人以及正常人進(jìn)行的腦成像研究表明, 可能存在一個(gè)網(wǎng)絡(luò)與反應(yīng)性攻擊相關(guān), 這個(gè)網(wǎng)絡(luò)包括杏仁核(Amygdala; McCloskey et al., 2016)、尾狀核(Glenn& Yang, 2012)和 OMPFC (Beyer, Münte, G?ttlich, &Kr?mer, 2015)。研究者進(jìn)一步指出降低的前額葉皮層的活動(dòng)與升高的皮層下腦區(qū)的活動(dòng)(如杏仁核、尾狀核) 可能共同導(dǎo)致了反應(yīng)性攻擊的發(fā)生(Rosell& Siever, 2015; Siever, 2008; Nelson & Trainor,2007), 也就是說(shuō)反應(yīng)性攻擊的產(chǎn)生可能源于前額葉皮層與皮層下腦區(qū)的功能連接失調(diào)。
已有研究或者根據(jù)相關(guān)研究(如 Coccaro,McCloskey, Fitzgerald, & Phan, 2007), 或者通過(guò)對(duì)比攻擊性較強(qiáng)的病人、罪犯與正常人群的大腦活動(dòng)差異(如Motzkin, Newman, Kiehl, & Koenigs, 2011)來(lái)推論反應(yīng)性攻擊的神經(jīng)機(jī)制, 這些探索雖然為揭示反應(yīng)性攻擊的神經(jīng)機(jī)制起到了重要的啟示作用, 但是尚不能提供直接的實(shí)證證據(jù)加以證實(shí)。究其原因, 在于反應(yīng)性攻擊的發(fā)生涉及多個(gè)心理過(guò)程(Bettencourt, Talley, Benjamin, & Valentine, 2006;Ramirez & Andreu, 2006), 這無(wú)疑為揭示其神經(jīng)機(jī)制造成了很大的困難。因此, 近年來(lái)一些研究者開(kāi)始嘗試運(yùn)用經(jīng)典的攻擊范式及其變式來(lái)探討反應(yīng)性攻擊的神經(jīng)機(jī)制, 其中, Taylor攻擊范式(Taylor Aggression Paradigm, TAP; Taylor, 1967)是應(yīng)用較為廣泛的一種經(jīng)典的攻擊范式(Riva et al., 2017;Liu, Teng, Lan, Zhang, & Yao, 2015)。在經(jīng)典的Taylor攻擊范式中, 主試安排被試與另一個(gè)(虛假)對(duì)手進(jìn)行反應(yīng)時(shí)任務(wù)的競(jìng)賽, 每次競(jìng)爭(zhēng)中速度快的一方可以為速度慢的一方施加電擊(或噪音), 然后根據(jù)施加的電壓或者噪音強(qiáng)度來(lái)判斷被試的反應(yīng)性攻擊。已有研究表明, 該范式具有良好的效度(Giancola& Parrott, 2008)。2007 年, Kr?mer, Riba, Richter和Münte進(jìn)而把Taylor攻擊范式中的虛假對(duì)手增加為兩個(gè), 分別對(duì)應(yīng)激發(fā)與非激發(fā)條件, 從而可以有效分析激發(fā)條件對(duì)被試反應(yīng)性攻擊的影響。但是, 綜合分析近年來(lái)針對(duì)正常人群的利用 Taylor范式及其變式對(duì)反應(yīng)性攻擊的神經(jīng)機(jī)制進(jìn)行的研究(Beyer,Münte, Erdmann, & Kr?mer, 2014; Beyer et al., 2015;Kr?mer, Rib, Richter, & Münte, 2011; Kr?mer, Jansma,Tempelmann, & Münte, 2007; Lotze, Veit, Anders, &Birbaumer, 2007)可以發(fā)現(xiàn), 這類研究的實(shí)驗(yàn)設(shè)置都各有不同, 甚至同一個(gè)課題組基于研究目的不同也對(duì)同一范式做了不同程度的修改, 因此得到的結(jié)論各不相同, 并且難以直接進(jìn)行比較。那么, 是否可以找到一種穩(wěn)定的指標(biāo), 通過(guò)找到其與反應(yīng)性攻擊的關(guān)系, 進(jìn)而探討反應(yīng)性攻擊的神經(jīng)機(jī)制呢?已經(jīng)較為成熟的靜息態(tài)功能磁共振技術(shù)為我們提供了新的思路。
靜息態(tài)功能磁共振具有不受實(shí)驗(yàn)任務(wù)的限制和制約、能夠可靠地測(cè)量到大腦內(nèi)在的自發(fā)活動(dòng)等優(yōu)勢(shì)(Zuo et al., 2010), 被廣泛應(yīng)用于健康被試與病人的研究中, 以用于發(fā)現(xiàn)個(gè)體在靜息狀態(tài)下大腦活動(dòng)情況與某些疾病、特質(zhì)以及行為的關(guān)系(Fulwiler,King, & Zhang, 2012; Wu et al., 2011; Greicius et al.,2007)。功能連接的方法普遍被用來(lái)探索腦區(qū)之間的聯(lián)系以及這種聯(lián)系與某種特定的心理和行為之間的關(guān)系(Takeuchi et al., 2012; Zeng et al., 2012;Hahn et al., 2011)。近年來(lái), 該方法在攻擊性相關(guān)領(lǐng)域也獲得了一些有意義的結(jié)果, 例如 Hoptman等(2010)的研究表明, 比起正常個(gè)體, 患有精神分裂癥的病人的杏仁核與腹側(cè)前額葉的功能連接降低,并且功能連接的強(qiáng)度與攻擊性呈負(fù)相關(guān)關(guān)系。
然而, 上述研究均是采用相關(guān)分析的方法考察腦區(qū)之間的協(xié)同工作, 還不能反映腦區(qū)之間信息的輸入輸出方向等關(guān)系。因此, 有研究者進(jìn)一步認(rèn)為廣義的功能連接應(yīng)該分為(狹義的)功能連接(Functional Connectivity, FC)與效應(yīng)連接(Effective Connectivity,EC)。關(guān)于效應(yīng)連接的分析中, 格蘭杰因果分析(Granger Causality Analysis, GCA)受到研究者們的廣泛關(guān)注和青睞。它主要是利用多重線性回歸的方法考察某個(gè)腦區(qū)在過(guò)去某一時(shí)間點(diǎn)的活動(dòng)是否可以反映現(xiàn)在另外一個(gè)腦區(qū)的活動(dòng)情況, 可以揭示腦區(qū)之間單向的因果關(guān)系(Hamilton, Chen, Thomason,Schwartz, & Gotlib, 2011; Chen, Hamilton, Thomason,Gotlib, & Saad, 2009)。
