誘變育種提高嗜鹽四聯球菌精氨酸和瓜氨酸利用能力
2018-07-18 09:39:30雷慶子王博堵國成方芳
食品與發酵工業 2018年6期
雷慶子,王博,堵國成,方芳*
1(江南大學 生物工程學院,江蘇 無錫,214122)2(工業生物技術教育部重點實驗室,江蘇 無錫,214122 ) 3(糖化學與生物技術教育部重點實驗室,江蘇 無錫,214122 )
氨基甲酸乙酯(ethyl carbamate,EC)是一種存在于發酵食品中的2A類致癌物,具有遺傳毒性,可導致肺腫瘤、淋巴癌、肝癌和皮膚癌[1-2]。EC曾被用作抗腫瘤藥物,由于具有致癌性,現已停止臨床使用[3]。攝入體內的EC經細胞色素P450的羥基化作用和氧化作用生成可損傷DNA的8-羥基-脫氧鳥苷和乙烯基-氨基-甲酸酯環氧化物,通過破壞DNA結構引發癌變[4-6]。
發酵食品中形成EC的主要前體包括氰化物、氨甲酰磷酸、尿素和瓜氨酸[7]。醬油生產過程中容易積累有害物氨基甲酸乙酯,這種現象在高鹽稀態法發酵生產醬油的過程中尤為顯著[8-9]。高鹽稀態醬油中,EC的主要前體是瓜氨酸,由乳酸菌通過精氨酸脫亞氨酶途徑(arginine deiminase pathway,簡稱ADI途徑)代謝精氨酸生成[10-14]。ADI途徑包括3個關鍵酶:精氨酸脫亞氨酶(arginine deiminase, ADI)、鳥氨酸轉氨甲酰酶(omithine transcarbamylase, OTC)和氨基甲酰激酶(carbamate kinase, CK),分別由arcA、arcB、arcC基因編碼[15-16]。由于醬油發酵是一個混菌發酵的食品體系,生產上不允許使用基因工程菌或用基因工程手段改造生產用菌株。通過內源微生物干預是實現控制和減少醬油中EC及其前體的有效措施。
尋找來源于醬醪微生物體系并可利用精氨酸且不積累瓜氨酸的菌株,通過在發酵過程中強化這一菌株來降低醬油中的EC前體瓜氨酸,可為建立消除食品混菌發酵體系中的有害微生物代謝物的方法提供研究基礎和理論支持。嗜鹽四聯球菌(Tetragenococcushalophilus)是存在于醬油、泰國魚醬、果汁等高鹽或高糖環境中的一類乳酸菌[17-19]。它具有提升食品風味、改善食品品質的功能[20-21]。已有研究表明,來源于醬醪的嗜鹽四聯球菌R23可利用醬醪中的精氨酸和瓜氨酸,從而減少EC前體,起到控制EC含量的作用[22]。然而,體系中葡萄糖和精氨酸含量對嗜鹽四聯球菌利用或積累瓜氨酸均有顯著影響:當體系中葡萄糖和精氨酸含量增加時,嗜鹽四聯球菌利用精氨酸和瓜氨酸能力顯著下降[22-23]。這說明,嗜鹽四聯球菌用于醬油發酵時,其精氨酸和瓜氨酸利用能力受環境因素制約。因此,通過誘變育種的方法增強嗜鹽四聯球菌利用精氨酸和瓜氨酸的能力,對于發展醬油產業、控制或降低醬油中EC含量、提高發酵食品的安全性具有重要意義。
