QUARTET突變四倍體擬南芥的獲得與表型分析
2018-08-18 01:44:20何碩康羅澤偉
生物技術通報 2018年7期
關鍵詞:背景
何碩康 羅澤偉
(復旦大學生命科學學院 生物統計所,上海 200433)
多倍化(Polyploidy)即生物體合子中擁有超過兩套染色體的現象,廣泛存在于自然界中,如魚類[1]、甲殼類[2]以及兩棲類[3]。植物中多倍化現象更是普遍存在。80%以上的被子植物在進化過程中存在多倍性[4]。以多倍體形式存在的物種有更豐富的基因型與表型多態性,進而提高對環境的適應性與群體遺傳多態性[5]。此外,多倍化也是產生與保持雜種優勢的重要途徑之一[6]。根據多倍體中染色體組的遺傳來源,可將多倍體分為同源多倍體(Autopolyploid)與異源多倍體(Allopolyploid)[7]。
多倍化會對細胞減數分裂產生深刻的影響[8]。在多倍體減數分裂過程中,既有可能染色體之間兩兩交叉形成二價體,染色體行為類似于二倍體,這種模式被稱為二體遺傳;也有可能超過兩條同源染色體或近源染色體之間交叉形成多價體,這種模式被稱為多體遺傳,這導致多倍體遺傳要比二倍體遺傳復雜很多[9]。目前對于多倍體遺傳的研究都是建立在群體數據的基礎上。即利用兩套背景不同、具有多態性標簽位點的染色體構建雜合同源或異源多倍體親本,然后對其后代回交或自交群體采集一定大小的樣本,進行多態性位點的檢測并繪制圖譜,統計分析[10]。實際上,群體數據的分析是極其依賴模型的,依據的模型不同所得結果常有較大差異[11,12]。但目前多是利用理論分析與計算機模擬的方法建模,而沒有一個由準確實驗數據支持的模型。因此我們希望利用四分子分析,這一能夠精準分析單次減數分裂的方法[13],將多倍體遺傳重組的研究從群體水平精確到個體水平,構建有實驗數據支持的多倍體遺傳重組模型。20世紀90年代以來,在擬南芥(Arabidopsis thaliana)中陸續發現的3個基因[14,15],這3個基因任意一個的純合失活突變都能使花粉母細胞單次減數分裂發育而來的4個花粉互不分離,并由此將這3個基因命名為QUARTET(QRT1、QRT2以及QRT3),是應用四分子分析的極好材料。