Coccaro, Sripada, Yanowitch 和 Phan (2011)基于以往研究指出, OMPFC與反應(yīng)性攻擊密切相關(guān)。OMPFC能對(duì)攻擊沖動(dòng)進(jìn)行有效調(diào)控, Pietrini,Guazzelli, Basso, Jaffe和Grafman (2000)給被試呈現(xiàn)一系列的場(chǎng)景要求被試抑制或者表達(dá)對(duì)侵犯者的攻擊, 結(jié)果顯示當(dāng)被試抑制攻擊沖動(dòng)時(shí), OMPFC激活程度增強(qiáng)。而OMPFC損傷的病人不能對(duì)敵意反應(yīng)進(jìn)行調(diào)節(jié)(Koenigs & Tranel, 2007), 脾氣變得暴躁(Damasio, Grabowski, Frank, Galaburda, & Damasio,1994)。OMPFC還與社會(huì)情緒信息加工相關(guān), Beyer等(2015)的研究通過(guò)在TAP范式的決策階段之前增加面孔(憤怒或者中性), 來(lái)考察憤怒在反應(yīng)性攻擊中的作用, 結(jié)果顯示呈現(xiàn)憤怒面孔時(shí), OMPFC的激活程度與反應(yīng)性攻擊負(fù)相關(guān)顯著, 這表明 OMPFC對(duì)社會(huì)情緒信息(即, 情緒面孔)的反應(yīng)可以預(yù)測(cè)其隨后的反應(yīng)性攻擊行為。New等(2007)在比較有邊緣人格障礙的個(gè)體與正常個(gè)體的大腦活動(dòng)情況時(shí)發(fā)現(xiàn), 比起那些有邊緣性人格障礙的個(gè)體, 正常個(gè)體的OMPFC與杏仁核之間的“耦合”更強(qiáng)。此外, 比起正常的個(gè)體, 當(dāng)看到憤怒的面孔時(shí), 患有間歇性爆發(fā)障礙的個(gè)體(攻擊性較強(qiáng))OMPFC與杏仁核之間的功能連接下降(Coccaro et al., 2007)。da Cunha-Bang等(2017)研究顯示, 比起正常人, 暴力罪犯在受到敵意激發(fā)后, 反應(yīng)性攻擊更強(qiáng), 尾狀核和杏仁核的激活強(qiáng)度升高, 并且尾狀核?前額葉以及杏仁核?前額葉的功能連接降低。以上研究都顯示OMPFC與反應(yīng)性攻擊密切相關(guān), 可以對(duì)敵意反應(yīng)以及攻擊沖動(dòng)進(jìn)行抑制, 并且和杏仁核、尾狀核等皮層下腦區(qū)的連接也在反應(yīng)性攻擊中起著重要作用。另外, TAP范式由于虛假對(duì)手的設(shè)置使得被試在實(shí)驗(yàn)過(guò)程中處于一種“競(jìng)爭(zhēng)和人際對(duì)抗”的背景下(Giancola & Parrott, 2008), 具有較強(qiáng)的社會(huì)互動(dòng)性, 而OMPFC在社會(huì)情緒信息加工、情緒性決策方面, 尤其是社會(huì)互動(dòng)背景下發(fā)揮重要作用(Rudebeck,Bannerman, & Rushworth, 2008), 因此OMPFC可以作為本研究的種子點(diǎn)。
綜上所述, 本研究擬選取OMPFC作為種子點(diǎn),首先運(yùn)用靜息態(tài)功能連接方法探索眶部?jī)?nèi)側(cè)前額葉?皮層下腦區(qū)的功能連接及其與反應(yīng)性攻擊的關(guān)系; 其次, 進(jìn)一步運(yùn)用格蘭杰因果模型分析探討靜息狀態(tài)下眶部?jī)?nèi)側(cè)前額葉?皮層下腦區(qū)的效應(yīng)連接及其與反應(yīng)性攻擊的關(guān)系。基于以往的研究結(jié)果(da Cunha-Bang et al., 2017; Coccaro et al., 2007; New et al., 2007), 我們假設(shè)OMPFC與皮層下腦區(qū)(例如,杏仁核、尾狀核)的連接與反應(yīng)性攻擊負(fù)相關(guān), 即當(dāng)OMPFC與皮層下腦區(qū)的連接較強(qiáng)時(shí), 個(gè)體反應(yīng)性攻擊較低; 反之, 當(dāng) OMPFC與皮層下腦區(qū)的連接較弱時(shí), 個(gè)體反應(yīng)性攻擊較強(qiáng)。在理論層面, 本研究利用靜息態(tài)功能磁共振技術(shù)的特點(diǎn)避開(kāi)了任務(wù)態(tài)功能磁共振研究因受任務(wù)設(shè)置不同而導(dǎo)致的結(jié)論分歧的不足, 直接為前額葉皮層?皮層下腦區(qū)功能連接與反應(yīng)性攻擊的關(guān)系研究提供了新的實(shí)證證據(jù), 為全面揭示反應(yīng)性攻擊的神經(jīng)機(jī)制提供了重要的基線資料。在實(shí)踐層面, 本研究可以為針對(duì)高反應(yīng)性攻擊傾向個(gè)體在認(rèn)知神經(jīng)層面的預(yù)測(cè)和診斷提供理論指導(dǎo)。
2 方法
2.1 被試
本研究隨機(jī)選取了 43名健康右利手在校大學(xué)生, 其中男性 19名, 女性 23名, 年齡 18~22歲(20.05 ± 0.92)。所有被試視力或者校正視力正常,無(wú)色盲, 無(wú)精神疾病史和手術(shù)外傷史。本實(shí)驗(yàn)通過(guò)了西南大學(xué)腦成像中心倫理委員會(huì)批準(zhǔn), 與所有被試簽訂了知情同意書(shū), 并在實(shí)驗(yàn)結(jié)束后給予一定的報(bào)酬。
2.2 范式
本研究采取了被廣泛使用的 Taylor攻擊范式(Taylor Aggression Paradigm, TAP), 行為學(xué)數(shù)據(jù)采集實(shí)驗(yàn)采用E-prime 2.0進(jìn)行。圖1呈現(xiàn)了單個(gè)試次的流程, 首先在屏幕上出現(xiàn)一個(gè)時(shí)間為8 s的注視點(diǎn)“+”, 然后屏幕上會(huì)出現(xiàn)此次的競(jìng)爭(zhēng)對(duì)手(1號(hào)或者2號(hào)), 在此過(guò)程中, 被試需要通過(guò)按鍵做出為1~8的強(qiáng)度的噪音懲罰的選擇1Level 1為70 db,Level 8為105 db,每級(jí)相差5 db。在練習(xí)階段,被試已經(jīng)聽(tīng)了Level 2、Level 5和Level 8的噪音強(qiáng)度。。接下來(lái)是競(jìng)爭(zhēng)反應(yīng)時(shí)任務(wù), 即當(dāng)屏幕上出現(xiàn)一個(gè)白色圓圈的時(shí)候被試必須盡可能快的按鍵, 緊接著被試會(huì)收到此回合的輸贏的結(jié)果和對(duì)手為他選擇的懲罰強(qiáng)度。
需要注意的是, 在整個(gè)實(shí)驗(yàn)過(guò)程中并沒(méi)有真正的兩位對(duì)手與被試共同完成競(jìng)爭(zhēng)反應(yīng)時(shí)任務(wù), 所有的反饋都是電腦按如下參數(shù)進(jìn)行設(shè)置的結(jié)果:對(duì)手1總是做出強(qiáng)度為1~4的選擇, 平均為2.0, 為非激發(fā)條件; 對(duì)手 2總是做出 5~8的選擇, 平均為 6.5,為激發(fā)條件。為了使實(shí)驗(yàn)更具有真實(shí)性, 將實(shí)驗(yàn)分為3個(gè)run進(jìn)行, 每個(gè)run中包括20個(gè)trials, 被試成功的次數(shù)占一半。為了增加反饋結(jié)果的真實(shí)性,我們參考以往研究(Maren, 2001), 對(duì)反饋結(jié)果的出現(xiàn)做了如下設(shè)置:在第一個(gè) run, 被試與 1號(hào)對(duì)手的競(jìng)爭(zhēng)中勝利3次, 失敗7次, 被試與2號(hào)對(duì)手的競(jìng)爭(zhēng)中勝利7次, 失敗3次; 在第二個(gè)run, 被試與1號(hào)對(duì)手的競(jìng)爭(zhēng)中勝利 5次, 失敗 5次, 被試與 2號(hào)對(duì)手的競(jìng)爭(zhēng)中勝利5次, 失敗5次; 在第三個(gè)run,被試與1號(hào)對(duì)手的競(jìng)爭(zhēng)中勝利7次, 失敗3次, 被試與2號(hào)對(duì)手的競(jìng)爭(zhēng)中勝利3次, 失敗7次, 即隨著實(shí)驗(yàn)的進(jìn)行, 被試受到的懲罰越來(lái)越大。