1 材料和方法
1.1 菌株
嗜鹽四聯球菌(Tetragenococcushalophilus)R23,分離自高鹽稀態醬油的醬醪,保藏于中國典型培養物保藏中心(保藏編號:CCTCCNo:M2013480)。
1.2 培養基
嗜鹽四聯球菌培養基(g/L):蛋白胨10,MnSO40.05,乙酸鈉5,MgSO40.2,蔗糖10,牛肉膏5,檸檬酸三銨2,葡萄糖 10,吐溫80 1,酵母粉4,K2HPO42,NaCl 100,pH 7.2。
初篩培養基(g/L):酵母粉5,吐溫80 1 ,MgSO40.2 ,蛋白胨5 ,MnSO40.05 ,葡萄糖0.5,FeSO40.4,檸檬酸三胺2,牛肉膏5,CaCO30.1,NaCl 100 ,吡哆醛-5-磷酸0.05,K2HPO42,瓜氨酸0.5,精氨酸3~4,pH 5.5。
復篩培養基(g/L):酵母粉5,葡萄糖0.5,MgSO40.2,牛肉膏5,MnSO40.05, 蛋白胨5,FeSO40.4,檸檬酸三胺2,吐溫80 1,CaCO30.1,吡哆醛-5-磷酸0.05,K2HPO42,NaCl 180,瓜氨酸0.5,精氨酸3~5,pH 5.5。
氨基酸利用培養基(g/L):酵母膏5,NaCl 180,葡萄糖0.5,吐溫80 1,牛肉膏5,MgSO40.2,蛋白胨5,MnSO40.05,FeSO40.4,檸檬酸三胺 2,CaCO30.1,吡哆醛-5-磷酸0.05,K2HPO42,瓜氨酸 0.5,精氨酸5,pH 5.5。
1.3 主要試劑
色譜純甲醇、瓜氨酸、色譜純乙腈、分析純三氯乙酸、鳥氨酸、氨基酸標準樣品、四氫呋喃、精氨酸購自上海生工生物工程股份有限公司。用于二乙酰一肟-氨基硫脲比色法顯色反應的試劑如文獻所述[24-25]。培養細胞/細菌總RNA提取試劑盒(DP430),反轉錄試劑盒(FastQuant RT Kit (With gDNase)(KR106))購自天根生化科技(北京)有限公司。氨測定試劑盒(100M6204)購自SIGMA公司。
1.4 誘變方法
將嗜鹽四聯球菌R23培養至對數生長期,離心收集菌體,用磷酸鹽緩沖液(pH 7.0)洗滌3次后懸浮菌體,分別進行紫外誘變和等離子誘變。
(1)紫外誘變
將菌液置于15 W紫外燈下30 cm處進行紫外誘變,誘變時間分別為0、20、30、60、90、100、120 s。照射后菌液置于避光處靜置培養30 min,避光稀釋涂平板,于30 ℃恒溫避光培養3 d。根據誘變致死率繪制誘變致死率曲線,選擇致死率在90%左右的誘變條件,確定為:距15 W紫外燈30 cm處、照射2 min。