因此本研究從帶有qrt1突變的二倍體擬南芥Columbia及Landsberg erecta生態型出發,誘導并篩選染色體組成功加倍的四倍體qrt1突變株,并測定它們與二倍體及四倍體參照的表型差異,能為在多倍體遺傳重組中應用四分子分析奠定基礎。
1 材料與方法
1.1 材料
野生型Columbia生態型二倍體Col-0及其四倍體CS3151、野生型Landsberg erecta生態型二倍體Ler及其四倍體CS3900均為實驗室保存。二倍體Columbia QRT1 -/- 突變體、二倍體Landsberg erecta QRT1 -/- 突變體以及特異性識別 擬南芥(Arabidopsis thaliana)著絲粒位置180 bp重復序列的熒光原位雜交探針獲贈于陸平利研究員(復旦大學生命科學院植物科學研究所)。
1.2 方法
1.2.1 擬南芥的誘導與傳代 用0.08%濃度的秋水仙素溶液處理10 d齡的QRT1 -/- 二倍體Columbia與Landsberg erecta幼苗(G0)5 h。培養G0幼苗,收集G0的種子。將這批種子種下,從這批苗(G1)中鑒定與篩選成功加倍的植株,收集這些植株的種子,傳代,并于后代中再次驗證倍性。
1.2.2 流式細胞技術 取50 mg左右植株幼嫩蓮座葉片,參照文獻[16]處理。不同的是,這里葉片材料經過破碎之后經過30 μm孔徑尼龍膜過濾,并用100 μg/mL的碘化丙啶(PI)染色30 min左右。所用儀器為FACSCalibur流式細胞儀(BD Biosciences),分析軟件為CellQuestPro。所用熒光染料為PI,選擇熒光檢測通道為FL2,同時對FL-2的值取對數,使等比DNA含量的峰在圖上顯示為等間距的峰。