2.3 實(shí)驗(yàn)程序
被試進(jìn)入實(shí)驗(yàn)室后, 首先簽署知情同意書(shū)。然后由主試介紹實(shí)驗(yàn)程序, 被試在掃描室外完成練習(xí)實(shí)驗(yàn), 確保被試了解指導(dǎo)語(yǔ)并且能正確做出反應(yīng),然后由掃描員將其送入掃描室, 依次進(jìn)行靜息態(tài)、任務(wù)態(tài)和結(jié)構(gòu)像的掃描2由于篇幅所限,本文沒(méi)有呈現(xiàn)任務(wù)態(tài)中的結(jié)果。。
2.4 數(shù)據(jù)采集
MRI數(shù)據(jù)采集使用西門子3.0T磁共振掃描儀(Siemens Medical, 德國(guó)), 被試的頭部使用MRI兼容的泡沫墊固定以減少頭動(dòng)。掃描前, 讓被試更換實(shí)驗(yàn)室專用服裝, 以避免衣服上的金屬物體對(duì)被試安全和成像質(zhì)量的影響, 同時(shí)取下被試佩戴的金屬首飾或者假牙等。在結(jié)構(gòu)像和靜息態(tài)掃描的過(guò)程中,都要求被試睜開(kāi)眼睛看屏幕上的注視點(diǎn), 保持頭部不要發(fā)生移動(dòng), 不要想特殊的事情。
T1加權(quán)結(jié)構(gòu)像使用磁化準(zhǔn)備快遞采集梯度回波序列(magnetization-prepared rapid acquisition gradient echo, MPRAGE)采集, 具體掃描參數(shù)為:重復(fù)時(shí)間(repetition time, TR) = 2600 ms, 回波時(shí)間(echo time, TE) = 3.02 ms, 反轉(zhuǎn)角(flip angle) = 8°,層厚(slice thickness) = 1.0 mm, 矩陣(matrix) = 256 ×256, 體素大小(voxel size) = 1 mm × 1 mm × 1 mm,掃描176層覆蓋全腦。

圖1 實(shí)驗(yàn)流程圖
靜息態(tài)數(shù)據(jù)使用 EPI (gradient-echo planar imaging, EPI)序列采集, 具體掃描參數(shù)為:重復(fù)時(shí)間(repetition time, TR) = 2000 ms, 回波時(shí)間(echo time, TE) = 30 ms, 反轉(zhuǎn)角(flip angle) = 90°, 層厚(slice thickness) = 3 mm, 層間距(slice gap) = 1 mm,矩陣(matrix) = 64 × 64, 體素大小(voxel size) =3.4 mm × 3.4 mm × 4 mm, 層數(shù)(slices) = 32, 掃描時(shí)間為8分鐘零4秒, 共獲得242個(gè)時(shí)間點(diǎn)的圖像。
2.5 數(shù)據(jù)預(yù)處理
靜息態(tài)數(shù)據(jù)使用 DPARSF軟件(http://www.restfmri.net/forum/DPARSF)進(jìn)行預(yù)處理(Yan & Zang,2010)。具體流程包括:第一步, 將原始DICOM數(shù)據(jù)轉(zhuǎn)換為 NIFTI數(shù)據(jù)格式; 第二步, 為了避免磁共振信號(hào)開(kāi)機(jī)時(shí)的不穩(wěn)定和被試剛剛進(jìn)入掃描儀的不適應(yīng)帶來(lái)的影響, 刪除前10個(gè)時(shí)間點(diǎn)的數(shù)據(jù), 最終剩余 232個(gè)時(shí)間點(diǎn)進(jìn)行后續(xù)處理; 第三步, 時(shí)間層校正(slice timing), 參考層為每個(gè)全腦掃描過(guò)程中位于中間時(shí)間點(diǎn)的那一層; 第四步, 頭動(dòng)校正(realign), 將平動(dòng)超過(guò)了 2 mm或者轉(zhuǎn)動(dòng)超過(guò)了 2°的被試進(jìn)行排除; 第五步, 去除協(xié)變量(nuisance covariates regression), 包括6個(gè)頭動(dòng)參數(shù)和白質(zhì)信號(hào)、腦脊液信號(hào); 第六步, 采用DARTEL進(jìn)行空間標(biāo)準(zhǔn)化(spatial normalization), 將圖像配準(zhǔn)到標(biāo)準(zhǔn)MNI (montreal neurological institute)空間; 第七步,高斯平滑(smoothing), 平滑核(FWHW) = 4 mm; 最后, 進(jìn)行帶通濾波(band filter), 范圍為0.01~0.1 Hz。
2.6 數(shù)據(jù)分析
2.6.1 行為數(shù)據(jù)分析
為了檢驗(yàn)激發(fā)條件實(shí)驗(yàn)操作的有效性, 將兩種條件下被試為對(duì)手選擇的懲罰強(qiáng)度進(jìn)行配對(duì)樣本 t檢驗(yàn), 然后將被試選擇的懲罰等級(jí)的平均數(shù)作為被試反應(yīng)性攻擊的指標(biāo)。
2.6.2 fMRI數(shù)據(jù)分析
利用SPM8 (http://www.fil.ion.ucl.ac.uk/spm/)下的 marsbar工具包(Brett, Anton, Valabregue, &Poline, 2002)選取 OMPFC作為種子點(diǎn), 參考以往文獻(xiàn)中的做法(Beyer et al., 2015; Tzourio-Mazoyer et al., 2002), 分別選取 AAL模板中左右兩側(cè)MNI_Frontal_Med_Orb和MNI_Rectus制作左右兩側(cè)OMPFC的感興趣區(qū)(Region of Interest, ROI)。
利用REST (Song et al., 2011)的工具包下的FC工具計(jì)算 OMPFC與全腦的功能連接(voxel-wise),之后使用SPM8的多元回歸(multiple regression)模型分析方法計(jì)算OMPFC與全腦的功能連接(voxelwise)與反應(yīng)性攻擊相關(guān)的腦區(qū), 其中被試的性別、年齡作為協(xié)變量回歸掉。接下來(lái), 將上一步中顯著的區(qū)域保存為ROI, 利用REST工具包下的FC工具計(jì)算雙側(cè)OMPFC與這些ROI的FC值(ROI-wise),計(jì)算 FC值與反應(yīng)性攻擊的Pearson相關(guān)系數(shù); 最后, 利用REST工具包下的GCA工具計(jì)算這些ROI與OMPFC的GCA值(ROI-wise), 計(jì)算GCA值與反應(yīng)性攻擊的Pearson相關(guān)系數(shù)。
需要注意的是, 由于FC值不符合正態(tài)分布, 因此涉及到功能連接的兩步, 我們均進(jìn)行了 Fisher-z轉(zhuǎn)換, 并且利用z轉(zhuǎn)化后的值用于進(jìn)一步的分析。另外, 由于腦成像數(shù)據(jù)分析過(guò)程中存在多重比較而使結(jié)果帶有誤差, 因此在全腦分析中, 使用體素水平p< 0.001, 團(tuán)塊水平p <0.05(使用FDR進(jìn)行多重比較校正), 團(tuán)塊大小(cluster sizes) > 20體素作為閾值。