(2)等離子誘變
取10 μL的菌懸液滴加在無菌載玻片上,用常壓室溫等離子體(ARTP)進行等離子誘變,在功率100 W,通氣量為10 SLM條件下分別誘變0、1、3、5、7、10 s,然后轉移至含1 mL生理鹽水EP管中,將樣品梯度稀釋后涂布于嗜鹽四聯球菌培養基平板上,30 ℃培養3 d。根據存活率繪制誘變致死率曲線,選擇致死率在95%左右的誘變劑量,確定誘變條件為:在功率100 W、通氣量為10 SLM條件下照射3 s。
1.5 突變株篩選
(1)高通量轉接培養:挑取突變株單菌落,接種至分別含有1 mL,3、3.5、4 g/L精氨酸初篩培養基的96深孔板中,30 ℃培養5 d。
(2)初篩:利用二乙酰一肟-氨基硫脲比色法測定瓜氨酸含量[26],進行誘變菌株初篩,利用酶標儀測定溶液530 nm處吸光值(OD530),以未進行誘變育種的嗜鹽四聯球菌R23的培養基比色測定吸光值作為對照組,選擇吸光值更小的突變株進行復篩。
(3)復篩:將初篩得到的菌株接種至有1 mL復篩培養基的深96孔板中,30 ℃靜置培養5 d,離心,利用二乙酰一肟-氨基硫脲比色法分別測得在530 nm處吸光值(OD530),以嗜鹽四聯球菌R23的培養基比色測定吸光值作為對照,選擇吸光值較其更小的突變株作為復篩菌株,并利用高效液相色譜法(HPLC)測定復篩菌株培養體系中氨基酸含量,得到優勢突變株。
1.6 氨基酸測定
利用HPLC測定體系中游離氨基酸的含量[26]。測定時取 1 mL菌液,10 000 r/min離心5 min,取上清液。用質量分數為5%的三氯乙酸將其稀釋5倍,用濾膜(0.22 μm)過濾稀釋液至液相瓶中,用HPLC測定濾液中瓜氨酸、鳥氨酸和精氨酸含量。測定條件為:色譜柱 ODS-2 HYPERSIL(250 mm×4.6 mm,5 μm),流速:1 mL/min。檢測器為 VWD 紫外檢測器,柱溫 40 ℃,檢測波長為 338 nm,分離時間 38 min[10]。流動相 A (1 L):無水乙酸鈉 5 g,超純水 1 L,三乙胺 200 μL,四氫呋喃5 mL, pH 7.2。流動相 B (1 L):無水乙酸鈉 5 g,超純水200 mL,甲醇 400 mL,乙腈 400 mL, pH 7.2。
1.7 環境因素對突變株利用精氨酸和瓜氨酸能力的影響
將突變株和野生菌株R23接種至嗜鹽四聯球菌培養基,30 ℃靜置培養3 d,離心收集菌體并用磷酸鹽緩沖液(pH7.0)洗滌。考察葡萄糖濃度、溫度以及醬油乙醇發酵時期環境因素對突變株利用精氨酸和瓜氨酸能力的影響時,菌體再用相應氨基酸利用培養基垂懸,在30 ℃(考察溫度影響在設定溫度下培養)靜置培養5 d,測定精氨酸和瓜氨酸的消耗量。