1.2.3 熒光原位雜交技術 取卡諾氏液固定好的幼嫩花序,按照前人方式[17]處理,觀察其中花粉母細胞的減數分裂。DNA染料為4,6-二脒基-2-苯基吲哚(DAPI),同時利用特異性識別擬南芥著絲粒位置180 bp重復序列的熒光原位雜交探針標識著絲粒的位置,進行染色體計數。熒光探針生色基團為Cy3,識別序列以及制作過程參照文獻進行[18]。
2 結果
2.1 四倍體的誘導與流式細胞儀檢測倍性結果
本研究中,為確定細胞倍性,利用流式細胞技術,以二倍體野生型Col與Ler(圖1-A,1-B)、四倍體野生型Col與Ler(圖1-G,1-H)為對照,從Columbia背景篩選出并編號為C313的植株(圖1-C),從Ler背景篩選出并編號為L311的植株(圖1-D)。自交傳代,并在G3中再次用流式細胞技術驗證C313與L311的倍性(圖1-E,1-F)。
本研究中的流式結果,顯示了擬南芥中廣泛存在的內復制現象[19],即核基因組復制而細胞不分裂的現象。這導致擬南芥葉片實際為多種倍性細胞共存的嵌合體,故其流式結果中有多個峰。而由于熒光強度取了對數,故等比的峰表現為等間距的峰。而二倍體與四倍體的區別就在于,二倍體有2n的峰(圖1-A,1-B),而四倍體沒有(圖1-G,1-H)。經過G1與G3的重復驗證,流式結果驗證C313與L311核基因組加倍成功。另外,有文章[20]報道擬南芥的內復制速率隨著倍性的提高而提高,在本研究中,也基本反映了這一現象(圖1-C,1-E,1-F,1-G,1-H),在這幾張圖中,4n的峰相比于二倍體中2n的峰(圖1-A,1-B)要更低,而內復制之后的峰相對的更高,表現出更高的內復制速率。
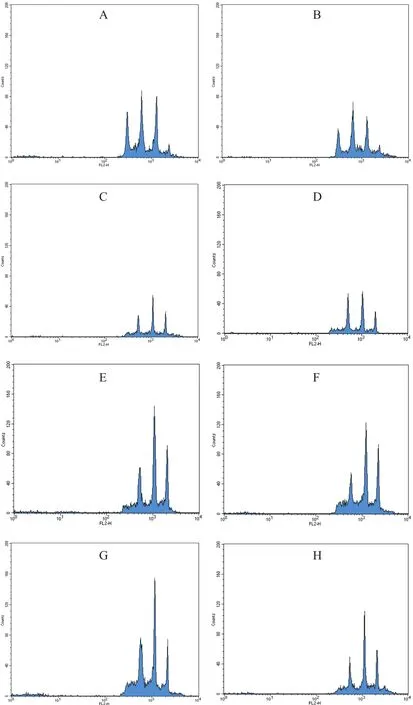
圖1 流式細胞技術檢測細胞倍性
2.2 熒光原位雜交技術鑒定倍性結果
在利用流式初步在G1群體中篩選出C313與L311植株后,于花期觀察花粉母細胞減數分裂(圖2-d-i)。在細線期,細胞已完成DNA復制,染色體開始凝聚。此時姐妹染色單體通過著絲粒相連,尚未分開。而同源染色體尚未配對。因此染色質彌散于核內,每一個著絲粒都代表一條染色體,代表其上的一對姐妹染色單體。可見C313與L311中皆能數到20個點,而二倍體對照(圖2-a-c)只能數到10個點,驗證了倍性。并繼續傳代,至G3時,再次觀察花粉母細胞減數分裂(圖2-j-p),驗證了C313與L311染色體加倍成功。