3 結(jié)果
3.1 行為實(shí)驗(yàn)結(jié)果
所有行為數(shù)據(jù)采用SPSS 16.0進(jìn)行分析。對(duì)激發(fā)與非激發(fā)條件下被試選擇的懲罰強(qiáng)度進(jìn)行配對(duì)樣本t檢驗(yàn)的結(jié)果表明, 激發(fā)條件下被試為對(duì)手選擇的懲罰強(qiáng)度顯著高于非激發(fā)條件下選擇的懲罰強(qiáng)度:激發(fā)條件:M= 3.91,SD= 1.71, 非激發(fā)條件:M= 3.11,SD= 1.49;t(38) = 3.44,p< 0.01, Cohen’d= 0.55。
3.2 腦成像數(shù)據(jù)結(jié)果
3.2.1 功能連接結(jié)果
將OMPFC與全腦的功能連接進(jìn)行多元回歸分析, 其中性別和年齡作為協(xié)變量控制。結(jié)果表明,左側(cè)OMPFC與右側(cè)角回、雙側(cè)尾狀核以及雙側(cè)內(nèi)側(cè)前額葉的功能連接與反應(yīng)性攻擊相關(guān)顯著, 右側(cè)OMPFC與右側(cè)尾狀核以及右內(nèi)側(cè)前額葉的功能連接與反應(yīng)性攻擊相關(guān)顯著, 見(jiàn)表 1。由于內(nèi)側(cè)前額葉不屬于皮層下腦區(qū), 因此后續(xù)的分析只針對(duì)左側(cè)OMPFC與尾狀核、角回以及右側(cè)OMPFC與尾狀核的功能連接與反應(yīng)性攻擊的關(guān)系。
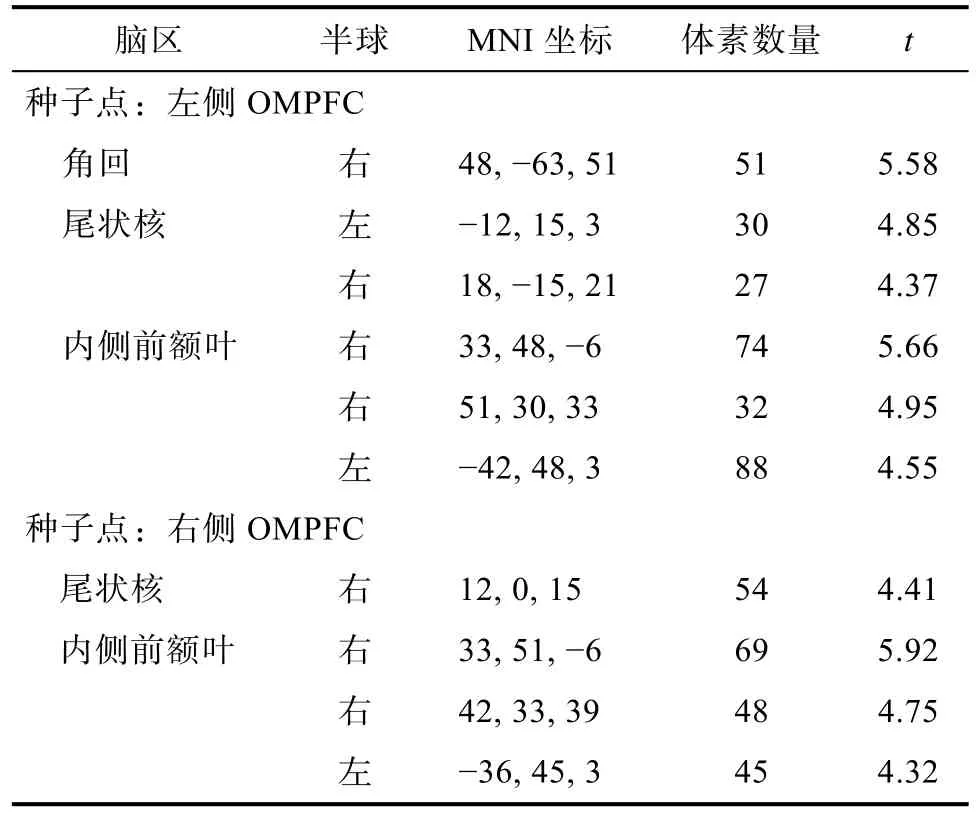
表1 雙側(cè)OMPFC功能連接結(jié)果
為了考察雙側(cè)OMPFC與皮層下腦區(qū)的功能連接與反應(yīng)性攻擊的關(guān)系, 我們進(jìn)一步提取上一步中顯著的皮層下腦區(qū)與OMPFC的功能連接值與反應(yīng)性攻擊做Pearson相關(guān)。結(jié)果顯示, 左側(cè) OMPFC與右側(cè)角回(r= ?0.44,p< 0.005, 見(jiàn)圖2)、左側(cè)OMPFC 與左側(cè)尾狀核(r= ?0.53,p< 0.001, 見(jiàn)圖3)、左側(cè) OMPFC 與右側(cè)尾狀核(r= ?0.59,p< 0.001,見(jiàn)圖4)以及右側(cè)OMPFC與右側(cè)尾狀核(r= ?0.61,p< 0.001, 見(jiàn)圖5)的FC值與反應(yīng)性攻擊均顯著負(fù)相關(guān), 并且不受激發(fā)條件的影響(即各功能連接值與兩種激發(fā)條件下被試的反應(yīng)性攻擊負(fù)相關(guān)均顯著,p< 0.05)。各功能連接值與反應(yīng)性攻擊的相關(guān)矩陣如表2所示。
3.2.2 格蘭杰因果分析結(jié)果
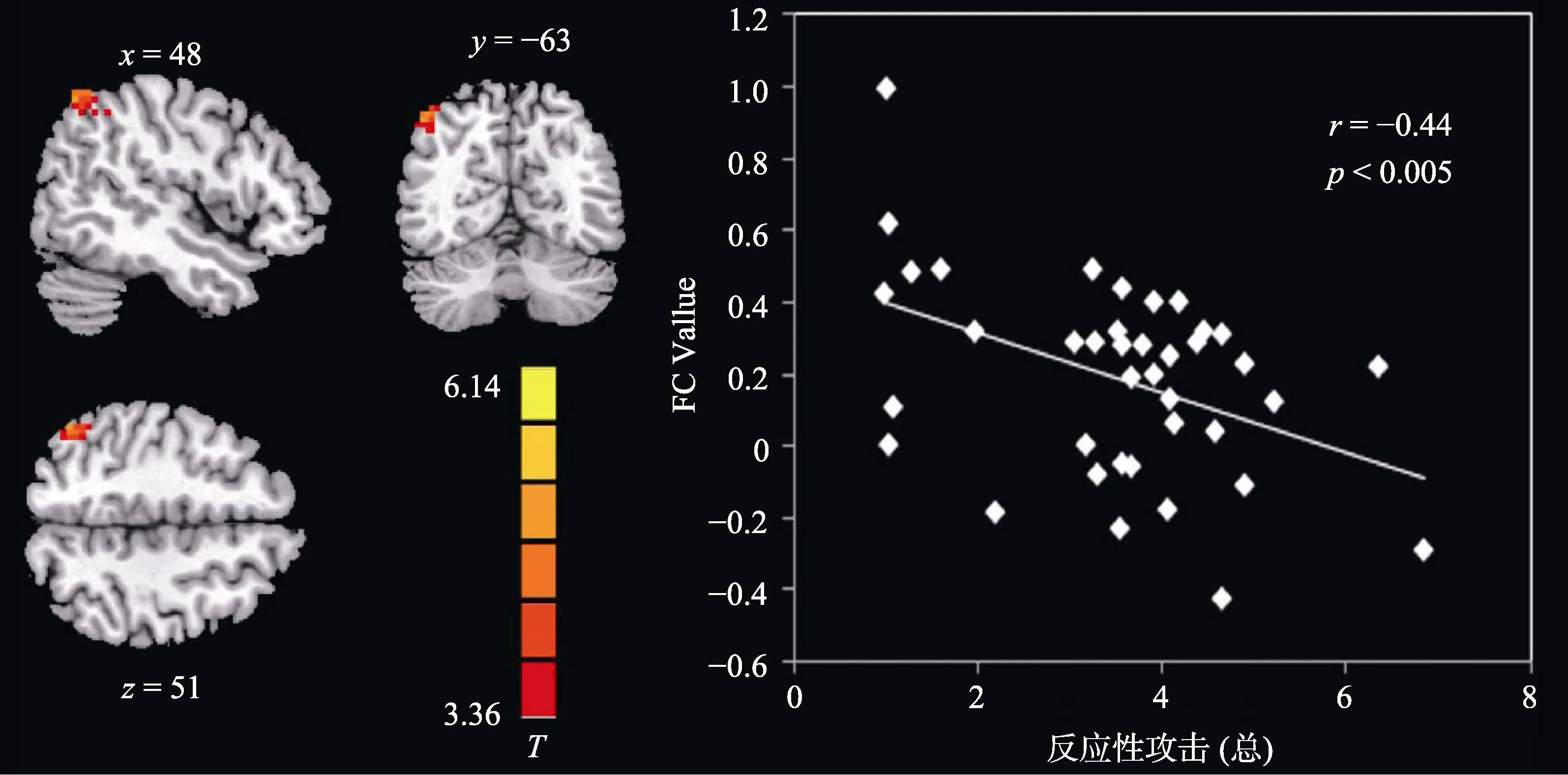
圖2 左側(cè)OMPFC?右側(cè)角回的功能連接與反應(yīng)性攻擊相關(guān)顯著(FC值使用z轉(zhuǎn)化之后的值)

圖3 左側(cè)OMPFC?左側(cè)尾狀核的功能連接與反應(yīng)性攻擊相關(guān)顯著(FC值使用z轉(zhuǎn)化之后的值)

圖4 左側(cè)OMPFC?右側(cè)尾狀核的功能連接與反應(yīng)性攻擊相關(guān)顯著(FC值使用z轉(zhuǎn)化之后的值)

圖5 右側(cè)OMPFC?右側(cè)尾狀核的功能連接與反應(yīng)性攻擊相關(guān)顯著(FC值使用z轉(zhuǎn)化之后的值)

表2 功能連接值與反應(yīng)性攻擊的相關(guān)矩陣
由于功能連接反映的是腦區(qū)之間的相關(guān)關(guān)系,不能反映信息輸入輸出的方向, 因此我們進(jìn)一步采用 GCA考察腦區(qū)之間效應(yīng)連接的異常與反應(yīng)性攻擊之間的關(guān)系。由于ROI水平的GCA分兩種情況,即信息輸入和信息輸出, 上述4個(gè)功能連接一共產(chǎn)生8種因果關(guān)系。即左側(cè)OMPFC→左側(cè)尾狀核、左側(cè)OMPFC→右側(cè)尾狀核、左側(cè)OMPFC→右側(cè)角回、右側(cè) OMPFC→右側(cè)尾狀核、左側(cè)尾狀核→左側(cè)OMPFC、右側(cè)尾狀核→左側(cè)OMPFC、右側(cè)角回→左側(cè)OMPFC以及右側(cè)尾狀核→右側(cè)OMPFC。GCA分析的結(jié)果顯示, 只有右側(cè)尾狀核→右側(cè)OMPFC的效應(yīng)連接與反應(yīng)性攻擊顯著負(fù)相關(guān)(r=?0.36,p< 0.05), 其他效應(yīng)連接與反應(yīng)性攻擊均不相關(guān)(ps > 0.05), 如圖6所示。進(jìn)一步對(duì)激發(fā)、非激發(fā)條件下的反應(yīng)性攻擊與右側(cè)尾狀核→右側(cè)OMPFC的GCA值進(jìn)行相關(guān)分析, 結(jié)果表明, 右側(cè)尾狀核→右側(cè)OMPFC與激發(fā)條件下的反應(yīng)性攻擊顯著負(fù)相關(guān)(r= ?0.33,p< 0.05), 而與非激發(fā)條件下的反應(yīng)性攻擊相關(guān)不顯著(r= ?0.31,p> 0.05)。效應(yīng)連接值與反應(yīng)性攻擊的相關(guān)矩陣如表 3所示,散點(diǎn)圖如圖6所示。

圖6 右側(cè)尾狀核→右側(cè)OMPFC的效應(yīng)連接與反應(yīng)性攻擊相關(guān)顯著(左為反應(yīng)性攻擊、右為激發(fā)條件下的反應(yīng)性攻擊)

表3 效應(yīng)連接值與反應(yīng)性攻擊的相關(guān)性統(tǒng)計(jì)
4 討論
研究通過(guò)采用 TAP變式, 運(yùn)用高空間分辨率的功能磁共振成像技術(shù), 探索性地對(duì)反應(yīng)性攻擊的神經(jīng)機(jī)制進(jìn)行了功能連接和效應(yīng)連接分析, 為進(jìn)一步理解反應(yīng)性攻擊的本質(zhì)提供了實(shí)證支持。在功能連接分析中, 我們發(fā)現(xiàn)右側(cè)角回、雙側(cè)尾狀核與左OMPFC的功能連接以及右側(cè)尾狀核與右 OMPFC的功能連接與反應(yīng)性攻擊相關(guān)顯著。在進(jìn)一步的格蘭杰因果分析中, 我們發(fā)現(xiàn)右側(cè)尾狀核到右側(cè)OMPFC的效應(yīng)連接在反應(yīng)性攻擊中起著重要的作用, 這與以往采用任務(wù)態(tài)功能磁共振技術(shù)的研究中發(fā)現(xiàn)前額葉皮層以及尾狀核在反應(yīng)性攻擊中起著重要作用的結(jié)果相一致(Kr?mer et al., 2007), 并且使用靜息態(tài)功能磁共振的手段對(duì)已有研究進(jìn)行有益補(bǔ)充。
OMPFC在情緒相關(guān)的決策, 尤其在社會(huì)互動(dòng)背景下的情緒性決策中起著重要的作用(Rudebeck et al., 2008), 如在Gage個(gè)案以及一些類似案例中,當(dāng) OMPFC受損之后, 個(gè)體變得暴躁、攻擊性強(qiáng)(Blair, 2012; Damasio et al., 1994)。具體而言, OMPFC與攻擊沖動(dòng)、敵意反應(yīng)的抑制有關(guān)(Koenigs & Tranel,2007; Pietrini et al., 2000)。在本研究中, 我們選取OMPFC作為種子點(diǎn)考察正常人群OMPFC與其他腦區(qū)的關(guān)系在反應(yīng)性攻擊中的作用, 結(jié)果發(fā)現(xiàn)OMPFC與尾狀核的連接與反應(yīng)性攻擊相關(guān)顯著,這說(shuō)明不僅 OMPFC損傷的病人反應(yīng)性攻擊增多,而且在正常人群中, OMPFC與尾狀核的連接也與反應(yīng)性攻擊密切相關(guān)。因此, 在本研究中選取OMPFC作為種子點(diǎn), 有效地反映了前額葉皮質(zhì)與其他腦區(qū)的協(xié)同作用在反應(yīng)性攻擊中的作用。