1.8 酶活的測定方法
粗酶液的制備:取30 mL對數生長后期嗜鹽四聯球菌菌液離心收集菌體,用磷酸鹽緩沖液(pH 7.0)洗滌后加入等體積的氨基酸利用培養基,30 ℃培養5 h,4 500 r/min離心 5 min 收集菌體。用磷酸鹽緩沖液(pH 7.0)洗滌并垂懸菌體,破壁取上清即得粗酶液。
(1)精氨酸脫亞胺酶(ADI)酶活的測定
在5 mL的EP管中加入150 μL的50 mmol/L精氨酸、2.3 mL的50 mmol/L磷酸鹽緩沖液(pH7.0)和50 μL粗酶液,37 ℃水浴30 min后用5%的三氯乙酸終止反應,對照用50 mmol/L磷酸鹽緩沖液(pH7.0)代替粗酶液。用高效液相色譜法測定反應液中瓜氨酸含量,計算ADI酶活(ADI 酶活為稀釋倍數乘以單位時間內轉化生成的瓜氨酸含量)。
(2)鳥氨酸轉氨酶(OTC)酶活的測定
在5 mL的EP管中加入160 μL的 50 mmol/L磷酸鹽緩沖液(pH 7.0)、40 μL的25 mmol/L鳥氨酸溶液、150 μL的133 mmol/L 氨甲酰磷酸溶液、240 μL的粗酶液和600 μL的超純水,37 ℃水浴15 min后用5%的三氯乙酸終止反應,對照用50 mmol/L磷酸鹽緩沖液(pH7.0)代替粗酶液。用高效液相色譜法測定反應液中瓜氨酸含量,計算 OTC 酶活(OTC 酶活為稀釋倍數乘以單位時間內轉化生成的瓜氨酸含量)。
(3)氨基甲酸酯激酶(CK)酶活的測定
在1.5 mL的EP管中加入160 μL 的50 mmol/L磷酸鹽緩沖液(pH 7.0)、200 μL的73 mmol/L的MgCl2溶液以及200 μL的50 mmol/L ADP,混勻后放置 10 min,加入200 μL的133 mmol/L氨甲酰磷酸溶液,混勻后繼續放置 10 min,向混合液中加入100 μL粗酶液,37 ℃水浴15 min后用5%的三氯乙酸終止反應,對照用50 mmol/L磷酸鹽緩沖液(pH7.0)代替粗酶液。將反應液稀釋一定倍數后,按照氨測定試劑盒(SIGMA公司)中的測定方法測定CK酶活(CK酶活為稀釋倍數乘以單位時間內轉化生成的NH3含量)。
1.9 實時定量PCR法分析基因轉錄水平