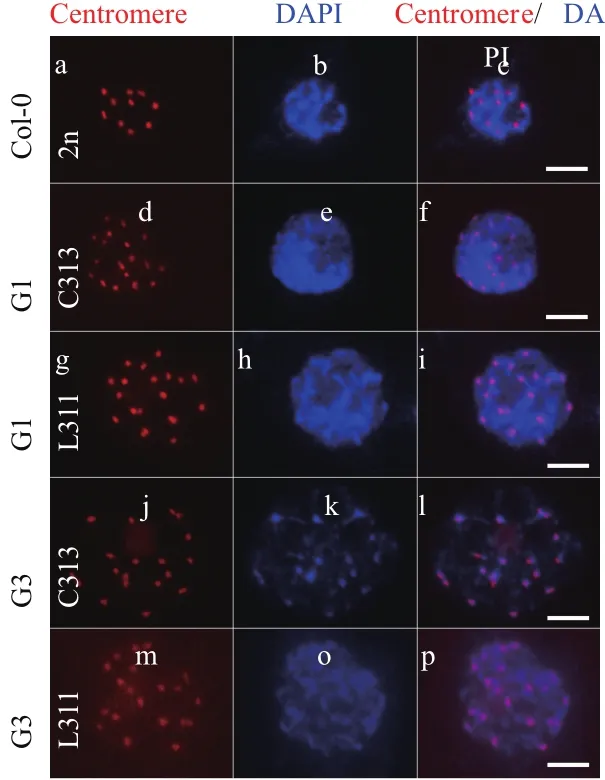
圖2 熒光原位雜交技術檢測細胞倍性
2.3 營養生長相關表型的測定與分析
2.3.1 倍性對主根及側根生長的影響 利用垂直平板培養記錄種子萌發期根的生長,包括縱向深度(主根長度)和橫向廣度(測根數目)。結果發現在Col與Ler背景中,多倍化都不對主根的生長造成影響,而qrt1突變在Ler中對主根生長有抑制作用(圖3-A-C),p=2.2e-16)。至于側根,多倍化以及qrt1突變在兩個被背景中都會抑制側根數目,而在Ler中,多倍化及qrt1突變的抑制作用顯現出一定的可加性,而在Col中則沒有(圖3-D,E)。
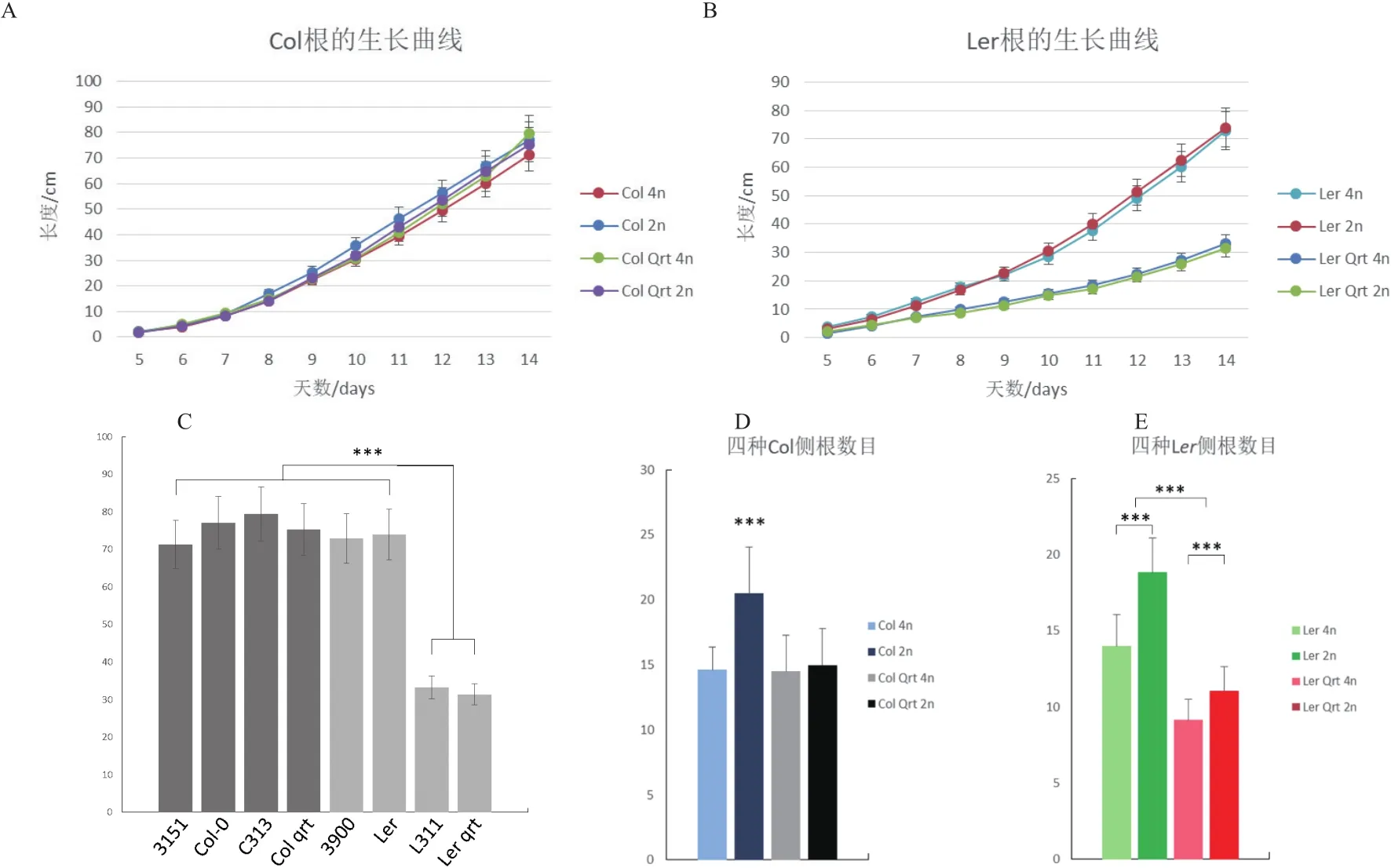
圖3 根的相關表型
2.3.2 倍性對莖的影響 與一般多倍化導致植株體型增大的報道不同,本研究中發現倍性對主莖高度的影響具有背景依賴性(圖4-C)),即在Col中,四倍體更高(37.72 cm增長至41.41 cm,p=2.06e-17),而在Ler中,四倍體更矮(33.52 cm降低至31.56 cm,p=2.50e-10),也許是由于樣本量(30株)不夠大導致的。同時,四倍體中主莖抽薹日期也被推遲(圖4-D),Col中由26.2 d推遲至39.0 d,Ler中由24.6 d推遲至35.0 d)。至于分枝,我們又分了兩個表型,一個是近蓮座的基部側枝(類似于分蘗),一個是距離土壤有一定高度的主莖分枝。多倍化僅僅在Ler背景中抑制了主莖分枝的數量,在Col背景中則無影響(圖4-B)。而對于基部側枝這個表型,不管是多倍化還是qrt1突變,都具有顯著的抑制效果,同時二者抑制作用可疊加(圖4-A)。實際上,在四倍體主莖基部,可以觀察到未生長的芽,所以四倍化抑制基部側枝數目主要是通過抑制芽的生長而不是芽的發生。