尾狀核是確保個(gè)體靈活應(yīng)對(duì)環(huán)境的重要區(qū)域,對(duì)獎(jiǎng)賞的事件或者信號(hào)做出反應(yīng)而對(duì)不帶獎(jiǎng)賞性質(zhì)的信號(hào)則沒(méi)有反應(yīng)(Zink, Pagnoni, Martin-Skurski,Chappelow, & Berns, 2004)。然而, 有幾種類型的非獎(jiǎng)賞刺激也可以激活尾狀核, 這表明尾狀核能夠編碼所有顯著刺激, 因?yàn)殡m然這些刺激本身不具有獎(jiǎng)賞的性質(zhì), 但是被個(gè)體認(rèn)為是具有獎(jiǎng)賞性質(zhì)的(Zink,Pagnoni, Martin, Dhamala, & Berns, 2003)。研究者采用任務(wù)態(tài)功能磁共振技術(shù)探討反應(yīng)性攻擊的神經(jīng)機(jī)制時(shí)進(jìn)一步發(fā)現(xiàn)了, 尾狀核的激活可能與實(shí)施反應(yīng)性攻擊這一行為被個(gè)體認(rèn)知為一種獎(jiǎng)賞信號(hào)有關(guān)。例如, Kr?mer等人(2007)的研究采用TAP和任務(wù)態(tài)fMRI發(fā)現(xiàn)了尾狀核的激活程度與被試為對(duì)手選擇的懲罰強(qiáng)度成正相關(guān)關(guān)系, 也就是說(shuō)當(dāng)被試為虛擬對(duì)手選擇了更大的懲罰等級(jí)時(shí), 尾狀核被顯著激活。Beyer等人(2014)的研究也表明, 尾狀核的激活程度與反應(yīng)性攻擊之間有著相關(guān)關(guān)系。另外一項(xiàng)研究發(fā)現(xiàn)早期反饋階段尾狀核的激活情況預(yù)測(cè)了后期決定階段反應(yīng)性攻擊的強(qiáng)度(Kr?mer et al.,2011)。這些使用 TAP的研究一致地發(fā)現(xiàn)了尾狀核在反應(yīng)性攻擊產(chǎn)生過(guò)程中的作用, 當(dāng)反應(yīng)性攻擊行為被認(rèn)為是受到獎(jiǎng)賞時(shí)則尾狀核得到激活。
其他研究采用獎(jiǎng)賞相關(guān)的任務(wù)發(fā)現(xiàn), 患有與攻擊性增強(qiáng)有關(guān)的精神疾病或者人格障礙的個(gè)體對(duì)獎(jiǎng)賞的敏感性增強(qiáng)。例如, V?llm等(2007)對(duì)有反社會(huì)人格或者邊緣人格障礙的個(gè)體以及正常個(gè)體進(jìn)行研究, 在任務(wù)中對(duì)被試的某些反應(yīng)給予獎(jiǎng)賞而對(duì)另外一部分反應(yīng)則不給獎(jiǎng)賞, 研究結(jié)果顯示, 對(duì)比獎(jiǎng)賞與非獎(jiǎng)賞條件(即獎(jiǎng)賞?非獎(jiǎng)賞), 有人格障礙的個(gè)體比正常控制組的個(gè)體的尾狀核活動(dòng)更低。隨后的兩項(xiàng)研究進(jìn)一步證明, 這種有人格障礙的人之所以在獎(jiǎng)賞和非獎(jiǎng)賞條件下的尾狀核活動(dòng)差異減小, 是由于其在非獎(jiǎng)賞條件下的活動(dòng)高于正常群體所導(dǎo)致的(Gatzke-Kopp et al., 2009; Finger et al.,2008)。也就是說(shuō), 對(duì)于正常被試來(lái)講, 在獎(jiǎng)賞條件下尾狀核激活, 而在非獎(jiǎng)賞條件下, 尾狀核激活強(qiáng)度下降或者不被激活; 而對(duì)于患有人格障礙的個(gè)體(例如, 邊緣性人格障礙等)來(lái)說(shuō), 在獎(jiǎng)賞和非獎(jiǎng)賞條件下尾狀核都被激活。綜上, 尾狀核的激活與反應(yīng)性攻擊密切相關(guān), 那些攻擊性較強(qiáng)的個(gè)體對(duì)獎(jiǎng)賞的敏感性增強(qiáng), 當(dāng)刺激不具有獎(jiǎng)賞性質(zhì)時(shí), 尾狀核仍然激活, 甚至, 當(dāng)實(shí)施反應(yīng)性攻擊被認(rèn)為“受到了獎(jiǎng)賞”時(shí), 尾狀核也會(huì)相應(yīng)地被更大程度激活。
在發(fā)現(xiàn)與反應(yīng)性攻擊相關(guān)的關(guān)鍵腦區(qū)的基礎(chǔ)上, 越來(lái)越多的研究者指出反應(yīng)性攻擊的產(chǎn)生是由于前額葉皮層與皮層下腦區(qū)功能連接失調(diào)引起的(Rosell & Siever, 2015; Siever, 2008; Nelson &Trainor, 2007)。一項(xiàng)研究同時(shí)使用了正電子斷層掃描(Positron Emission Tomography, PET)和任務(wù)態(tài)功能磁共振技術(shù)的技術(shù)發(fā)現(xiàn)了尾狀核和執(zhí)行功能區(qū)域之間在結(jié)構(gòu)和功能上都存在著很強(qiáng)的聯(lián)系(Grahn,Parkinson, & Owen 2009)。da Cunha-Bang 等人(2017)采用點(diǎn)減攻擊范式(point subtraction aggression paradigm, PSAP)比較正常個(gè)體和暴力罪犯在受到敵意激發(fā)(輸?shù)舸饚?后大腦的活動(dòng)情況, 結(jié)果顯示比起正常個(gè)體, 暴力罪犯在受到敵意激發(fā)后, 反應(yīng)性攻擊更強(qiáng), 尾狀核的激活強(qiáng)度升高, 并且尾狀核?前額葉的功能連接降低。我們的研究結(jié)果一致的發(fā)現(xiàn)了, 在正常群體中, 尾狀核?前額葉(主要是OMPFC)的功能連接與反應(yīng)性攻擊負(fù)相關(guān)。
此外, 由于功能連接分析只能得到腦區(qū)之間的相關(guān)關(guān)系, 而無(wú)法確定影響方向的問(wèn)題, 也就是說(shuō)反應(yīng)性攻擊的增加到底是由于尾狀核對(duì)OMPFC自下而上信息傳遞存在問(wèn)題還是由于OMPFC對(duì)尾狀核自上而下的調(diào)控功能不足導(dǎo)致的呢?因此, 在功能分析的基礎(chǔ)上, 我們進(jìn)一步使用格蘭杰因果分析對(duì)效應(yīng)連接與反應(yīng)性攻擊的關(guān)系進(jìn)行探討。結(jié)果表明, 右側(cè)尾狀核到右側(cè)OMPFC的效應(yīng)連接與反應(yīng)性攻擊呈顯著的負(fù)相關(guān)關(guān)系, 右側(cè)尾狀核到右側(cè)OMPFC的GCA值越大, 則反應(yīng)性攻擊越弱。因此,我們推測(cè)反應(yīng)性攻擊的增強(qiáng)是由于個(gè)體大腦內(nèi)部尾狀核對(duì)OMPFC的自下而上的信息傳遞出現(xiàn)問(wèn)題導(dǎo)致的。