將嗜鹽四聯球菌培養至對數生長期,離心收集菌體,用磷酸鹽緩沖液(pH 7.0)洗滌3遍后分別垂懸于含4 g/L和5 g/L精氨酸的氨基酸利用培養基中,30 ℃培養4 h。低速離心獲得菌體,與2 mL RNA 穩定劑混合,-80 ℃保藏。采用液氮研磨的方式對上述獲得的菌體進行破壁。用培養細胞/細菌總RNA提取試劑盒(天根生化科技(北京)有限公司)提取RNA并用反轉錄試劑盒(FastQuant RT Kit (With gDNase) (天根生化科技(北京)有限公司)制備cDNA。qRT-PCR使用的引物如表1所示,選擇16S rRNA作為內參基因。使用2-ΔΔCt的方法進行轉錄水平分析[27]。
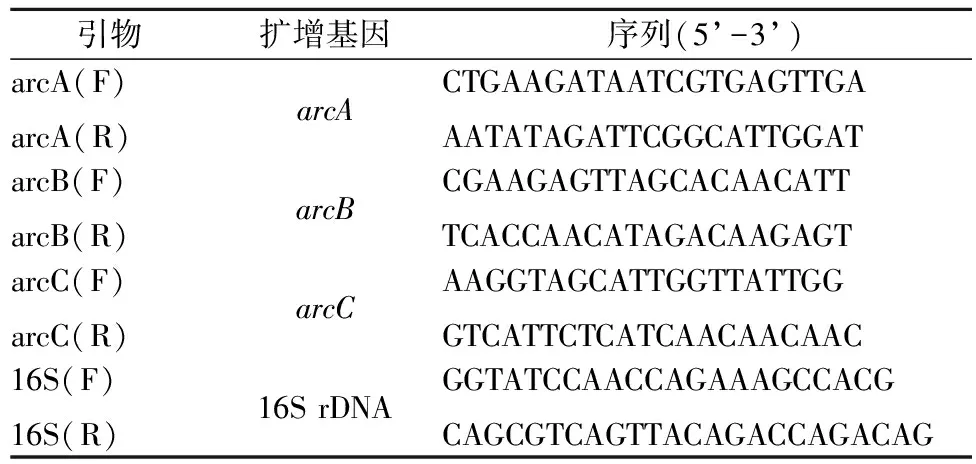
表1 用于實時定量PCR的引物Table 1 Primers for qRT-PCR
2 結果和分析
2.1 嗜鹽四聯球菌誘變育種及高效利用瓜氨酸突變株篩選
嗜鹽四聯球菌R23在精氨酸質量濃度低于3 g/L時可利用醬醪中的精氨酸且不積累EC前體瓜氨酸,但是當精氨酸含量高于3 g/L時,體系中瓜氨酸開始積累[22]。為獲得精氨酸和瓜氨酸利用能力提高的菌株,通過誘變嗜鹽四聯球菌R23,共得到紫外誘變突變株386株,等離子誘變突變株237株。采用二乙酰-肟-氨基硫脲比色法確定瓜氨酸相對含量的篩選方法,用于突變株的初篩(圖1)[25]。
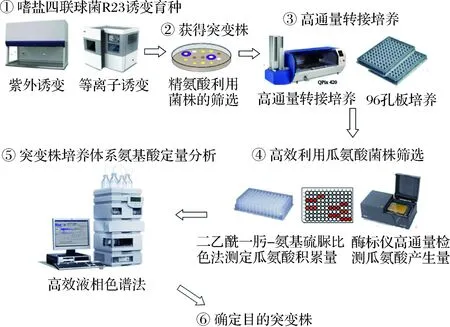
圖1 嗜鹽四聯球菌誘變及突變株篩選Fig.1 Mutagenesis of Tetragenococcus halophilus and screening of mutants
通過初篩和復篩,獲得了4株在精氨酸質量濃度高于3 g/L時,瓜氨酸積累少于出發菌株R23的突變株,分別命名為1-A2,1-A10,5-H1和3-H9(圖2)。
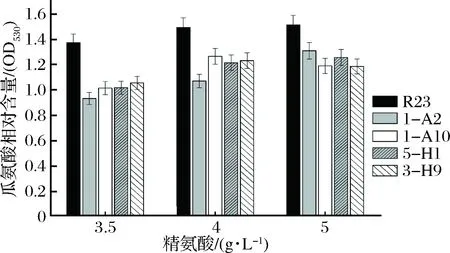
圖2 嗜鹽四聯球菌利用精氨酸和積累瓜氨酸能力分析Fig.2 Conversion of citrulline from arginine by T. halophilus 1-A2,1-A10,5-H1 and 3-H9,mutants of T. halophilus R23
2.2 嗜鹽四聯球菌突變株利用精氨酸和瓜氨酸的能力
為進一步確定突變株利用精氨酸和瓜氨酸的能力,以及尋找具有有效降低醬油中EC含量潛力的菌株,分別對4株嗜鹽四聯球菌突變株利用精氨酸和瓜氨酸的能力進行了分析和比較(圖3)。
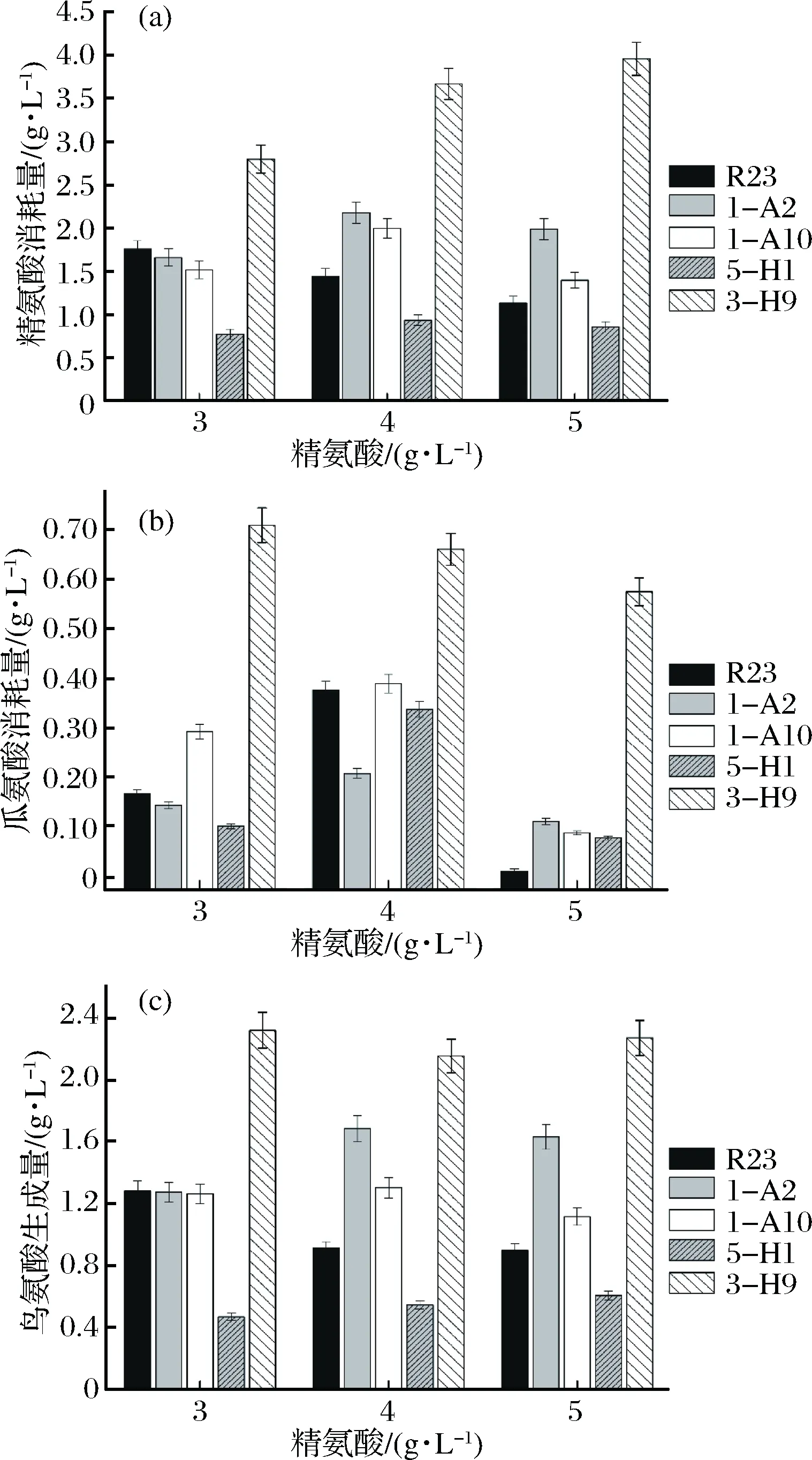
圖3 嗜鹽四聯球菌利用精氨酸和瓜氨酸的能力比較Fig.3 Utilization of arginine and citrulline by T. halophilus 1-A2,1-A10,5-H1 and 3-H9,mutants of T. halophilus R23