2.3.3 倍性促進葉的初生生長 至于對葉的影響,在兩個背景中,多倍化都導致蓮座葉片面積更大,同時qrt1突變都抑制了葉片面積(圖4-E)。
2.4 生殖生長相關表型的測定與分析
首先對花藥進行亞歷山大紅染色(圖5-A),所有的花粉都被染上了紅色,說明多倍化或者qrt1突變都不會影響花粉的活性。其次,我們測量了花粉(圖5-B)以及花藥中隔(圖5-C)的大小,發現多倍化都使其體積增大。至于果莢,多倍化導致每果莢種子數顯著降低(二倍體Col平均每果莢50.49粒,四倍體Col平均每果莢30.18粒;二倍體Ler平均每果莢40.99粒,四倍體Ler平均每果莢18.84粒)。
本研究中新制四倍體C313與L311相比四倍體參照,每果莢種子數又進一步顯著性的降低(圖5-D),3151平均每果莢32.37粒,C313平均每果莢27.98粒,3900平均每果莢20.28粒,L311平均每果莢17.40粒)。多倍化對果莢長度的影響具有背景依賴性(圖5-E),在Col中沒有影響,而在Ler中使果莢長度顯著降低(p=2.29e-68)。
多倍化對每株果莢個數的影響同樣具有背景依賴性(圖5-F),但并不是依賴其是Col還是Ler,而是依賴于qrt1突變與否。在二倍體中,qrt1突變使果莢數顯著降低。在此基礎上,多倍化使野生型果莢數降低,使qrt1突變體果莢數上升。
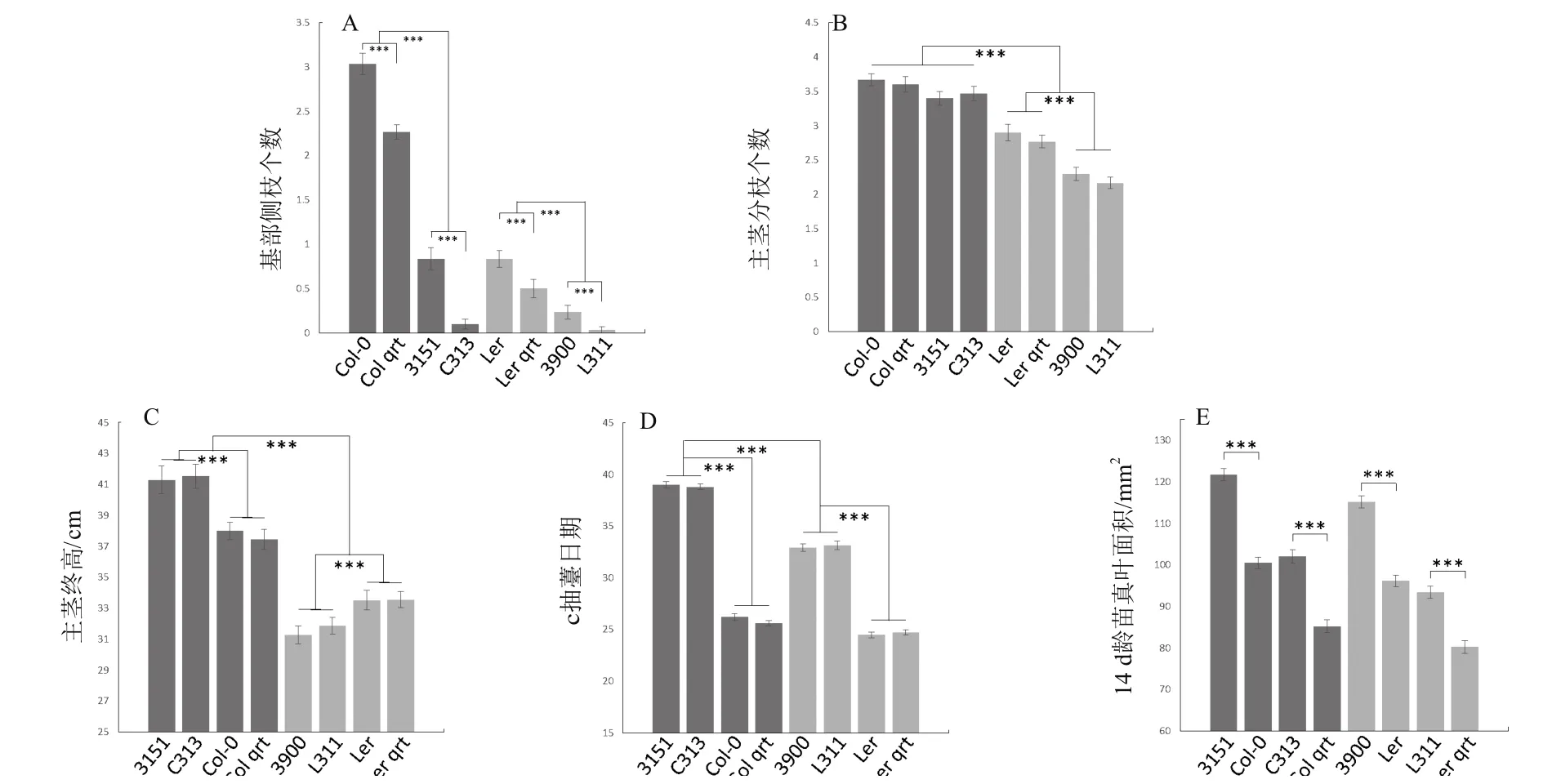
圖4 莖及葉的相關表型
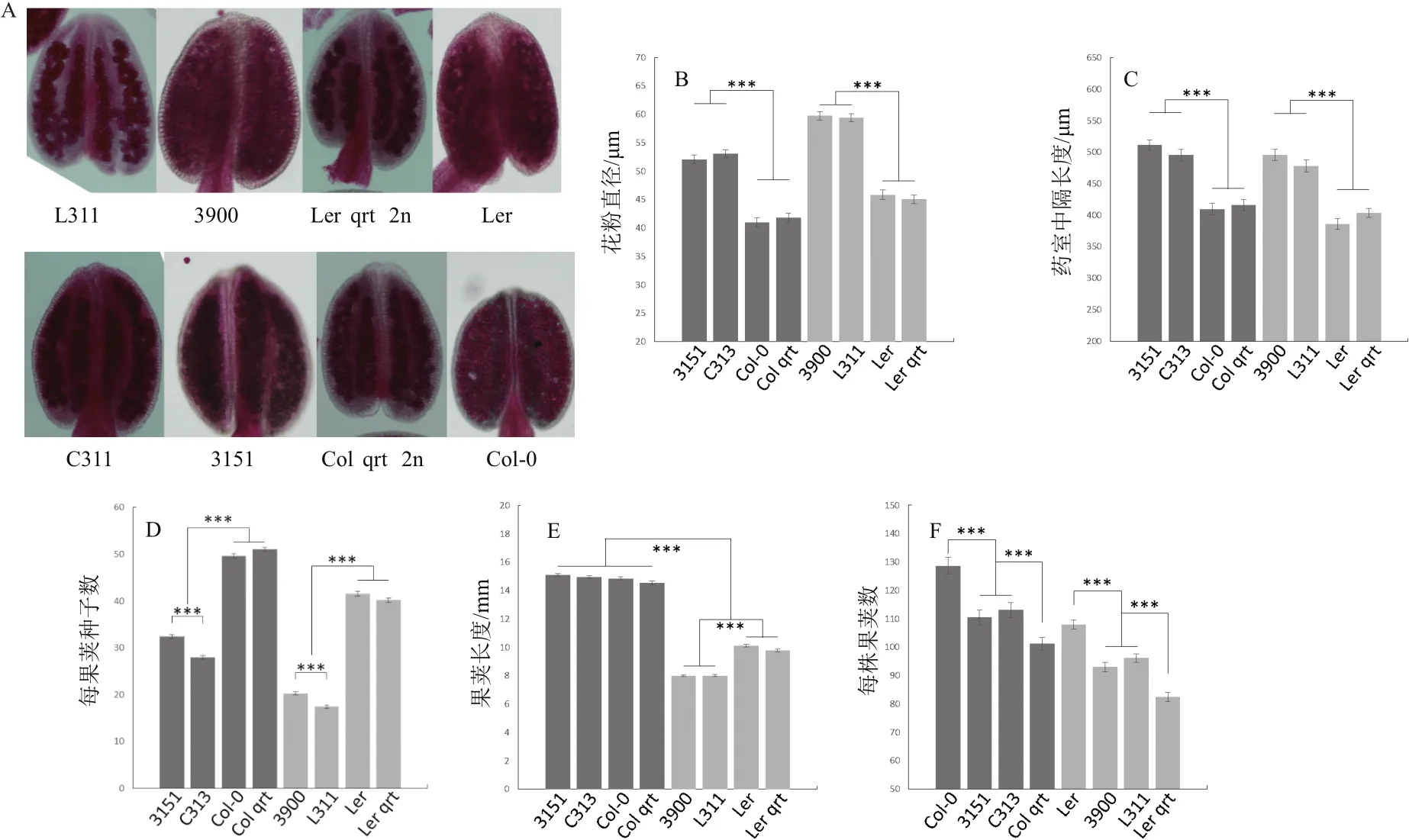
圖5 花以及果莢相關表型
在對種子影響上,四倍化使種子顯著增大(圖6-A)。大小,即種子的長軸長與短軸長,形狀,即種子的離心率。四倍化導致種子的長軸與短軸同時增大,并最終保持種子的離心率,即形狀基本不變(圖6-B-D)。當然Col與Ler,不同背景之間,種子的形狀就有差異,Col種子離心率更大,種子更扁。同時,加倍之前,種子長軸基本一致,而加倍之后,短軸基本一致(圖6-C-D)。在種子的發芽率方面,8個背景種子的發芽率都接近100%(圖6-F),可見雖然多倍化導致每果莢種子數大幅降低,但結出的種子都仍然保持著很好的發芽率。