鑒于沒(méi)有直接的相關(guān)研究可以對(duì)此作出解釋, Sagvolden, Johansen, Aase和Russell (2005)關(guān)于注意缺損障礙(Attention Deficit Hyperactivity Disorder,ADHD)的理論為我們提供了新的思路, 他們認(rèn)為ADHD的發(fā)生與多巴胺系統(tǒng)的信號(hào)傳遞失調(diào)有關(guān)。后來(lái)該理論也被用來(lái)解釋其他行為問(wèn)題的產(chǎn)生, 如外部行為障礙(Externalizing Behavior Disorders;Shannon, Sauder, Beauchaine, & Gatzke-Kopp, 2009)。已有關(guān)于多巴胺系統(tǒng)的研究顯示, 在大腦內(nèi)部存在4條多巴胺回路, 其中兩條回路與攻擊行為相關(guān):第一條包括尾狀核、杏仁核、海馬等皮層下腦區(qū); 另外一條包括背外側(cè)前額葉、前部扣帶回、顳葉和OMPFC等腦區(qū)(Gatzke-Kopp & Beauchaine, 2007)。有研究者進(jìn)一步指出, 綜合考慮多條多巴胺回路之間的關(guān)系可能對(duì)于揭示目標(biāo)導(dǎo)向行為更為準(zhǔn)確和合理(Bj?rklund & Dunnett, 2007)。由于尾狀核和OMPFC都是多巴胺回路的一部分, 因此, 我們推測(cè)比起正常的個(gè)體, 那些反應(yīng)性攻擊較強(qiáng)的個(gè)體也有可能是由于大腦內(nèi)部多巴胺系統(tǒng)存在問(wèn)題, 使得某些行為與可能結(jié)果之間的關(guān)系不能被正確認(rèn)識(shí),從而導(dǎo)致在某些情境(如敵意情境)下, 皮層下那條回路被“誤”激活, 致使尾狀核對(duì) OMPFC的自下而上的信息傳遞出現(xiàn)了問(wèn)題。但是反應(yīng)性攻擊與ADHD以及外部行為障礙還存在很大的差異, 因此至于這種尾狀核到右側(cè)OMPFC的效應(yīng)連接中出現(xiàn)的信息傳遞問(wèn)題究竟是否是多巴胺系統(tǒng)的失調(diào)導(dǎo)致尚需采用PET等技術(shù)進(jìn)一步研究來(lái)揭示和證明。另外, 該分析結(jié)果還顯示這種相關(guān)關(guān)系在激發(fā)條件下顯著,而在非激發(fā)條件下不顯著, 這可能是由于激發(fā)條件下尾狀核更容易被“誤”激活引起的。
值得注意的是, 很多早期基于病人的研究提出杏仁核與反應(yīng)性攻擊相關(guān), 例如, 在對(duì)癲癇病人的研究中, 一致地發(fā)現(xiàn)了電刺激杏仁核導(dǎo)致會(huì)攻擊反應(yīng)(Mark, Sweet, & Ervin, 1975), 切除杏仁核或者對(duì)杏仁核結(jié)構(gòu)造成損傷則可以用來(lái)降低反應(yīng)性攻擊(Lee et al., 1998; Narabayashi, Nagao, Saito, Yoshida,& Nagahata, 1963)。近期一些研究也證明了杏仁核以及杏仁核?前額葉皮質(zhì)的功能連接在反應(yīng)性攻擊中的作用, 例如上文提到的 da Cunha-Bang等人(2017)的研究結(jié)果也顯示比起正常人, 暴力罪犯在受到敵意激發(fā)后, 反應(yīng)性攻擊更強(qiáng), 尾狀核和杏仁核的激活強(qiáng)度升高, 并且尾狀核?前額葉以及杏仁核?前額葉的功能連接降低。比起普通罪犯, 患有精神疾病的罪犯在靜息狀態(tài)下杏仁核?內(nèi)側(cè)前額葉的功能連接較低(Motzkin et al., 2011)。而在本研究中, 卻未能發(fā)現(xiàn)杏仁核?OMPFC之間的功能連接與反應(yīng)性攻擊相關(guān), 據(jù)推測(cè), 可能是由以下原因?qū)е碌模菏紫? 越來(lái)越多的研究者提出, 杏仁核不是一個(gè)功能單一的區(qū)域, 也就是說(shuō)不同的區(qū)域可能功能存在差異, 其中被廣泛認(rèn)同的一個(gè)觀點(diǎn)是杏仁核主要由基底外側(cè)、中央內(nèi)側(cè)以及表面組成(Sah,Faber, Lopez de Lopez, & Power, 2003), 其中表面的作用很少得到揭示, 而基底外側(cè)主要負(fù)責(zé)感覺(jué)信息的輸入, 中央內(nèi)側(cè)主要是負(fù)責(zé)輸出到其他腦區(qū)。就反應(yīng)性攻擊來(lái)講, 外側(cè)杏仁核負(fù)責(zé)整合輸入的信息并激發(fā)中央核喚起“戰(zhàn)或逃”的反應(yīng)(Davis & Whalen,2001; Maren, 2001; LeDoux, 2000, 1998), 中央核則負(fù)責(zé)放大或抑制反應(yīng)性攻擊的傾向(D?biec, 2005;Huber, Veinante, & Stoop, 2005)。以往關(guān)于腦形態(tài)學(xué)的研究也發(fā)現(xiàn), 整個(gè)杏仁核的體積與特質(zhì)攻擊相關(guān)不顯著, 而背側(cè)杏仁核的體積則與特質(zhì)攻擊相關(guān)(Gopal et al., 2013), 因此, 在未來(lái)的研究中應(yīng)該將杏仁核區(qū)分為不同的亞區(qū)來(lái)考察杏仁核?OMPFC的功能連接與反應(yīng)性攻擊的關(guān)系。其次, 在以往使用 TAP范式的任務(wù)態(tài)研究中一致地沒(méi)有發(fā)現(xiàn)杏仁核的激活(Kr?mer et al., 2007; Lotze et al., 2007),因此, 是否 TAP范式不能很好檢測(cè)到杏仁核的激活甚至TAP范式的行為指標(biāo)也不能反映出杏仁核?前額葉皮質(zhì)的功能連接與反應(yīng)性攻擊的關(guān)系也有待進(jìn)一步研究。
當(dāng)然, 本研究也存在一些不足。例如, 種子點(diǎn)的選取差異可能會(huì)對(duì)結(jié)果產(chǎn)生影響, 因此, 在未來(lái)的研究中可以選擇其他的種子點(diǎn)考察其與其他腦區(qū)的協(xié)同作用在反應(yīng)性攻擊產(chǎn)生過(guò)程中的作用。另外, 本研究以普通大學(xué)生為被試, 得到的結(jié)論是否適用于暴力罪犯或者患有精神分裂癥等精神疾病的個(gè)體, 還有待進(jìn)一步的考察。
5 結(jié)論
本研究采用被廣泛使用的TAP, 運(yùn)用靜息態(tài)功能成像技術(shù)對(duì)反應(yīng)性攻擊的腦機(jī)制進(jìn)行了研究。功能連接的結(jié)果發(fā)現(xiàn)了雙側(cè)OMPFC與尾狀核之間的功能連接與反應(yīng)性攻擊呈負(fù)相關(guān)。格蘭杰因果分析的結(jié)果進(jìn)一步表明右側(cè)尾狀核到右側(cè)OMPFC的效應(yīng)連接與反應(yīng)性攻擊負(fù)相關(guān)。與以往采用任務(wù)功能態(tài)的研究一致地發(fā)現(xiàn)了尾狀核在反應(yīng)性攻擊中的作用, 為進(jìn)一步揭示反應(yīng)性攻擊的神經(jīng)機(jī)制提供了部分證據(jù), 是首個(gè)利用靜息態(tài)功能連接和效應(yīng)連接對(duì)反應(yīng)性攻擊進(jìn)行系統(tǒng)分析的研究。
Anderson, C. A., & Bushman, B. J. (2002). Human aggression.Annual Review of Psychology, 53(1), 27?51.