由圖3可以看出,當體系中含有0.5 g/L瓜氨酸和3、4、5 g/L精氨酸時,突變株3-H9利用精氨酸能力最高,比R23提高了0.6~2.49倍。突變株3-H9利用瓜氨酸能力也比R23提高了0.74~56倍。突變株5-H1精氨酸利用能力下降,瓜氨酸利用能力沒有顯著提高。突變株1-H10和1-A2利用精氨酸的能力比R23略有提高,但對瓜氨酸的利用能力遠低于突變株3-H9。因此,選擇突變株3-H9進行后續研究。
2.3 環境因素對突變株利用精氨酸和瓜氨酸能力的影響
ADI途徑是細菌獲得能量的一條輔助代謝途徑。當培養體系中還原糖充足時菌株優先利用還原糖供能,因此ADI途徑代謝通量會受到還原糖的抑制[10]。由于乙醇和游離脂肪酸的協同作用,細菌通過ADI途徑代謝精氨酸時,會積累瓜氨酸,對EC前體含量的有效控制時期為醬油發酵前期,即乳酸發酵時期[28]。在醬油發酵前期(1~7 d),醬醪中葡萄糖的質量濃度為0~7 g/L[29]。通過考察葡萄糖濃度對突變株利用精氨酸、瓜氨酸能力的影響發現,葡萄糖質量濃度大于2 g/L時野生菌株R23已不能消耗精氨酸,而突變株3-H9在葡萄糖質量濃度為7 g/L時仍能利用精氨酸。突變株3-H9利用精氨酸的能力在葡萄糖質量濃度為2、3、5、7 g/L時,分別比R23提高 5.05、5.62、3.41、1.22 g/L(表2)。這說明,突變株3-H9精氨酸代謝受葡萄糖抑制作用被減弱,具有進一步減控醬油中EC前體的潛力。
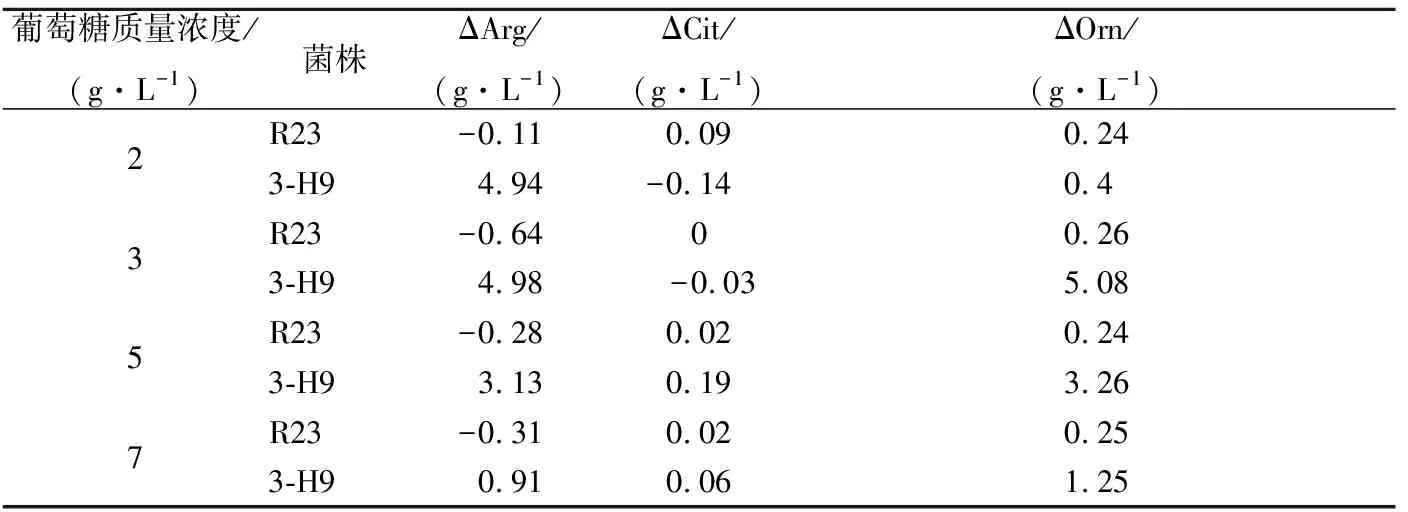
表2 葡萄糖對嗜鹽四聯球菌利用精氨酸和瓜氨酸能力的影響Table 2 Effects of glucose concentration on arginine and citrulline utilization of T. halophilus mutants
注: ΔArg為精氨酸消耗量;ΔCit為瓜氨酸消耗量; ΔOrn為鳥氨酸生成量。
高鹽稀態醬油發酵過程前期溫度較低,為了研究溫度對嗜鹽四聯球菌突變株利用精氨酸能力的影響,考察了突變株3-H9在20、25、30 ℃條件下利用精氨酸能力。由表3可知,溫度低于30 ℃時,突變株3-H9和野生菌株R23利用瓜氨酸和生成鳥氨酸的能力均有所減弱。較低溫培養條件下,突變株3-H9利用精氨酸的能力高于野生菌株R23。在20、25 ℃培養時,3-H9利用精氨酸能力分別比R23提高57.7%和47.8%(圖4)。