圖6 種子相關表型
3 討論
四分子分析是研究遺傳重組的一項強有力的手段,但一直以來,由于高等真核生物中沒有適合應用四分子分析的物理結構,導致這種實驗手段無法被應用于真核生物減數分裂的研究。但隨著QUARTET基因被發現,在擬南芥中應用四分子分析成為了可能。陸平利教授[21]應用四分子分析成功分析了二倍體Col qrt與Ler qrt雜交后代的遺傳重組。
在本研究中,首先從帶有QUARTET1突變的二倍體Columbia與Landsberg erecta生態型擬南芥出發,利用秋水仙素誘導體細胞突變,并從嵌合體植株的后代中,利用流式細胞儀與熒光原位雜交技術,通過對兩個傳代群體之間的縱向重復以及群體內部的橫向重復,篩選出成功加倍的帶有qrt1突變的四倍體Columbia與Landsberg erecta生態型擬南芥C313與L311。為后續構建Col與Ler的雜交分離四倍體群體,并應用四分子分析研究多倍體遺傳重組準備了材料。其次,對篩選出的Col與Ler同源四倍體的根、莖、葉、花、果莢、種子等表型進行了測定并與相對應的二倍體、四倍體Col、Ler參照進行了比對。在前人對QUARTET1表達定位研究的基礎上[22],通過垂直培養幼苗,統計側根數目,計算葉片面積,進一步確認了qrt1突變抑制了側根生長以及葉的初生生長。本研究在Columbia與Landsberg erecta兩個生態型中。既觀察到了被廣泛闡述的四倍化對營養生長、生殖生長的促進作用,如蓮座葉片、花粉、花藥、種子在四倍化的影響下明顯增大;也發現了四倍化對某些器官的抑制效應,如側根數量、基部側枝數量、抽薹日期受到了四倍化的抑制;另外,還發現有些表型并未受到四倍化的影響,如主根長度、種子形狀。