Bettencourt, B. A., Talley, A., Benjamin, A. J., & Valentine, J.(2006). Personality and aggressive behavior under provoking and neutral conditions: A meta-analytic review.Psychological Bulletin, 132(5), 751?777.
Beyer, F., Münte, T. F., Erdmann, C., & Kr?mer, U. M. (2014).Emotional reactivity to threat modulates activity in mentalizing network during aggression.Social Cognitive and Affective Neuroscience, 9(10), 1552?1560.
Beyer, F., Münte, T. F., G?ttlich, M., & Kr?mer, U. M. (2015).Orbitofrontal cortex reactivity to angry facial expression in a social interaction correlates with aggressive behavior.Cerebral Cortex, 25(9), 3057?3063.
Bj?rklund, A., & Dunnett, S. B. (2007). Dopamine neuron systems in the brain: An update.Trends in Neurosciences,30(5), 194?202.
Blair, R. J. R. (2012). Considering anger from a cognitive neuroscience perspective.Wiley Interdisciplinary Reviews:Cognitive Science, 3(1), 65?74.
Brett, M., Anton, J. L., Valabregue, R., & Poline, J. B. (2002).Region of interest analysis using the MarsBar toolbox for SPM 99.NeuroImage, 16(2), S497.
Chen, G., Hamilton, J. P., Thomason, M. E., Gotlib, I. H., Saad,Z. S., & Cox, R. W. (2009). Granger causality via vector auto-regression tuned for fMRI data analysis. InProceedings of the 17th annual scientific meeting and exhibition(Vol.17, p. 1718). Honolulu, Hawaii.
Coccaro, E. F., McCloskey, M. S., Fitzgerald, D. A., & Phan,K. L. (2007). Amygdala and orbitofrontal reactivity to social threat in individuals with impulsive aggression.Biological Psychiatry, 62(2), 168–178.
Coccaro, E. F., Sripada, C. S., Yanowitch, R. N., & Phan, K. L.(2011). Corticolimbic function in impulsive aggressive behavior.Biological Psychiatry, 69(12), 1153?1159.
da Cunha-Bang, S., Fisher, P. M., Hjordt, L. V., Perfalk, E.,Skibsted, A. P., Bock, C., ... Knudsen, G. M. (2017).Violent offenders respond to provocations with high amygdala and striatal reactivity.Social Cognitive and Affective Neuroscience, 12(5), 802?810.
Damasio, H., Grabowski, T., Frank, R., Galaburda, A. M., &Damasio, A. R. (1994). The return of Phineas Gage: Clues about the brain from the skull of a famous patient.Science,264(5162), 1102?1105.
Davis, M., & Whalen, P. J. (2001). The amygdala: Vigilance and emotion.Molecular Psychiatry, 6(1), 13?34.
D?biec, J. (2005). Peptides of love and fear: Vasopressin and oxytocin modulate the integration of information in the amygdala.BioEssays, 27(9), 869?873.
Finger, E. C., Marsh, A. A., Mitchell, D. G., Reid, M. E., Sims,C., Budhani, S., ... Blair, J. R. (2008). Abnormal ventromedial prefrontal cortex function in children with psychopathic traits during reversal learning.Archives of General Psychiatry, 65(5), 586?594.
Fite, P. J., Rubens, S. L., Preddy, T. M., Raine, A., & Pardini,D. A. (2014). Reactive/proactive aggression and the development of internalizing problems in males: The moderating effect of parent and peer relationships.Aggressive Behavior, 40(1), 69?78.
Fulwiler, C. E., King, J. A., & Zhang, N. Y. (2012). Amygdalaorbitofrontal resting state functional connectivity is associated with trait anger.Neuroreport, 23(10), 606?610.
Gatzke-Kopp, L. M., & Beauchaine, T. P. (2007). Central nervous system substrates of impulsivity: Implications for the development of attention-deficit/hyperactivity disorder and conduct disorder. In D. Coch, G. Dawson, & K. W.Fischer (Eds.),Human behavior, Learning, and the developing brain: Atypical development(pp. 239?263). New York:Guilford.
Gatzke-Kopp, L. M., Beauchaine, T. P., Shannon, K. E.,Chipman, J., Fleming, A. P., Crowell, S. E., ... Aylward, E.(2009). Neurological correlates of reward responding in adolescents with and without externalizing behavior disorders.Journal of Abnormal Psychology, 118(1), 203?213.
Giancola, P. R., & Parrott, D. J. (2008). Further evidence for the validity of the Taylor aggression paradigm.Aggressive Behavior, 34(2), 214?229.
Glenn, A. L., & Yang, Y. L. (2012). The potential role of the striatum in antisocial behavior and psychopathy.Biological Psychiatry, 72(10), 817?822.
Gopal, A., Clark, E., Allgair, A., D'Amato, C., Furman, M.,Gansler, D. A., & Fulwiler, C. (2013). Dorsal/ventral parcellation of the amygdala: Relevance to impulsivity and aggression.Psychiatry Research: Neuroimaging, 211(1),24?30.
Grahn, J. A., Parkinson, J. A., & Owen, A. M. (2009). The role of the basal ganglia in learning and memory:Neuropsychological studies.Behavioural Brain Research,199(1), 53?60.
Greicius, M. D., Flores, B. H., Menon, V., Glover, G. H.,Solvason, H. B., Kenna, H., ... Schatzberg, A. F. (2007).Resting-state functional connectivity in major depression:Abnormally increased contributions from subgenual cingulate cortex and thalamus.Biological Psychiatry, 62(5), 429?437.
Hahn, A., Stein, P., Windischberger, C., Weissenbacher, A.,Spindelegger, C., Moser, E., ... Lanzenberger, R. (2011).Reduced resting-state functional connectivity between amygdala and orbitofrontal cortex in social anxiety disorder.NeuroImage, 56(3), 881?889.
Hamilton, J. P., Chen, G., Thomason, M. E., Schwartz, M. E.,& Gotlib, I. H. (2011). Investigating neural primacy in major depressive disorder: Multivariate Granger causality analysis of resting-state fMRI time-series data.Molecular Psychiatry, 16(7), 763?772.
Herpertz, S. C., Nagy, K., Ueltzh?ffer, K., Schmitt, R.,Mancke, F., Schmahl, C., & Bertsch, K. (2017). Brain mechanisms underlying reactive aggression in borderline personality disorder—Sex matters.Biological Psychiatry,82(4), 257?266.
Hoptman, M. J., D'Angelo, D., Catalano, D., Mauro, C. J.,Shehzad, Z. E., Kelly, A. M. C., ... Milham, M. P. (2010).Amygdalofrontal functional disconnectivity and aggression in schizophrenia.Schizophrenia Bulletin, 36(5), 1020?1028.