表3 溫度對突變株利用瓜氨酸能力的影響Table 3 Effects of temperature on the use of citrulline in mutant strains
注:ΔCit為瓜氨酸消耗量(g/L); ΔOrn為鳥氨酸生成量(g/L)。
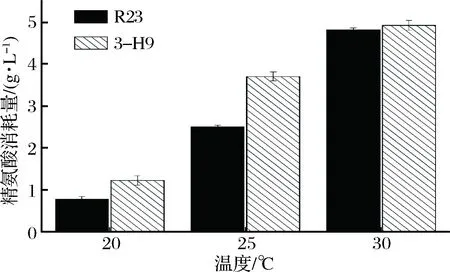
圖4 溫度對嗜鹽四聯球菌利用精氨酸的影響Fig.4 Effect of temperature on arginine utilization of T. halophilus
醬油發酵過程中,瓜氨酸的積累有2個時期:一是乳酸發酵時期,占主導地位的微生物是乳酸菌(主要為魏斯氏菌屬和足球菌屬的菌株),二是乙醇發酵時期,葡萄球菌屬、微球菌屬和芽孢桿菌屬是優勢菌群[28,30-31]。乙醇發酵時期醬醪中含有的乙醇和游離脂肪酸是導致細菌利用精氨酸積累瓜氨酸的主要因素。因此,我們考察了突變株3-H9在含有2%的乙醇與0.25%游離脂肪酸(模擬醬油乙醇發酵時期)的培養體系中利用精氨酸的情況。由表4可知,在此條件下,野生菌株R23不利用精氨酸,而突變株3-H9仍可利用精氨酸。由于突變株3-H9在這一培養條件下利用精氨酸積累了瓜氨酸,因此它更適用于在醬油乳酸發酵時期發揮作用。

表4 乙醇和游離脂肪酸對嗜鹽四聯球菌利用精氨酸和瓜氨酸能力的影響Table 4 Effect of ethanol and fatty acids on arginine and citrulline utilization of T. halophilus
注: ΔArg為精氨酸消耗量;ΔCit為瓜氨酸消耗量; ΔOrn為鳥氨酸生成量。
2.4 突變株瓜氨酸代謝相關基因轉錄水平及酶活分析
細菌對精氨酸的利用以及瓜氨酸的生成和消耗與ADI途徑中3種關鍵酶:精氨酸脫亞胺酶(ADI)、鳥氨酸轉氨酶(OTC)和鳥氨酸氨基甲酸激酶(CK)密切相關。由表5結果可知,突變株3-H9較R23相比,ADI酶、OTC酶和CK酶活性分別提高了27%、38%和0.4%,說明突變株3-H9利用精氨酸和瓜氨酸的能力都比出發菌株高。

表5 突變株及野生菌株中 ADI、OTC、CK酶酶活分析Table 5 Activity of ADI,OTC,CK in mutant and wild strains
ADI酶、OTC酶和CK酶是分別由arcA、arcB和arcC基因編碼。為了探明突變株3-H9利用精氨酸和瓜氨酸能力及調控水平變化的原因,對突變株3-H9和野生菌株R23arc基因簇轉錄水平進行了比較分析。由圖5可知,突變株3-H9中arcA、arcB和arcC轉錄水平分別比R23提高了7.9倍、9.5倍和7.8倍。