上述這些表型在兩個生態型背景中受到四倍化影響是相似的,同為促進或同為抑制或都不受影響,但還有一些表型,如主莖分枝數量、最終株高、果莢長度,在兩個背景中受到四倍化的影響是不同的,顯示出四倍化對植株的作用會受到整體基因組背景的影響。在育性方面,發現多倍化導致每果莢種子數變少。同時還發現了本研究中所誘導得來的新制四倍體C313與L311相對于四倍體對照,每果莢中種子更少,體現了新制四倍體的特征。盡管種子數降低了,但不管是二倍體還是四倍體對照還是本研究中的新制四倍體,都表現出乎了接近100%的發芽率。
本研究構建的帶有qrt1突變的四倍體Col與Ler擬南芥為在四倍體中應用四分子分析準備了材料;結合本研究中測定的表型,提供了研究多倍體轉錄組的實驗對象;還發現了新制四倍體與相對穩定四倍體之間的表型差異,提供了研究新制四倍體中染色體組如何變化并趨于穩定的實驗對象。
4 結論
利用秋水仙素處理帶有qrt1突變的Col與Ler生態型擬南芥,通過流式細胞技術與熒光原位雜交計數篩選出成功加倍的、帶有qrt1突變的四倍體Col與Ler生態型擬南芥,并觀察測量了8個遺傳背景擬南芥的多個營養與生殖表型,發現了qrt1突變以及倍性對擬南芥不同表型的差異化影響。
猜你喜歡
教學考試(高考化學)(2022年5期)2022-11-19 14:15:16
汽車工程師(2021年12期)2022-01-17 02:29:54
當代陜西(2020年14期)2021-01-08 09:30:42
奧秘(創新大賽)(2020年7期)2020-07-27 08:26:32
活力(2019年21期)2019-04-01 12:18:06
中國自行車(2018年10期)2018-11-30 02:09:04
貴州師范學院學報(2016年4期)2016-12-01 03:54:07
紡織服裝流行趨勢展望(2016年1期)2016-05-04 03:45:20
語文知識(2015年11期)2015-02-28 22:01:59
中國衛生(2014年10期)2014-11-12 13:10:16