Huber, D., Veinante, P., & Stoop, R. (2005). Vasopressin and oxytocin excite distinct neuronal populations in the central amygdala.Science, 308(5719), 245?248.
Koenigs, M., & Tranel, D. (2007). Irrational economic decision-making after ventromedial prefrontal damage:Evidence from the ultimatum game.Journal of Neuroscience,27(4), 951?956.
Kr?mer, U. M., Jansma, H., Tempelmann, C., & Münte, T. F.(2007). Tit-for-tat: The neural basis of reactive aggression.NeuroImage, 38(1), 203?211.
Kr?mer, U. M., Riba, J., Richter, S., & Münte, T. F. (2011). An fMRI study on the role of serotonin in reactive aggression.PLoS One, 6(11), e27668.
LeDoux, J. (1998). Fear and the brain: Where have we been,and where are we going?Biological Psychiatry, 44(12),1229?1238.
LeDoux, J. E. (2000). Emotion circuits in the brain.Annual Review of Neuroscience, 23, 155?184.
Lee, G. P., Bechara, A., Adolphs, R., Arena, J., Meador, K. J.,Loring, D. W., & Smith, J. R. (1998). Clinical and physiological effects of stereotaxic bilateral amygdalotomy for intractable aggression.The Journal of Neuropsychiatry and Clinical Neurosciences, 10(4), 413?420.
Liu, Y. L., Teng, Z. J., Lan, H. Y., Zhang, X., & Yao, D. Z.(2015). Short-term effects of prosocial video games on aggression: An event-related potential study.Frontiers in Behavioral Neuroscience, 9, 193.
Lotze, M., Veit, R., Anders, S., & Birbaumer, N. (2007).Evidence for a different role of the ventral and dorsal medial prefrontal cortex for social reactive aggression: An interactive fMRI study.NeuroImage, 34(1), 470–478.
Maren, S. (2001). Neurobiology of Pavlovian fear conditioning.Annual Review of Neuroscience, 24, 897?931.
Mark, V. H., Sweet, W., & Ervin, F. (1975). Deep temporal lobe stimulation and destructive lesions in episodically violent temporal lobe epileptics. In W. Fields & W. Sweet(Eds.),Neural bases of violence and aggression(pp.379?400). St. Louis: Warren H. Greem, Inc.
McCloskey, M. S., Phan, K. L., Angstadt, M., Fettich, K. C.,Keedy, S., & Coccaro, E. F. (2016). Amygdala hyperactivation to angry faces in intermittent explosive disorder.Journal of Psychiatric Research, 79, 34?41.
McEwen, C. A., & McEwen, B. S. (2017). Social structure,adversity, toxic stress, and intergenerational poverty: An early childhood model.Annual Review of Sociology, 43,445?472.
Motzkin, J. C., Newman, J. P., Kiehl, K. A., & Koenigs, M.(2011). Reduced prefrontal connectivity in psychopathy.Journal of Neuroscience, 31(48), 17348?17357.
Narabayashi, H., Nagao, T., Saito, Y., Yoshida, M., & Nagahata,M. (1963). Stereotaxic amygdalotomy for behavior disorders.Archives of Neurology, 9(1), 1?16.
Nelson, R. J., & Trainor, B. C. (2007). Neural mechanisms of aggression.Nature Reviews Neuroscience, 8(7), 536?546.
New, A. S., Hazlett, E. A., Buchsbaum, M. S., Goodman, M.,Mitelman, S. A., Newmark, R., ... Siever, L. J. (2007).Amygdala-prefrontal disconnection in borderline personality disorder.Neuropsychopharmacology, 32(7), 1629?1640.
Pietrini, P., Guazzelli, M., Basso, G., Jaffe, K., & Grafman, J.(2000). Neural correlates of imaginal aggressive behavior assessed by positron emission tomography in healthy subjects.American Journal of Psychiatry, 157(11), 1772?1781.
Ramirez, J. M., & Andreu, J. M. (2006). Aggression, and some related psychological constructs (anger, hostility, and impulsivity) Some comments from a research project.Neuroscience and Biobehavioral Reviews, 30(3), 276?291.
Riva, P., Gabbiadini, A., Lauro, L. J. R., Andrighetto, L.,Volpato, C., & Bushman, B. J. (2017). Neuromodulation can reduce aggressive behavior elicited by violent video games.Cognitive, Affective, and Behavioral Neuroscience, 17(2),452?459.
Rosell, D. R., & Siever, L. J. (2015). The neurobiology of aggression and violence.CNS Spectrums, 20(3), 254–279.
Rudebeck, P. H., Bannerman, D. M., & Rushworth, M. F. S.(2008). The contribution of distinct subregions of the ventromedial frontal cortex to emotion, social behavior,and decision making.Cognitive, Affective, and Behavioral Neuroscience, 8(4), 485?497.
Sagvolden, T., Johansen, E. B., Aase, H., & Russell, V. A.(2005). A dynamic developmental theory of attentiondeficit/hyperactivity disorder (ADHD) predominantly hyperactive/impulsive and combined subtypes.Behavioral and Brain Sciences, 28(3), 397?419.
Sah, P., Faber, E. S. L., Lopez de Lopez, M., & Power, J. P.(2003). The amygdaloid complex: Anatomy and physiology.Physiological Reviews, 83(3), 803?834.
Shannon, K. E., Sauder, C., Beauchaine, T. P., & Gatzke-Kopp,L. M. (2009). Disrupted effective connectivity between the medial frontal cortex and the caudate in adolescent boys with externalizing behavior disorders.Criminal Justice andBehavior, 36(11), 1141?1157.
Siever, L. J. (2008). Neurobiology of aggression and violence.American Journal of Psychiatry, 165(4), 429–442.
Song, X. W., Dong, Z. Y., Long, X. Y., Li, S. F., Zuo, X. N.,Zhu, C. Z., ... Zang, Y. F. (2011). REST: A toolkit for resting-state functional magnetic resonance imaging data processing.PLoS One, 6(9), e25031.
Takeuchi, H., Taki, Y., Hashizume, H., Sassa, Y., Nagase, T.,Nouchi, R., & Kawashima, R. (2012). The association between resting functional connectivity and creativity.Cerebral Cortex, 22(12), 2921?2929.
Taylor, S. P. (1967). Aggressive behavior and physiological arousal as a function of provocation and the tendency to inhibit aggression.Journal of Personality, 35(2), 297?310.
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello,F., Etard, O., Delcroix, N., ... Joliot, M. (2002). Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain.NeuroImage, 15(1), 273?289.
V?llm, B., Richardson, P., McKie, S., Elliott, R., Dolan, M., &Deakin, B. (2007). Neuronal correlates of reward and loss in cluster B personality disorders: A functional magnetic resonance imaging study.Psychiatry Research: Neuroimaging,156(2), 151?167.
Wu, Q. Z., Li, D. M., Kuang, W. H., Zhang, T. J., Lui, S.,Huang, X. Q., ... Gong, Q. Y. (2011). Abnormal regional spontaneous neural activity in treatment-refractory depression revealed by resting-state fMRI.Human Brain Mapping,32(8), 1290?1299.
Yan, C. G., & Zang, Y. F. (2010). DPARSF: A MATLAB toolbox for “pipeline” data analysis of resting-state fMRI.Frontiers in Systems Neuroscience, 413.
Zeng, L. L., Shen, H., Liu, L., Wang, L. B., Li, B. J., Fang,P., ... Hu, D. W. (2012). Identifying major depression using whole-brain functional connectivity: A multivariate pattern analysis.Brain, 135(5), 1498?1507.
Zink, C. F., Pagnoni, G., Martin, M. E., Dhamala, M., & Berns,G. S. (2003). Human striatal response to salient nonrewarding stimuli.Journal of Neuroscience, 23(22), 8092?8097.
Zink, C. F., Pagnoni, G., Martin-Skurski, M. E., Chappelow, J.C., & Berns, G. S. (2004). Human striatal responses to monetary reward depend on saliency.Neuron, 42(3), 509?517.
Zuo, X. N., Di Martino, A., Kelly, C., Shehzad, Z. E., Gee, D.G., Klein, D. F., ... Milham, M. P. (2010). The oscillating brain:Complex and reliable.NeuroImage, 49(2), 1432?1445.
猜你喜歡
中華詩(shī)詞(2022年6期)2022-12-31 06:41:24
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國(guó)語(yǔ)大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
中國(guó)科技論壇(2017年7期)2017-07-25 08:49:53
汽車工程學(xué)報(bào)(2017年2期)2017-07-05 08:13:02
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國(guó)際漢語(yǔ)學(xué)報(bào)(2016年1期)2017-01-20 08:21:20