在ADI途徑中,arcA編碼的ADI酶催化精氨酸向瓜氨酸轉化,arcB基因編碼的OTC酶催化瓜氨酸向鳥氨酸轉化。通過比較arcA與arcB基因表達水平可以得知該條件下ADI途徑產物是終產物鳥氨酸,還是含有中間產物瓜氨酸。此途徑中,若arcA/arcB大于1,表現為瓜氨酸積累,arcA/arcB小于1則表現為瓜氨酸利用。通過對突變株3-H9和野生菌株R23的arcA/arcB轉錄水平之比分析發現:在4 g/L精氨酸濃度下,兩株菌arcA/arcB比值均小于1,均表現為利用瓜氨酸;當精氨酸濃度為5 g/L時,野生菌arcA/arcB>1,表現為積累瓜氨酸,而突變株3-H9arcA/arcB<1,表現為利用瓜氨酸(表6)。
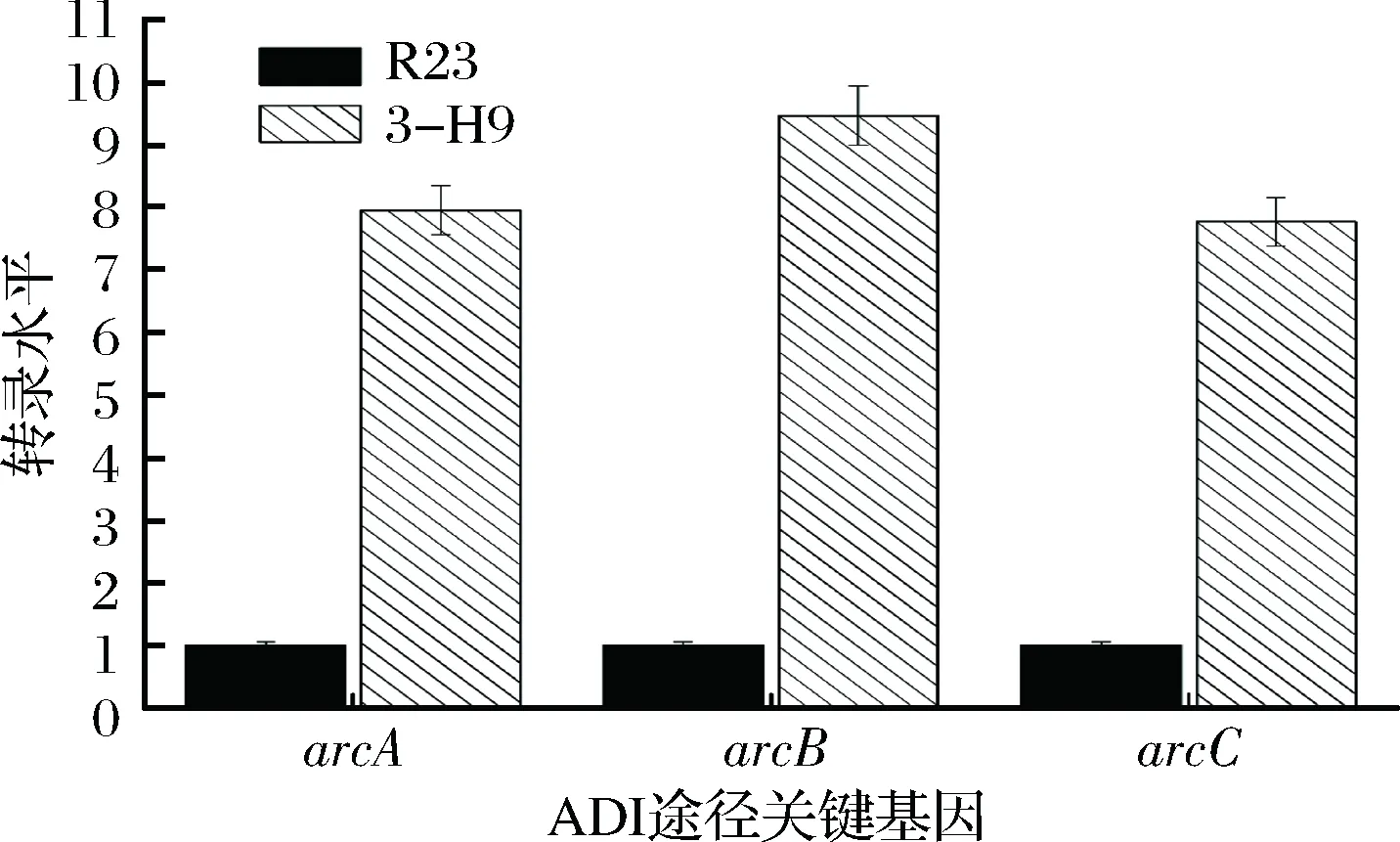
圖5 ADI途徑相關基因轉錄水平分析Fig.5 Transcriptional analysis of arc genes in T. halophilus
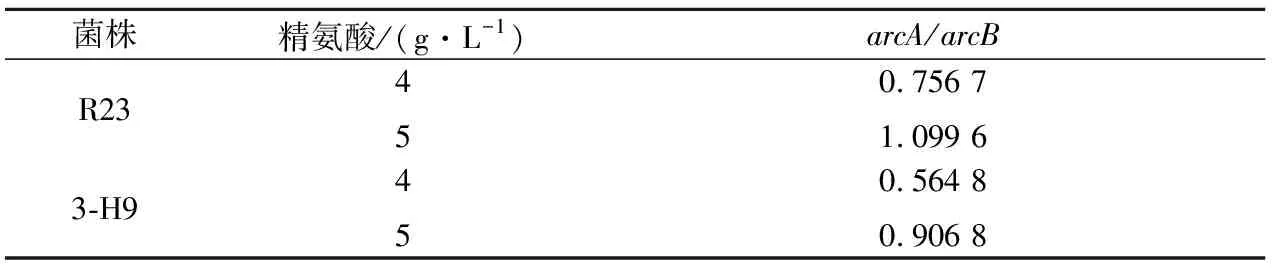
菌株精氨酸/(g·L-1)arcA/arcBR23450.756 71.099 63-H9450.564 80.906 8
3 結論
本研究以對醬油風味有強化作用的內源微生物嗜鹽四聯球菌為出發菌株,通過等離子和紫外誘變的方法獲得了1株利用精氨酸和瓜氨酸能力顯著提高的嗜鹽四聯球菌突變株3-H9。該突變株具有減少醬油中氨基甲酸乙酯前體瓜氨酸的應用潛力。研究結果提供了一種通過改造發酵食品體系中某一有益菌的氮源代謝方式來減少發酵食品中有害物的策略,可為控制或降低醬油中氨基甲酸乙酯及其前體提供理論依據,對于提高嗜鹽四聯球菌的工業應用性能以及發展醬油產業,提高食品的安全性具有有利意義。
猜你喜歡
發明與創新(2022年30期)2022-10-03 08:40:56
中等數學(2022年2期)2022-06-05 07:10:50
中學生數理化·七年級數學人教版(2021年11期)2021-12-06 05:38:48
動漫星空(興趣百科)(2020年12期)2020-12-12 05:31:40
小學生學習指導(低年級)(2020年6期)2020-07-25 02:31:36
小學生學習指導(低年級)(2018年9期)2018-09-26 05:59:44
瘋狂英語·新讀寫(2018年2期)2018-09-07 09:32:10
人大建設(2018年6期)2018-08-16 07:23:10
文理導航·科普童話(2017年5期)2018-02-10 19:42:14
無人機(2017年10期)2017-07-06 03:04:36
